向日葵对列当寄生的生理响应及关键代谢通路分析
2024-03-08赵雅杰朱孔艳温玉洁包海柱
赵 榕,王 猛,赵雅杰,朱孔艳,温玉洁,包海柱
(内蒙古农业大学 农学院,呼和浩特 010019)
向日葵(Helianthus annuusL.)是中国主要的油料作物之一,广泛种植于内蒙古、新疆、东北三省以及甘肃、河北、宁夏、陕西等地,并以内蒙古的种植面积最大,占全国种植面积50%以上[1]。近年来向日葵列当在中国已经大面积发生,部分地区已经造成向日葵绝收,成为中国向日葵稳产、高产最大的隐患之一[2],因此,向日葵列当的防治已经成为内蒙古地区向日葵生产中亟待解决的问题。
向日葵列当(Orobanche cumana)又称毒根草,是列当科列当属的一年生全寄生草本植物,株高15~54 cm 不等[3],主要寄生在向日葵的根上,严重影响向日葵的产量和品质。研究表明,向日葵列当寄生后显著降低向日葵灌浆期植株叶面积指数,降低向日葵群体光合速率,进而影响干物质的合成与积累[4]。研究发现,在向日葵第7号染色体上存在1个列当抗性基因HaOr7,该基因编码富含亮氨酸的受体样激酶,可防御向日葵列当的寄生[5];黄瓜被列当寄生后可发生应激反应,通过提高过氧化氢酶、过氧化物酶、酚类化合物活性来一定程度上增强对列当的抗性[6];生长素(IAA)和水杨酸(SA)处理能显著降低番茄幼苗被列当感染引起的氧化损伤;SA处理的番茄种子可以通过增加抗氧化防御标志物(包括抗氧化代谢物类胡萝卜素、酚类物质、类黄酮、ASC、GSH、生育酚以及和抗氧化酶CAT、POX、GPX、SOD、GR、APX、MDHAR、DHAR)来提高列当抗性[7]。Sisou等[8]认为,当列当种子萌发后,向日葵抗性材料的初级防御系统会感知到列当幼苗对根部的附着,进而激活PR基因,产生β-葡聚糖酶,分解列当细胞壁,同时木质素和其他酚类化合物在寄生区域积累形成物理屏障,从而阻止列当与向日葵建立连接,导致列当死亡。Yang等[9]通过对向日葵感(TK0409)、抗(JY209)列当材料接种列当,并进行转录组测序,发现刺激列当萌发的向日根系分泌物在2个向日葵品种之间无显著差异,并且发现PAR1(蛋白酶激活受体)可能是潜在的植物防御反应的物质。近年来随着高通量测序技术的迅速发展,转录组学技术因具有高通量、高灵敏度、高分辨率等诸多优势而被广泛应用于植物抗逆研究中[10]。本研究以转录组学和植物生理学为基础,通过分析向日葵抗、感材料在列当寄生下的代谢通路和生理响应,明确向日葵对列当的寄生抗性反应,以期揭示向日葵对列当寄生的抗性分子机制。
1 材料和方法
1.1 试验材料
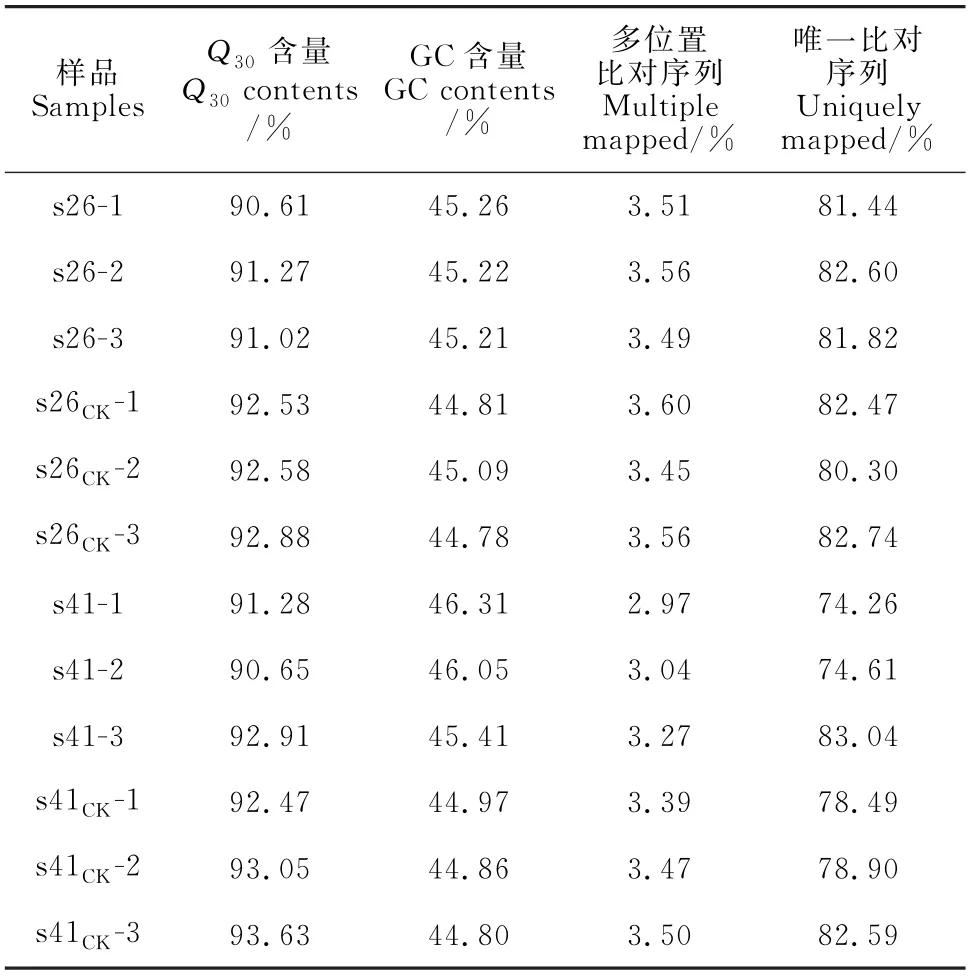
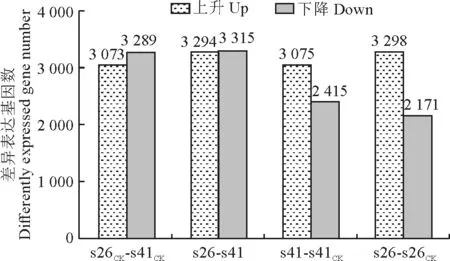
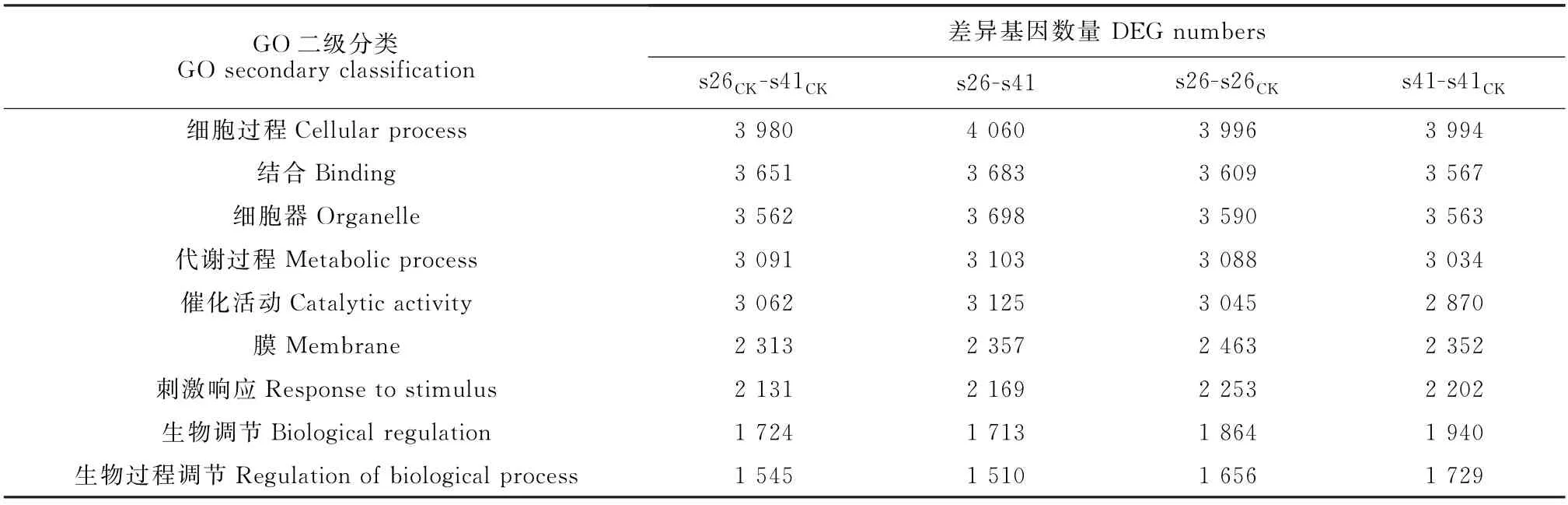
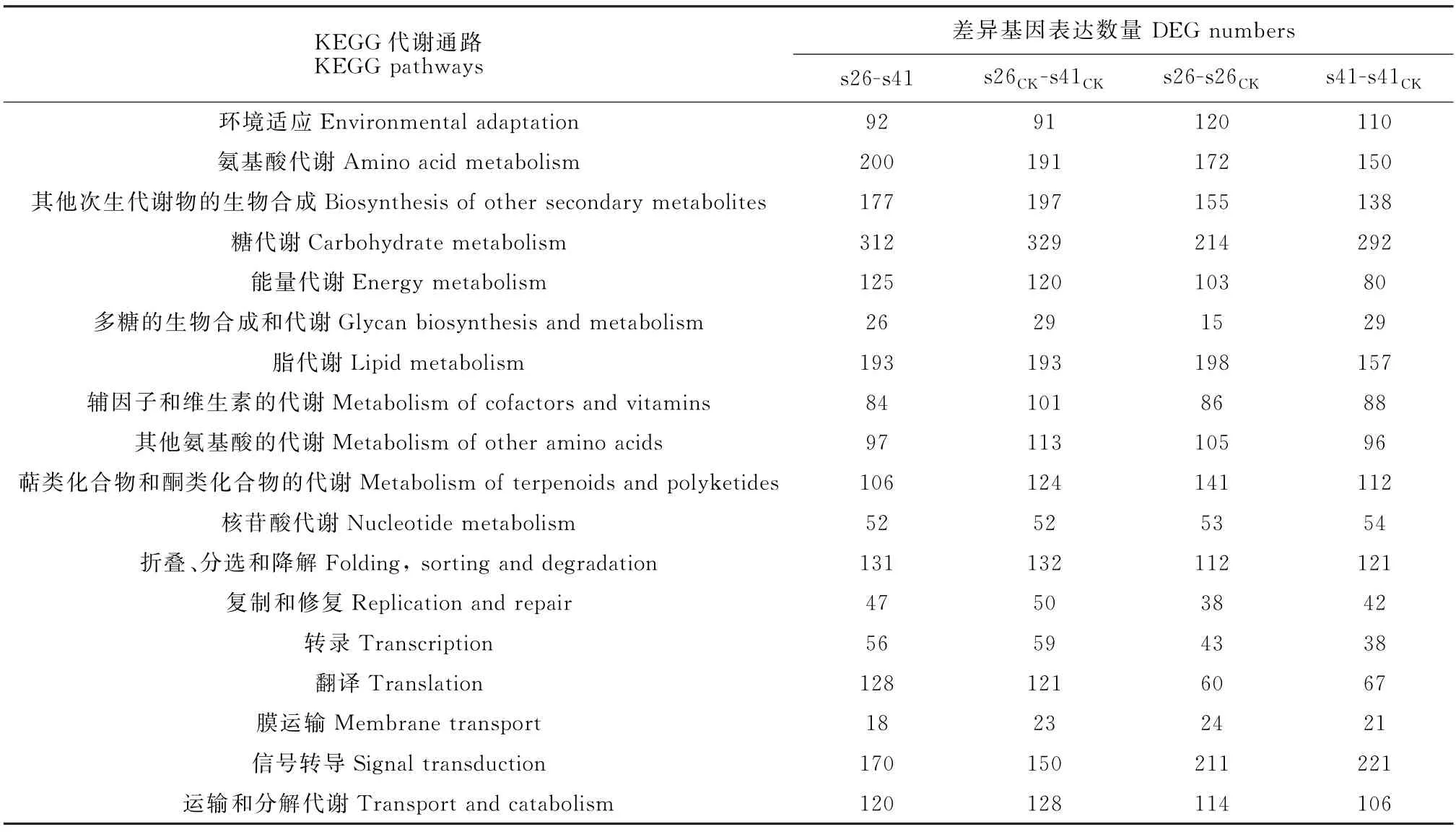
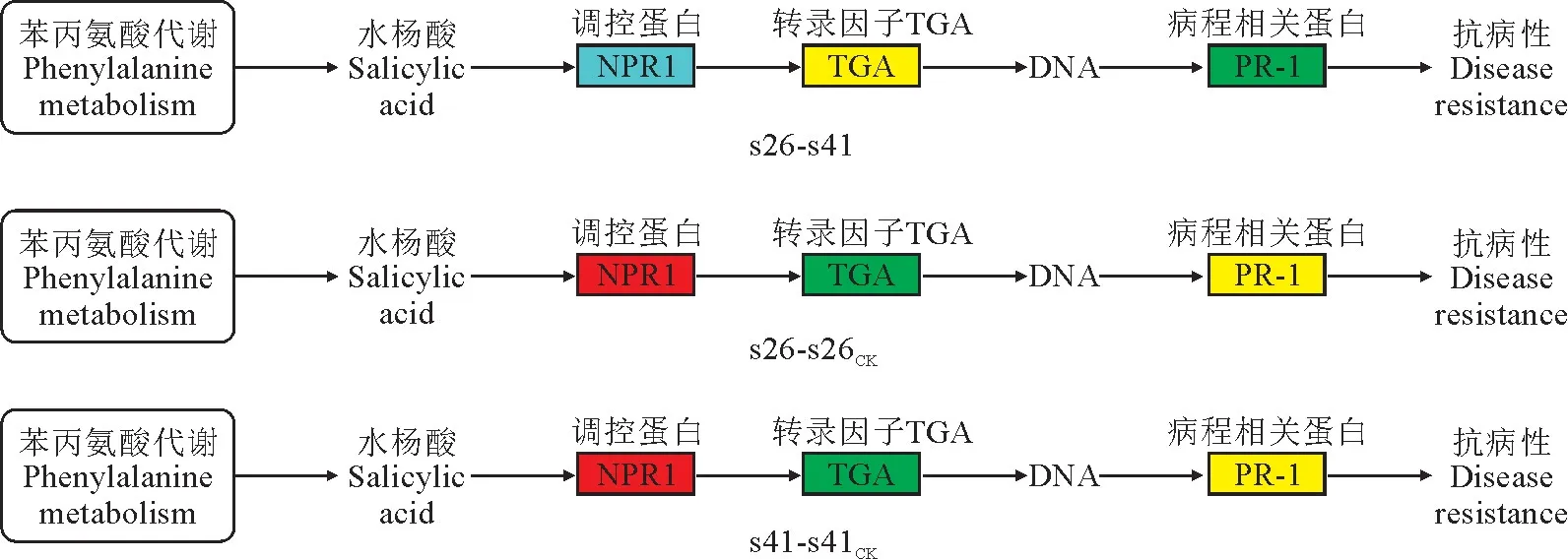
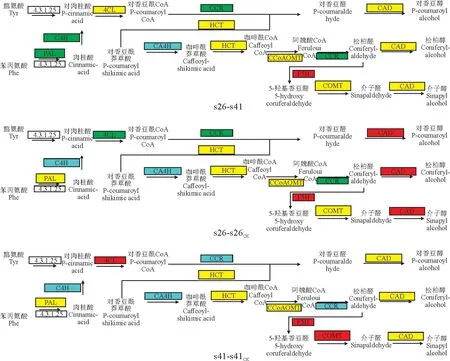
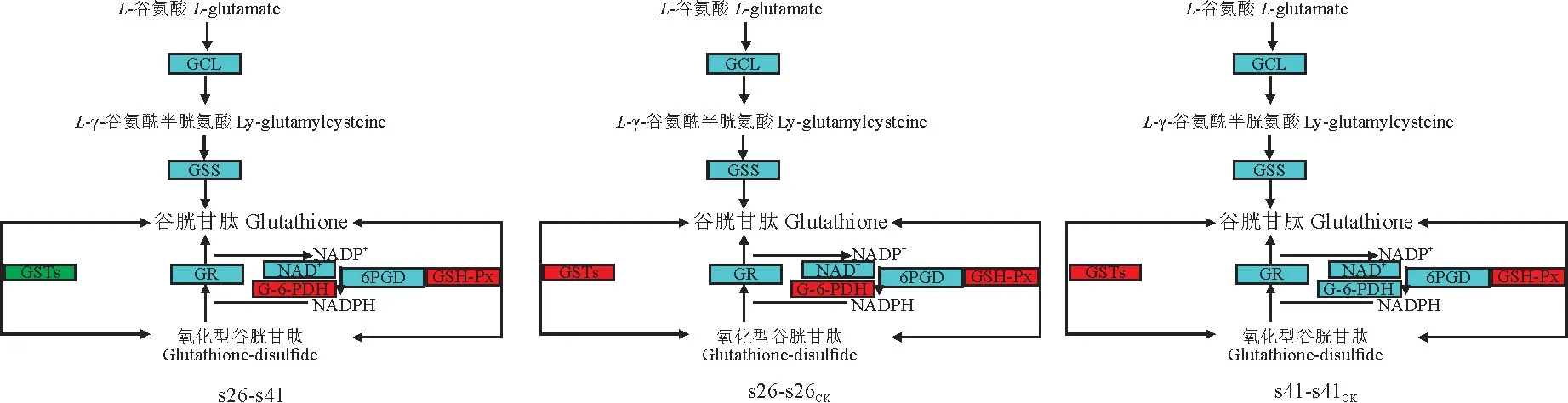
向日葵供试材料分别是抗列当材料s41和高感列当材料s26,其抗性分级依据向日葵抗列当鉴定标准《向日葵品种抗列当等级田间鉴定技术规程》(DB 15/T 1946—2020)[11],并依据技术规程,将向日葵种质资源对列当抗性分为5个等级,分别为免疫[寄生程度(parasitism degree,AD)与寄生率(parasitism rate,F)均 为0]、高 抗(0 试验于2021年7-8月在内蒙古农业大学农学院温室中进行,采用盆栽法种植抗、感列当向日葵材料,盆高18 cm、直径17 cm。准备好培养土(营养土∶蛭石∶砂土=1∶1∶1混合)、接种土(列当种子∶培养土=0.02 g∶500 g混合),先在盆底装1/3盆体积的培养土,再装入1/3盆体积的接种土,而后用培养土装满,用水浇透,备用。选取大小均匀,颗粒饱满的向日葵种子播种在盆中,每个向日葵材料各接种5盆(s26、s41),以不接种列当的5盆为对照组(s26CK、s41CK),4株/盆。 在向日葵长出5对真叶后,分别采集接种组和对照组5盆向日葵全部根系,用蒸馏水洗净,混样后将向日葵根样分为两部分,采用液氮速冻后将其放入-80℃冰箱保存,分别用于转录组测序和生理指标测定,每个指标设置3个生物学重复。 1.3.1 生理生化指标 向日葵根系谷胱甘肽、水杨酸含量及苯丙氨酸解氨酶、肉桂醇脱氢酶、抗坏血酸过氧化物酶活性的测定均采用酶联免疫吸附法,试验步骤按照相关试剂盒说明书进行(江苏酶标生物科技有限公司)。总酚含量检测采用福林比色法,试验步骤参照试剂盒的说明书的方法进行(南京建成生物工程研究所)。 1.3.2 总RNA提取、文库构建及测序 提取样品总RNA(mir VanaTMmiRNA ISOlation Kit,Ambion-1561试剂盒)并使用DNase消化DNA 后,用带有Oligo(d T)的磁珠富集真核生物m RNA;加入打断试剂将mRNA 打断成短片段,以打断后的m RNA 为模板,用六碱基随机引物合成一链cDNA,然后配制二链合成反应体系合成二链cDNA,并使用试剂盒纯化双链cDNA;纯化的双链cDNA 再进行末端修复、加A 尾并连接测序接头,然后进行片段大小选择,最后进行PCR 扩增;构建好的文库用Agilent 2100 Bioanalyzer质检合格后,使用Illumina HiSeq TM 2500进行测序。 1.3.3 测序数据处理 为了得到可用于后续分析的高质量数据(reads),需要对原始数据(raw reads)进行进一步质量过滤。首先使用Trimmomatic[13]软件去除接头,过滤低质量碱基,得到高质量的有效数据(clean reads)。使用Cufflinks[14]软件对基因FPKM[15]表达量定量。计算基因的表达量差异,通过Htseqcount[16]软件获取落到各个样本中基因的reads数目,使用DESeq R package[17]的estimateSizeFactors函数对数据进行标准化,并使用nbinom Test函数计算差异比较的P值和差异倍数。挑选出P<0.05,差异倍数>2的差异基因,进行差异基因的GO和KEGG[18]分析。 列当寄生向日葵后,向日葵的根系形态被破坏(图1)。其中,列当寄生的高感向日葵材料(s26)根系数量明显少于其对照(s26CK)和列当寄生的高抗材料(s41),s41的根系数量与其对照(s41CK)则无明显差别。 图1 高感(s26)、高抗(s41)列当向日葵材料接种和对照根部、植株形态及列当形态Fig.1 Morphology of roots,seedlings,and broomrapes of the resistant(s41)and susceptible(s26)sunflowers under inoculations 向日葵根系被破坏,严重影响其正常生发发育,且列当从向日葵根部吸取水分和营养,导致高感材料(s26)植株高度同时明显低于其对照(s26CK),也明显低于高抗材料(s41),高抗材料(s41)植株高相较于对照(s41CK)则无明显差别。 2.2.1 转录组质量分析 对全部样本进行有参考基因组的转录组测序(表1),共获得85.77 G 的Clean数据,各样本的有效数据量分布在6.56~7.45 G 之间,Q30碱基分布在90.61%~93.63%之间,平均GC含量为45.23%,通过将reads比对到参考基因组上,得到各个样本的基因组比对情况,比对率为77.23%~86.31%,数据显示向日葵根系转录组测序质量良好,可进行后续代谢通路及GO 分析。 表1 转录组质量分析Table 1 Analysis of transcriptome sequencing quality 2.2.2 感、抗材料差异基因表达分析 差异基因筛选分析结果(图2)显示,向日葵感、抗材料接种列当后共筛选出差异表达基因23 930个。其中,感列当材料对照(s26CK)与抗列当材料对照(s41CK)相比,差异基因共有6 362个,其中上调3 073个,下调3 289 个;感、抗列当材料接种列当(s26、s41)后,两者差异基因共有6 609个,其中上调3 294个,下调3 315个;s41与s41CK相比,差异基因共有5 490个,包括上调3 075个,下调2 415个;s26与s26CK相比,差异基因共有5 469个,包括上调3 298个,下调2 171个。s41-s41CK差异基因总数较s26-s26CK多21个(0.38%),上调基因数较s26-s26CK少223个,下调基因数较s26-s26CK多244个。s26-s41差异基因总数较s26CK-s41CK多247个(3.18%),上调基因数和下调基因数分别较s26CK-s41CK多221个和26个。综上表明,接种列当处理相较于对照而言,无论是抗性材料还是高感材料,差异表达基因数中上调基因数均明显多于下调基因数。 图2 接种列当与对照向日葵材料差异表达基因数量Fig.2 Number of differential genes between the inoculated and control sunflowers 2.2.3 差异表达基因GO 功能富集分析 对列当寄生的向日葵根系差异表达基因进行注释和功能富集分析(表2)可知,4个比较组均分为3个主要类型:生物过程(BP)、细胞组分(CC)和分子功能(MF)。其中,s26-s41组富集到的差异基因共有6 452个,上调3 164个,下调3 288个,差异基因注释到GO 分类中,其中生物过程有22个小类别,细胞组分有9个小类别,分子功能有13个小类别;差异基因数量位列前10 的小类别分别为:细胞过程、结合、细胞器、代谢过程、催化活性、膜、刺激响应、生物调节、生物过程的调节、发育过程。s26-s26CK组富集到差异基因共6 455个,上调3 485个,下调2 970个,差异基因注释到GO 分类中,其中生物过程有22个小类别,细胞组分有9个小类别,分子功能有13个小类别;差异基因数量位列前10的小类别分别为细胞过程、结合、细胞器、代谢过程、催化活性、膜、刺激响应、生物调节、生物过程的调节、发育过程。s41-s41CK组富集到差异基因共6 322个,上调3 407个,下调2 915个,差异基因注释到GO分类中,其中生物过程有22个小类别,细胞组分有9个小类别,分子功能有13个小类别;差异基因数量位列前10的小类别分别为细胞过程、细胞器、结合、代谢过程、催化活性、膜、刺激响应、生物调节、生物过程的调节、发育过程。s26CK-s41CK组富集到差异基因共6 565个,上调3 112个,下调3 453个,差异基因注释到GO 分类中,其中生物过程有22个小类别,细胞组分有9个小类别,分子功能有12个小类别;差异基因数量位列前10的小类别分别为细胞过程、细胞器、结合、代谢过程、催化活性、膜、刺激响应、生物调节、生物过程的调节、发育过程。 表2 GO 二级分类数前10位差异表达基因Table 2 The top 10 differentially expressed genes of GO secondary classification 2.2.4 差异表达基因KEGG pathway富集分析 通过对KEGG 代谢途径的分析,差异表达基因总体上被分为细胞过程、环境信息、遗传信息、代谢和生化系统等5大类,进一步可细分为18个子类,即信号转导,膜转运,翻译,转录,复制和修复、折叠,分选和降解,核苷酸代谢,萜类化合物和酮类化合物代谢,辅酶和维生素代谢,其他氨基酸代谢,脂代谢,多糖的合成和代谢,能量代谢,糖代谢,其他次生代谢产物生物合成,氨基酸代谢,环境适应,运输和分解代谢。在接种列当条件下,主要影响着感、抗材料的糖代谢、氨基酸代谢、脂代谢、次生代谢物合成、信号转导、折叠、分选和降解、翻译等过程(表3)。 表3 差异基因KEGG pathway分类Table 3 Differential genes from KEGG pathway 2.2.5 代谢通路分析 在水杨酸代谢途径(图3)中,接种列当后向日葵根系通过苯丙氨酸代谢途径生成水杨酸,影响NPR1蛋白相关基因、TGA 家族相关基因和病程相关蛋白PR-1基因表达,表现为植物的抗病性。s26相较于s26CK和s41相较于s41CK,水杨酸代谢途径编码NPR1蛋白相关基因表达均上调、转录因子TGA家族相关基因表达均下调,病程相关蛋白PR-1基因既有上调又有下调。s26与s41相比,编码NPR1基因表达无显著变化,TGA 家族相关基因表达均有上调又有下调,而病程相关蛋白PR-1基因表达呈下调。 KEGG通路图上红色表示上调基因,绿色表示下调基因,黄色表示对应的基因中既有上调也有下调。下同。图3 水杨酸代谢途径Red color on the KEGG pathway map indicates up-regulated genes,green color indicates down-regulated genes,and yellow color indicates both up and down regulation in the corresponding genes.The same as below.Fig.3 Metabolic pathwaies of the alicylic acid 同时,在木质素合成途径(图4)中,由酪氨酸和苯丙氨酸经过一系酶催化生成对香豆醇、松柏醇、芥子醇3种木质素的单体。苯丙氨酸解氨酶作为苯丙氨酸代谢的第一步,将苯丙氨酸在苯丙氨酸解氨酶作用下转化为反式肉桂酸。其中,s26与s41相比,差异基因在第一步反应均表现为下调,表明s41差异基因表达量更高;s41与s41CK相比,苯丙氨酸解氨酶基因的表达既有上调也有下调;s26与s26CK相比,苯丙氨酸解氨酶基因的表达既有上调也有下调。肉桂醇脱氢酶作为合成木质素的关键酶,参与形成木质素的单体反应。s26与s26CK相比,肉桂醇脱氢酶的基因表达为上调,而s41与s41CK相比及s26与s41相比较肉桂醇脱氢酶基因的表达均为既有上调又有下调。 图4 木质素合成途径Fig.4 Lignin synthesis pathway 另外,在谷胱甘肽合成途径(图5)中,通过L-谷氨酸形成谷胱甘肽,并在多种酶的催化下完成氧化型与还原型之间转化。其中,谷氨酰半胱氨酸连接酶、谷胱甘肽合成酶的基因表达均无变化。在还原型谷胱甘肽和氧化型谷胱甘肽转化之中,s41材料谷胱甘肽-S-转移酶、谷胱甘肽过氧化物酶基因表达量上调;s26材料在谷胱甘肽-S-转移酶、6-磷酸葡萄糖-1-脱氢酶、谷胱甘肽过氧化物酶基因表达量上调;s26与s41 相比,6-磷酸葡萄糖-1-脱氢酶、谷胱甘肽过氧化物酶基因表达量上调,谷胱甘肽-S-转移酶基因下调。 图5 谷胱甘肽合成途径Fig.5 Glutathione synthesis pathway 高抗和高感向日葵材料根系生理指标在接种列当条件下均不同程度地高于相应对照(图6)。 其中,s41根系谷胱甘肽含量、苯丙氨酸解氨酶活性、水杨酸含量和总酚含量较s41CK分别显著提高37.00%、118.01%、96.36%和5.82%;s26根系谷胱甘肽含量、水杨酸含量以及苯丙氨酸解氨酶、肉桂醇脱氢酶、抗坏血酸过氧化物氧酶活性相较于s26CK分别显著提高45.50%、108.33%、17.61%、9.60%和10.49%;s26根系的谷胱甘肽含量及苯丙氨酸解氨酶、肉桂醇脱氢酶、抗坏血酸过氧化物氧酶活性比s41分别显著提高99.06%、99.35%、52.76%和17.89%,而s41根系水杨酸含量却较s26显著提高52.76%。以上结果说明,向日葵感、抗材料各生理指标在列当寄生下均有积极响应,高感材料虽无法抵御列当的寄生,但通过提高水杨酸和谷胱甘肽含量来帮助维持植株体内代谢平衡,以减轻列当对向日葵植株的危害;抗性材料通过提高苯丙烷类代谢、增强抗氧化酶活性、增加水杨酸含量来增强对列当的抗性。 当植物受到逆境胁迫时,植物会产生多种应激反应,最终表现为胁迫抗性和胁迫耐性,这些适应和应答可以提高植物在逆境下的适应性。列当是向日葵生长和生产的主要限制因素之一[19]。由于列当的侵染导致严重的氧化损伤和产量损失,向日葵会发生许多生理和生化响应[6]。为了应对列当的寄生,寄主植物实施了各种策略,包括利用抗氧化剂、诱导细胞壁强化、产生茉莉酸(JA)或水杨酸(SA),以对抗侵染或杀死列当[20-21]。 苯丙烷代谢是植物次级代谢中的重要途径,苯丙氨酸解氨酶(PAL)作为苯丙烷代谢途径中的第一种酶,在苯丙烷代谢中占据着重要的地位,其产物是多种次级代谢产物的直接或间接前体,中间产物经过一系列转化生成如类黄酮、木质素、生物碱等次生代谢产物,这些次级产物对植物响应胁迫、抗逆防御、提高抗氧化能力等众多方面具有非常重要的意义[22]。肉桂醇脱氢酶(CAD)在木质素合成过程中具有关键性作用。肉桂醇脱氢酶活力降低,会引起木质素含量的减少[23]。本试验中向日葵列当抗性材料根系PAL活性比高感材料低,但活性在接种前后变化幅度很大。通过比较分析感、抗材料代谢通路发现,PAL参与苯丙氨酸代谢中生成反式肉桂酸相关基因表达,高感材料较高抗材料表现为下调,说明抗性材料PAL 基因表达量更高,这与抗性材料PAL活性在接种列当后大幅提升得到互相验证。在接种列当条件下,抗、感向日葵材料CAD 活性均有升高,但高感材料活性升高更显著,可能是由于高感材料的CAD 基因表达表现为上调,CAD 基因大量表达提了高感材料CAD 的活性,高抗材料的CAD基因表达则是既有上调又有下调,因此高抗材料相应酶活性改变不显著。 活性氧(reactive oxygen species,ROS)急剧增加是植物面临各种逆境胁迫时的一个共同表现,表明ROS 在植物逆境响应中起着十分重要的作用[24]。张默靖等[25]和Li等[26]研究表明,向日葵被列当寄生后会产生大量的ROS,导致膜脂过氧化,细胞膜损伤。抗坏血酸-谷胱甘肽循环(ASA-GSH)是植物抗氧化系统中最重要的防御机制,抗坏血酸过氧化物酶是ASA-GSH 循环中的一种关键酶,具有将H2O2转化为水的功能,是保护植物细胞免受超氧化物氧化胁迫的重要酶类之一。谷胱甘肽含量和抗坏血酸过氧化物酶活性的增加,可以有效清除向日葵体内的大量ROS,防止向日葵细胞膜损伤,达到保护向日葵植株的效果。Yang 等[27]研究表明,ROS清除机制在感抗材料被寄生后都被激活,但作用不同;与对照相比,在接种列当后,抗氧化酶在向日葵中积累并且它们的活性显著增加,本研究结果与之相同。此外,谷胱甘肽-S-转移酶(GST)被认为是抗性向日葵响应列当寄生的ROS 解毒机制[28],在本研究的谷胱甘肽代谢通路中,向日葵列当抗性材料转移酶基因表达量更高,表明抗性材料调节体内ROS代谢平衡的能力更强。酚类物质是植物中最重要、分布和研究最广泛的次生代谢物质,可参与植物生殖发育,提高抗逆性,且总酚含量增加有助于保护植物免受氧化损伤[29]。张桂伟等[30]发现总酚、总黄酮含量与葡萄柚抗氧化能力呈显著的正相关关系。本研究中向日葵抗性材料总酚含量的变化呈显著性水平,由此可知,向日葵列当抗性材料可能具有更强的抗氧化能力。 水杨酸(SA)是植物的内源性信号分子,与植物的多种抗性有关。在拟南芥中,NPR1介导了SA诱导的发病相关基因(PRs)和系统获得性抗性的表达。NPR1是SA 信号通路中的关键因子,NPR1、NPR3和NPR4在SA 介导的植物抗病性中起关键作用[31]。本研究中向日葵列当抗、感材料水杨酸含量在接种后均上升显著;在水杨酸代谢通路中,抗、感材料相较于各自对照,NPR1表达均呈现上调,而在病程相关蛋白的基因表达上抗性材料高于高感材料,表明抗性材料通过提高调控蛋白、病程相关蛋白的基因表达,进而提高向日葵对列当的抗性。此外,Saidi等[32]研究表明,SA 可以直接作为一种抗氧化剂来清除ROS或通过激活抗氧化反应间接调节氧化还原平衡。 列当寄生对向日葵根系形态有明显的破坏作用,从而对其正常生长和发育产生严重影响;差异表达基因富集分析发现,向日葵差异表达基因数量位列前10的小类别基本一致,主要包括细胞过程、结合、细胞器、代谢过程、催化活性、膜等,表明列当寄生下的向日葵差异表达基因主要涉及这些方面。进一步分析其代谢通路发现,接种列当后向日葵根系通过苯丙氨酸代谢途径生成水杨酸,影响了NPR1蛋白相关基因、TGA家族相关基因和病程相关蛋白PR-1基因的表达,从而表现出植物的抗病性,同时也使向日葵根系木质素合成途径和谷胱甘肽合成途径受到影响。分析根系生理指标变化发现,在列当寄生下高抗和高感向日葵材料的谷胱甘肽含量、苯丙氨酸解氨酶活性、水杨酸含量以及总酚含量均表现出不同程度增加,表明这些生理指标在提高向日葵对列当抗性方面有一定的积极作用。1.2 试验设计
1.3 观测指标及方法
2 结果与分析
2.1 接种列当对向日葵根系和植株形态的影响

2.2 接种列当向日葵根系转录组分析
2.3 接种列当对向日葵根系生理指标的影响
3 讨论
4 结论
