miR-10b靶向TGFBR1/SMAD3通路对特发性矮小症的软骨细胞增殖、肥大的影响机制
2024-02-18胡娜李正宇叶春风吴英姚庆黄世祥李文朱海琴
胡娜 李正宇 叶春风 吴英 姚庆 黄世祥 李文 朱海琴
摘要:目的 研究miR-10b對特发性矮小症(ISS)的影响及作用机制。方法 收集ISS患儿(ISS组)和体检健康儿童(健康对照组)各54例,qPCR检验血清miR-10b表达量,分析ISS组患儿血清miR-10b表达与患儿临床资料的关系。采用miR-10b inhibitor、si-TGFBR1及各自阴性对照转染C28/I2细胞,利用CCK-8实验检测C28/I2细胞增殖能力,Western blot检测侏儒相关转录因子2(RUNX2)、X型胶原α1链(COL10A1)、转化生长因子β受体1(TGFBR1)、SMAD3、pSMAD3蛋白表达量。在StarBase数据库筛选miR-10b靶点,利用双萤光素酶报告基因实验验证miR-10b与TGFBR1的靶向关系。结果 ISS组血清miR-10b表达量高于健康对照组,且miR-10b表达越高,患儿的身高、IGF-1、骨特异性碱性磷酸酶的下降越明显(P<0.05)。与NC组相比,miR-10b inhibitor组细胞增殖能力升高,RUNX2、COL10A1、TGFBR1、pSMAD3蛋白表达上调(P<0.05);StarBase数据库提示miR-10b存在TGFBR1的结合位点,双萤光素酶报告基因实验证实两者结合。与si-NC相比,si-TGFBR1组TGFBR1表达量下调,细胞增殖能力下降(P<0.05)。结论 miR-10b通过靶向TGFBR1/SMAD3通路在特发性矮小症中抑制软骨细胞的增殖、肥大。
关键词:特发性矮小症;受体,转化生长因子βⅠ型;miR-10b;Smad3蛋白质
中图分类号:R752.8文献标志码:ADOI:10.11958/20230659
The mechanism of miR-10b targeting TGFBR1/SMAD3 pathway on chondrocyte proliferation and hypertrophy in idiopathic short stature
Abstract: Objective To investigate the effect and mechanism of microRNA-10b (miR-10b) on idiopathic short stature (ISS). Methods A total of 54 children with ISS and 54 healthy children were collected. The serum expression of miR-10b was detected by RT-qPCR, and the relationship between serum miR-10b expression and clinical data of children with ISS was analyzed. miR-10b inhibitor, si-TGFBR1 and their negative control transfection C28/I2 cells were used. CCK-8 experimental detection was used to detect C28/I2 cell proliferation. Western blot assay was used to detect gnome related transcription factor 2 (RUNX2), collagen type X alpha 1 chain (COL10A1), transforming growth factor beta receptor 1 (TGFBR1), SMAD3 and pSMAD3 protein expression. The target of miR-10b was screened in StarBase database, and the targeting relationship between miR-10b and TGFBR1 was verified by dual luciferase reporter gene assay. Results The serum expression of miR-10b was higher in the ISS group than that of the healthy control group, and the higher the miR-10b expression, the more obvious the decrease of child height, IGF-1 and alkaline phosphatase (P<0.05). Compared with the NC group, the cell proliferation ability and RUNX2, COL10A1, TGFBR1, and pSMAD3 protein expression were up-regulated in the miR-10b inhibitor group (P<0.05). StarBase database suggested that miR-10b had a binding site of TGFBR1, and dual luciferase reporter gene assay confirmed that TGFBR1 interacted with miR-10b (P<0.05). Compared with the si-NC group, the expression of TGFBR1 was down-regulated and the cell proliferation ability was decreased in the si-TGFBR1 group (P<0.05). Conclusion miR-10b inhibits chondrocyte proliferation and hypertrophy in idiopathic short stature by targeting TGFBR1/SMAD3 pathway.
Key words: idiopathic short stature; receptor, transforming growth factor-beta type Ⅰ; miR-10b; Smad3 protein
特发性矮小(idiopathic short stature,ISS)是矮小症中最常见的类型,即患者生长激素水平正常,而身高在不伴有潜在病理状态下低于同龄、同性别人群身高平均值2个标准差(SD)[1]。ISS病因不明,占所有身材矮小儿童的60%~80%,严重影响儿童生长发育与生活质量,给心理健康造成极大影响,早发现、早诊断对改善患儿终身高至关重要。近年来,非编码RNA生物标志物研究成为热点。研究表明,血液中微小RNA(microRNA,miRNA)在不同生理、病理状态下具有特定的表达水平和模式,稳定性、重复性较好,有助于疾病的早期诊断[2-3]。转化生长因子β(TGF-β)信号通路在胚胎骨发育和出生后骨稳态中都有重要作用,与早期胚胎发育和器官发生、免疫监督、组织修复密切相关[4]。本课题组前期预试验发现miR-10b在ISS儿童血清中上调表达量最高,本文进一步研究miR-10b与ISS的关系及作用机制。
1 资料与方法
1.1 一般资料 选取2019年8月—2021年8月在我院接受治疗的54例ISS患者(ISS组)。纳入标准:(1)身高低于同性别、同年龄、同地区、同种族正常参考值平均数-2 SD。(2)出生时身长和体质量正常,而且身材匀称。(3)无明显的慢性器质性疾病(肝、肾、心、肺、内分泌代谢病和骨骼发育障碍)。(4)无心理和严重的情感障碍,饮食正常。(5)生长速率稍慢或正常,一般每年生长速率<5 cm。(6)染色体正常。(7)生长激素(GH)激发试验GH峰值≥10 μg/L,且胰岛素样生长因子1(IGF-1)浓度正常。(8)骨龄正常或延迟。排除标准:(1)资料不全者。(2)伴有系统性慢性疾病,如先天性心脏病、哮喘、营养不良等疾病者。ISS组男30例,女24例,年龄5~12岁,平均(8.77±0.35)岁,平均身高(122.3±11.42)cm。选取同期进行体检的健康儿童54例(健康对照组),男29例,女25例,年龄5~12岁,平均(8.69±0.46)岁,平均身高(133.2±10.61)cm,2组性别(χ2=0.037)、年龄(t=1.017)比较差异无统计学意义(均P>0.05)。ISS组身高较健康对照组下降(t=37.311,P<0.01)。本研究经我院伦理委员会审核并批准(2019医研伦审第10号),患儿或家属均知情并签署研究同意书。所有对象于入院后首次检查时采集血液样本,采集后立即在液氮中冷冻,-80 °C的冰箱中保存待用。
1.2 细胞及试剂 人软骨细胞系C28/I2、293T细胞购自中国科学院典型培养物保藏委员会细胞库。DMEM/F12细胞培养基、胎牛血清(FBS)和链霉素-青霉素双抗均购自美国Gibco公司。TGF-β受体1(TGFBR1)、重组人SMAD家族成员3(SMAD3)、磷酸化SMAD3(pSMAD3)、X型胶原α1链(COL10A1)、侏儒相关转录因子2(RUNX2)、甘油醛-3-磷酸脱氢酶(GAPDH)抗体、辣根过氧化物酶(HRP)标记的兔抗IgG二抗均购自英国Abcam公司,红细胞裂解液购自北京索莱宝科技有限公司。BeyoRTTM II cDNA第一链合成试剂盒及ECL发光试剂购自碧云天生物技术有限公司。TaqMan? Universal PCR Master Mix、TaqMan miRNA特异性引物、pmirGLO-TGFBR1萤光素酶载体(野生型或突变型质粒)均购自深圳华大基因有限公司,StepOne Plus实时荧光定量PCR仪购自上海凌仪生物科技有限公司。Lipofectamine 3000 购自Invitrogen公司。miR-10b抑制物(miR-10b inhibitor)、模拟物(miR-10b mimic)及其对照(miR-NC)、TGFBR1小干扰RNA(si-TGFBR1)及其阴性对照(si-NC)均购自广州锐博生物科技有限公司,DAB显色剂购自武汉博士德公司。双萤光素酶报告基因购自美国Promega公司。IGF?1、骨特异性碱性磷酸酶(BAP)酶联免疫吸附试验(ELISA)试剂盒购于英国Immunodiagnostic systems公司。
1.3 方法
1.3.1 人血miR-10b检测 利用3倍体积的红细胞裂解液与血液样本混匀,3 000 r/min离心10 min后移除上清液,可見白色颗粒沉于管底。随后利用Trizol试剂提取总RNA,使用TaqMan MicroRNA逆转录试剂盒将总RNA进行逆转录,使用SYBR Green PCR Master Mix进行实时荧光定量PCR(qPCR),以U6基因作为内参。20 μL反应体系:SYBR? Green Realtime PCR Master Mix 10 μL,上、下游引物各1 μL,cDNA 2 μL,ddH2O 6 μL。反应条件:95 ℃ 30 s;95 ℃ 5 s,55℃ 45 s,72 ℃ 30 s,40个循环。
1.3.2 血清IGF-1和BAP检测 取ISS组空腹静脉血5 mL,5 000 r/min离心10 min后取血清,采用ELISA法检测IGF-1和BAP表达水平,检测步骤按照试剂盒说明书进行操作。
1.3.3 细胞培养 使用添加10%胎牛血清和1%青霉素-链霉素的DMEM/F12培养基培养人软骨细胞系C28/I2,将细胞置于5%CO2和37 ℃恒温培养箱中培养。利用胰蛋白酶按照1∶2的比例进行细胞传代。
1.3.4 细胞转染 取对数生长期C28/I2细胞,消化后制成单细胞悬液,将2×104个/mL的单细胞悬液接种至6孔板内。待软骨细胞融合度达到30%~40%时,使用Lipofectamine 3000行瞬时转染miR-10b inhibitor、miR-NC、si-TGFBR1及si-NC,分别为miR-10b inhibitor组、NC组、si-TGFBR1组、si-NC组。
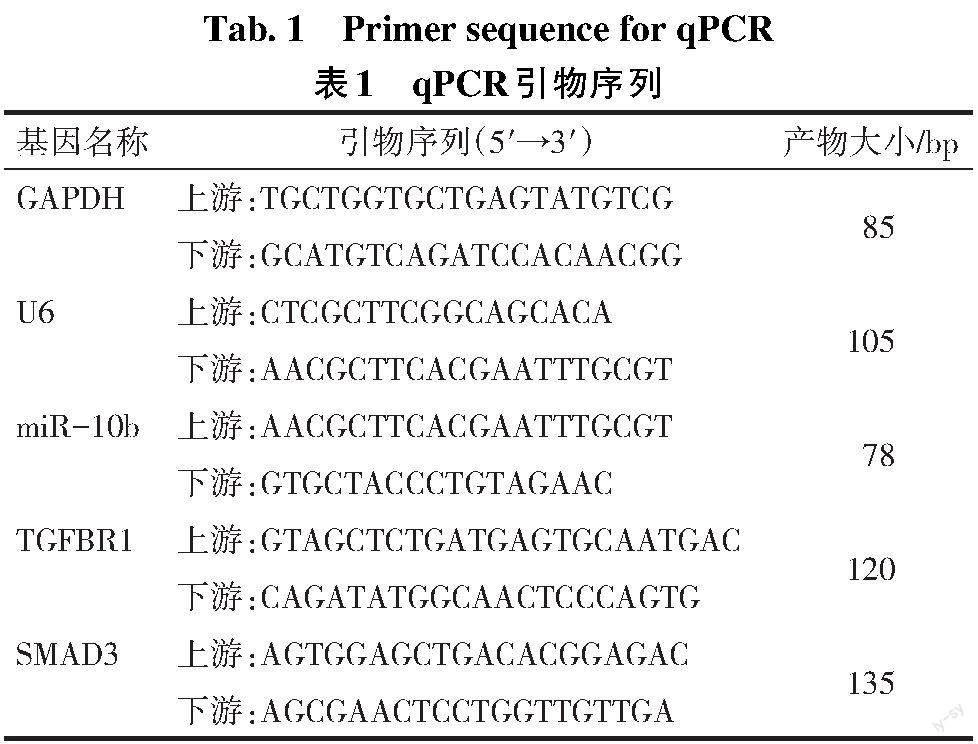
1.3.5 qPCR检测基因表达 利用Trizol试剂提取细胞中总RNA,参照1.3.1进行qPCR检测miR-10b表达,以U6基因作为内参。另使用BeyoRTTM II cDNA第一链合成试剂盒逆转得到cDNA,以GAPDH为内参,使用TaqMan? Universal PCR Master Mix进行qPCR,检测TGFBR1、pSMAD3的表达水平。反应体系及反应条件同方法1.3.1,以2-ΔΔct法计算目的基因的表达量,引物序列由华大基因公司合成,见表1。
1.3.6 CCK-8实验检测细胞增殖 4组细胞接种于96孔板(每孔1×103个细胞),加入培养基正常培养,各时间点(0、24、48、72 h)均于37 ℃下加入CCK-8试剂 1 h。在450 nm波长处检测光密度(OD)值。
1.3.7 Western blot检测蛋白表达 采用RIPA试剂提取细胞总蛋白,BCA法进行蛋白定量。蛋白质经10% SDS-PAGE分离,电转移至PVDF膜。用5%脱脂奶粉孵育1 h,TBST洗涤5次后滴加TGFBR1、SMAD3、pSMAD3、RUNX2、COL10A1、GAPDH一抗(均为1∶1 000稀释),于4 ℃孵育过夜。与HRP标记的兔抗IgG(1∶5 000)室温孵育1 h,TBST洗涤5次后用ECL发光试剂检测蛋白条带。以GAPDH为内参,使用Image J软件分析目的蛋白相对表达量。
1.3.8 miR-10b与TGFBR1靶向关系的预测与验证 使用StarBase v2.0数据库(https://starbase.sysu.edu.cn/index.php)预测miR-10b与TGFBR1的结合位点;进行双萤光素酶报告基因测定确认TGFBR1和miR-10b之间的结合关系。在12孔板中,1×105个细胞悬液培养至40%,利用Lipofectamine 2000将miR-10b模拟物或NC(50 nmol/L)和1 μg pmirGLO-TGFBR1萤光素酶载体(野生型或突变型质粒)共转染293T细胞。在转染后24 h进行萤光素酶活性检测。细胞分组:TGFBR1 3?-UTR-WT+miR-10b mimic/NC、TGFBR1 3?-UTR-MUT+ miR-10b mimic/NC。
1.4 统计学方法 采用GraphPad Prism 8软件进行数据分析。符合正态分布的计量资料以[x] ±s表示,2组间均数比较采用独立样本t检验,P<0.05为差异有统计学意义。
2 结果
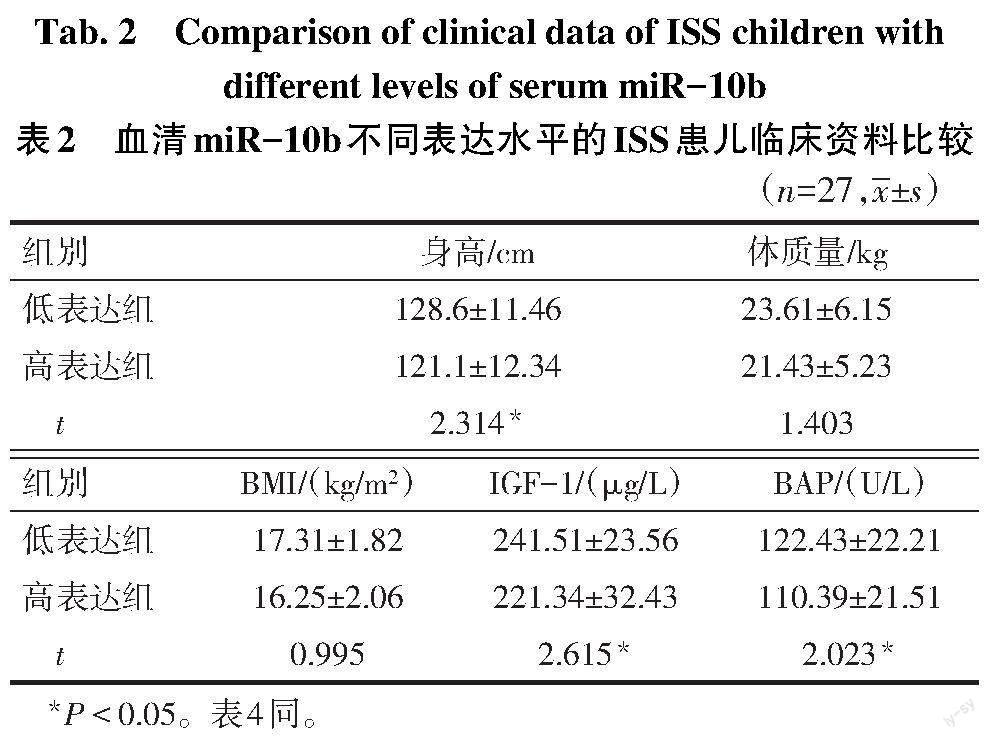
2.1 miR-10b在ISS中的表达 qPCR结果表明,ISS组血清miR-10b表达量高于健康对照组(2.68±0.25 vs. 1.12±0.21,t=35.111,P<0.01)。以miR-10b中位表达量2.54为界,将ISS组分为miR-10b高表达组(≥2.54)和低表达组(<2.54)各27例。
2.2 ISS组患儿血清miR-10b表达与患儿临床资料的关系 miR-10b高、低表达组患儿体质量和BMI差异均无统计学意义(P>0.05);miR-10b低表达组身高、IGF-1、BAP高于高表达组(P<0.05)。见表2。
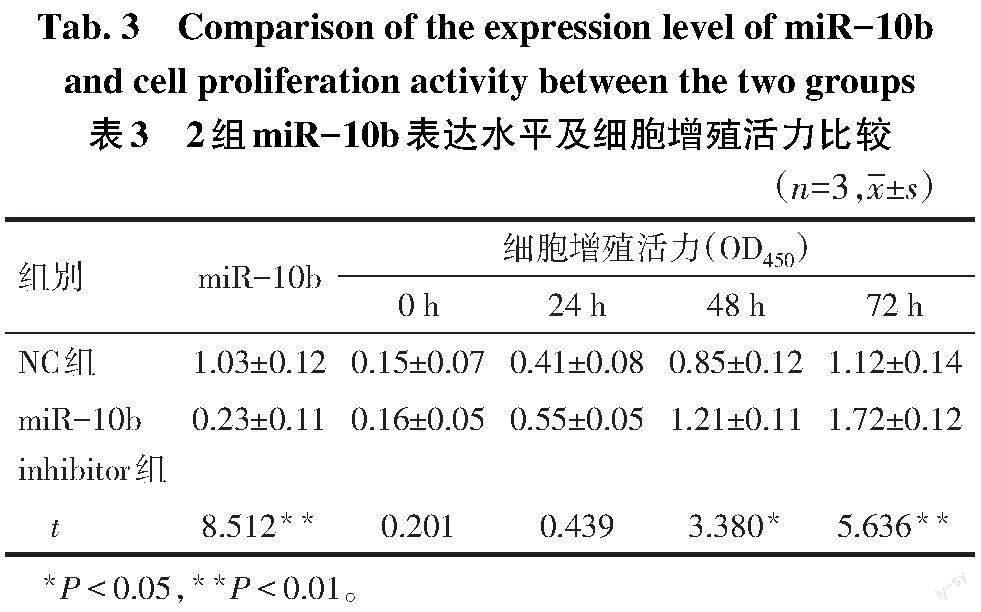
2.3 抑制miR-10b表达促进软骨细胞的增殖、肥大 qPCR结果表明,miR-10b inhibitor组miR-10b表达量低于NC组。CCK-8实验结果显示,抑制miR-10b后48、72 h,C28/I2软骨细胞增殖能力较NC组升高(P<0.05),见表3。Western blot结果显示,抑制miR-10b表达后,RUNX2及COL10A1蛋白表达量上调(P<0.05),见图1。
2.4 miR-10b调控TGFBR1/SMAD3通路 Western blot结果显示,与NC组相比,抑制miR-10b表达后TGFBR1、pSMAD3表达上调(P<0.01),见图2。
2.5 miR-10b靶向调控TGFBR1 StarBase数据库分析发现,miR-10b能与TGFBR1 mRNA的3'-UTR结合,见图3A。双萤光素酶报告实验表明,在TGFBR1 WT中,与NC组相比,miR-10b mimic组萤光素酶活性降低(P<0.05),在TGFBR1 MUT中,2组间差异无统计学意义,见图3B。
2.6 敲低TGFBR1表达可抑制C28/I2细胞增殖 qPCR结果表明,si-TGFBR1组TGFBR1表达量低于si-NC组,TGFBR1低表达模型构建成功。CCK-8结果显示,抑制TGFBR1表达72 h后,C28/I2细胞增殖能力较si-NC组降低(P<0.05),见表4。
3 讨论
ISS是一种内分泌相关疾病,诊断困难,发病机制不明,其发病原因与遗传、骨骼发育障碍等多因素有关,已成为严重影响儿童身心健康的疾病之一[5]。目前,ISS的主要治疗方法是重组人生长激素,但ISS患者并不表现生长激素缺乏,因此重组人生长激素对ISS患者疗效不一。同时,临床上使用大剂量重组人生长激素还会引起高血糖、高胰岛素血症及骨和软骨瘤等不良反应[6]。因此,进一步阐明ISS的发病机制,探讨ISS生物标志物是当前临床关注的重点,做到早发现、早诊断,以期降低ISS患病率或提高ISS患者的生活质量。
ISS患者的主要临床症状为生长迟缓,具体表现为身高、体质量、骨骼生长、发育异常。骨骼发育主要与骨纵向生长有关,研究表明纵向骨主要与生长板驱动有关,软骨细胞在生长板内经历细胞增殖、分化肥大,进而转化为骨,影响骨的纵向生长[7]。目前,非编码RNA的稳定性、特异性已引起临床广泛关注,在肿瘤、炎症、发育异常等各类疾病中发挥作用[8]。例如,LncRNA MALAT-1通过吸附miR-22促進上皮性卵巢癌的生长和转移[9],下调miR-21是克服结肠癌、肝癌等癌症耐药的一种有前景的策略[10]。非编码RNA可调控软骨细胞的增殖和凋亡[11-12],从而可能影响骨的纵向生长。lncRNA对骨代谢平衡有重要调控作用,可通过多种信号通路和转录因子影响间充质干细胞成骨分化过程[13]。miR-143可以调节生长板的损伤[14]。既往研究发现,miR-10b过表达可以抑制乳腺癌[15]、胃癌[16]细胞株的生长。另有研究发现[17],miR-10b-5p可促进成肌细胞增殖,抑制肌纤维形成,但关于miR-10b对ISS的影响尚未见报道。笔者前期预试验通过对ISS患者和健康人标本进行高通量测序,发现miR-10b在ISS患者血清中高表达,但其具体发病机制尚不清楚。在本研究中,进一步扩大患者样本量,采用RT-qPCR进一步证实了miR-10b在ISS患者组的表达量较健康对照组上调,提示miR-10b可能与ISS的发生和发展密切相关,是一个潜在的诊断生物标志物。IGF?1与营养、骨代谢密切相关,且在青少年时期随年龄增长和身体发育而增高,BAP主要由活跃的成骨细胞释放,其水平升高表示骨代谢增加。本研究中ISS患儿miR-10b表达越高,IGF?1、BAP表达量越低,表明miR-10b与骨代谢、发育有关。骨是体内最具活力的器官,通过骨的形成和吸收不断进行重塑。一系列的分子和途径导致成骨细胞分化,而成骨分化又归因于骨生成或骨形成。RUNX2是一种对成骨细胞分化和软骨细胞成熟至关重要的转录因子[18]。在成骨细胞分化过程中,RUNX2在未定向间充质细胞中弱表达,在前成骨细胞中表达上调,在未成熟成骨细胞中达到最高水平,在成熟成骨细胞中表达下调。近年来发现,许多miRNA可调控RUNX2的表达或活性,从而影响成骨过程。如miR-135和miR-203靶向RUNX2会抑制乳腺癌和转移性骨病的进展[19]。本研究发现,抑制miR-10b表达后,RUNX2与COL10A1表达量较NC组升高,提示敲除miR-10b显著促进了软骨细胞的增殖、肥大。
為探讨miR-10b的潜在作用机制,笔者进一步预测并证明TGFBR1基因是miR-10b的直接靶基因。TGFBR1是TGF-β/SMAD通路的重要组成部分,可以将TGF-β信号从细胞表面转导到细胞质中,这一过程对于软骨细胞的增殖和分化是必不可少的。TGF-β作为一种多功能多肽细胞因子,在早期胚胎发育和成人体内稳态中起着至关重要的作用。SMAD蛋白不仅是信号转导蛋白,而且是介导多种信号通路的转录调节因子,如TGF-β、BMP和Wnt信号通路。TGF-β在与Ⅱ型受体(TGFBR2)结合后,会将TGFBR1募集到高度保守的近膜区域;然后激活的TGFBR1会磷酸化其下游靶标,包括信号转导子SMAD家族成员SMAD2和SMAD3[20]。Xiao等[21]发现TGF-β/SMAD信号通路通过调控miR-455-5p/RUNX2轴抑制icmt诱导的终板软骨细胞退变。本研究发现敲低TGFBR1的表达可抑制软骨细胞的增殖,另外抑制miR-10b表达可上调TGFBR1蛋白及SMAD3的磷酸化水平,表明miR-10b可能通过靶向调控TGFBR1表达抑制SMAD3信号通路,从而调节软骨细胞的增殖、肥大。
参考文献
[1] LIU X,DU Z,YI X,et al. Circular RNA circANAPC2 mediates the impairment of endochondral ossification by miR-874-3p/SMAD3 signalling pathway in idiopathic short stature[J]. J Cell Mol Med,2021,25(7):3408-3426. doi:10.1111/jcmm.16419.
[2] HILL M,TRAN N. MiRNA interplay:mechanisms and consequences in cancer[J]. Dis Model Mech,2021,14(4):dmm047662. doi:10.1242/dmm.047662.
[3] DIENER C,KELLER A,MEESE E. Emerging concepts of miRNA therapeutics:from cells to clinic[J]. Trends Genet,2022,38(6):613-626. doi:10.1016/j.tig.2022.02.006.
[4] PENG D,FU M,WANG M,et al. Targeting TGF-β signal transduction for fibrosis and cancer therapy[J]. Mol Cancer,2022,21(1):104. doi:10.1186/s12943-022-01569-x.
[5] SAVAGE M O,STORR H L. GH resistrnce is a component of idiopathic short stature:implications for rhGH therapy[J]. Front Endocrinol(Lausanne),2021,12:781044. doi:10.3389/fendo.2021.781044.
[6] VYAS V,MENON R K. Management of short stature:use of growth hormone in GH-Deficient and non-GH-Deficient conditions[J]. Indian J Pediatr,2021,88(12):1203-1208. doi:10.1007/s12098-021-03892-5.
[7] WU Z,YUAN J,LI J,et al. Hsa_circ_0008870 suppresses bone formation of growth plate through inhibition of miR-185-3p/ MAPK1 axis in idiopathic short stature[J]. Front Bioeng Biotechnol,2022,10:1022830. doi:10.3389/fbioe.2022.1022830.
[8] DU X,ZHANG J,WANG J,et al. Role of miRNA in lung cancer-potential biomarkers and therapies[J]. Curr Pharm Des,2018,23(39):5997-6010. doi:10.2174/1381612823666170714150118.
[9] PEI C,GONG X,ZHANG Y. LncRNA MALAT-1 promotes growth and metastasis of epithelial ovarian cancer via sponging microRNA-22[J]. Am J Transl Res,2020,12(11):6977-6987.
[10] AKHTARKHAVARI T,BAHRAMI A R,MATIN M M. Downregulation of miR-21 as a promising strategy to overcome drug resistance in cancer[J]. Eur J Pharmacol,2022,932:175233. doi:10.1016/j.ejphar.2022.175233.
[11] ZHANG Y,WANG F,CHEN G,et al. LncRNA MALAT1 promotes osteoarthritis by modulating miR-150-5p/AKT3 axis[J]. Cell Biosci,2019,9:54. doi:10.1186/s13578-019-0302-2.
[12] IAQUINTA M R,LANZILLOTTI C,MAZZIOTTA C,et al. The role of microRNAs in the osteogenic and chondrogenic differentiation of mesenchymal stem cells and bone pathologies[J]. Theranostics,2021,11(13):6573-6591. doi:10.7150/thno.55664.
[13] 張瑞欣,董语迪,肖建辉. lncRNA调控间充质干细胞向成骨细胞分化的研究进展[J].天津医药,2021,49(6):662-667. ZHANG R X,DONG Y D,XIAO J H. Research progress of lncRNA regulating the differentiation of mesenchymalstem cells into osteoblasts[J]. Tianjin Med J,2021,49(6):662-667. doi:10.11958/20203229.
[14] ZHANG F Y, ZHEN Y F, GUO Z X, et al. miR-143 is implicated in growth plate injury by targeting IHH in precartilaginous stem cells[J]. Int J Med Sci,2021,18(9):1999-2007. doi:10.7150/ijms.46474.
[15] RAVAL A,JOSHI J,SHAH F. Significance of metastamiR-10b in breast cancer therapeutics[J]. J Egypt Natl Canc Inst,2022,34(1):19. doi:10.1186/s43046-022-00120-9.
[16] LIU F,SHI Y,LIU Z,et al. The emerging role of miR-10 family in gastric cancer[J]. Cell Cycle,2021,20(15):1468-1476. doi:10.1080/15384101.2021.1949840.
[17] GE G,YANG D,TAN Y,et al. miR-10b-5p regulates C2C12 myoblasts proliferation and differentiation[J]. Biosci Biotechnol Biochem,2019,83(2):291-299. doi:10.1080/09168451.2018.
1533805.
[18] KOMORI T. Runx2,an inducer of osteoblast and chondrocyte differentiation[J]. Histochem Cell Biol,2018,149(4):313-323. doi:10.1007/s00418-018-1640-6.
[19] CHEN J,CHEN F,BIAN H,et al. Hypertrophic chondrocyte-specific Col10a1 controlling elements in Cre recombinase transgenic studies[J]. Am J Transl Res,2019,11(10):6672-6679.
[20] HU H H,CHEN D Q,WANG Y N,et al. New insights into TGF-β/Smad signaling in tissue fibrosis[J]. Chem Biol Interact,2018,292:76-83. doi:10.1016/j.cbi.2018.07.008.
[21] XIAO L,XU S,XU Y,et al. TGF-β/SMAD signaling inhibits intermittent cyclic mechanical tension-induced degeneration of endplate chondrocytes by regulating the miR-455-5p/RUNX2 axis[J]. J Cell Biochem,2018,119(12):10415-10425. doi:10.1002/jcb.27391.
