西南桦节间茎段植株再生体系的建立
2024-01-20郭俊杰王春胜尹海锋
王 欢,郭俊杰,王春胜,尹海锋,曾 杰
(1.中国林业科学研究院热带林业研究所,热带林业研究国家林草局重点实验室 广东 广州 510520;2.南京林业大学,江苏 南京 210037)
西南桦(Betula alnoidesBuch.-Ham.ex D.Don)是中国南方造林面积较大的乡土珍贵阔叶树种之一,其木材材质优良,是制作木地板、高档家具、乐器等的优良材料[1];其树皮可用于治疗感冒、产后疼痛以及消化系统和骨科疾病等[2],树皮提取物具有抗HIV-1 整合酶,并具有抗炎、降血脂、减肥之功效[3-5]。西南桦在生物多样性和地力维持、水源涵养以及碳素固定等方面亦发挥着重要作用,被广泛应用于生态公益林和珍贵用材林建设。随着分子生物技术的快速发展,利用基因工程进行遗传改良已成为林木育种的重要途径之一。遗传转化是开展植物遗传改良的必要技术,同时也是验证基因功能、解析性状形成机制的关键技术,而高效稳定的转化受体再生体系是遗传转化的必要前提。
中国西南桦的离体培养始于2000 年[6],主要有直接器官发生[7-9]、间接器官发生[10]以及体胚发生[6]等构建方式。目前能形成完整植株的主要是通过直接器官发生途径,特别是腋芽萌发途径,但该途径增殖率通常较低,且只适合于西南桦优良品种的规模扩繁,并不适用于遗传转化。间接器官发生是离体培养中通过外植体诱导愈伤组织再分化的形态建成过程[11],相比直接器官发生,其潜在繁殖系数更高,作为分子育种的平台也更具优势[12-13]。目前,西南桦通过间接器官发生方式的再生体系尚未见成功报道。
关于桦木科的间接器官发生,目前以白桦(Betula platyphyllaSuk)和欧洲白桦(Betula pendulaRoth)研究较多,所用外植体一般为种子[14-16]、叶片[17-19]、根段[20]、茎段[21]等。以种子为外植体研发的再生体系,具有分化率高且再生芽多的优势,但是,由于该体系需要“切种子”,对于细小粒的桦木种子而言技术要求极高,且每粒种子的遗传背景不同,严格地说,它并不利于开展转基因植株的分子试验。利用无性系组培苗的根、茎、叶作为外植体,可使受体材料保持相对一致。有研究发现,利用白桦茎段、叶柄和叶片均能诱导愈伤组织和不定芽分化,茎段的效果整体上优于叶片和叶柄[22];欧洲白桦根较难诱导愈伤组织形成,且其愈伤组织易老化,需要较长时间诱导才能分化出不定芽[20]。前期研究发现,西南桦茎段与白桦[22]一样较易诱导愈伤组织和不定芽分化,但因节间较短容易切到芽点,增加了操作难度,且材料利用率不高。因此,增加西南桦试管苗的节间长度对茎段再生的外植体取材非常重要。
本研究以西南桦无性系TC2 试管苗为材料,针对愈伤组织诱导、不定芽分化、生根等器官间接发生阶段开展试验,重点研究激素浓度、暗培养时间和茎段外植体培育条件等对不定芽诱导的影响,以期建立高效稳定、重复性良好的西南桦间接器官发生再生体系,为今后开展西南桦组织培养以及利用基因工程进行其遗传性状改良奠定基础。
1 材料与方法
1.1 试验材料及条件
以课题组前期培养的西南桦无性系TC2 试管苗为材料;如无特别注释,其培养条件为光照强度2 000 lx,光照时间16 h·d-1,温度25 ℃ ± 2 ℃。
1.2 研究方法
1.2.1 茎段再生阶段培养基的筛选 茎段再生阶段培养基的筛选试验中,选取继代培养30 d 的TC2 试管苗,剪取其带顶芽茎段,接种于培养基WPM + 0.15 mg·L-1NAA + 20 g·L-1蔗糖 + 5.8 g·L-1琼脂(pH5.8),培养30 d 后避开腋芽剪取4~7 mm 节间茎段用于愈伤组织诱导试验。
(1)愈伤组织诱导阶段 愈伤组织诱导研究包括基本培养基筛选、外源激素的浓度筛选以及暗培养时间筛选3 个单因素试验(表1),分别含4、9 和3 个处理(水平)。所有试验每个处理均3 次重复,每次10 个茎段。
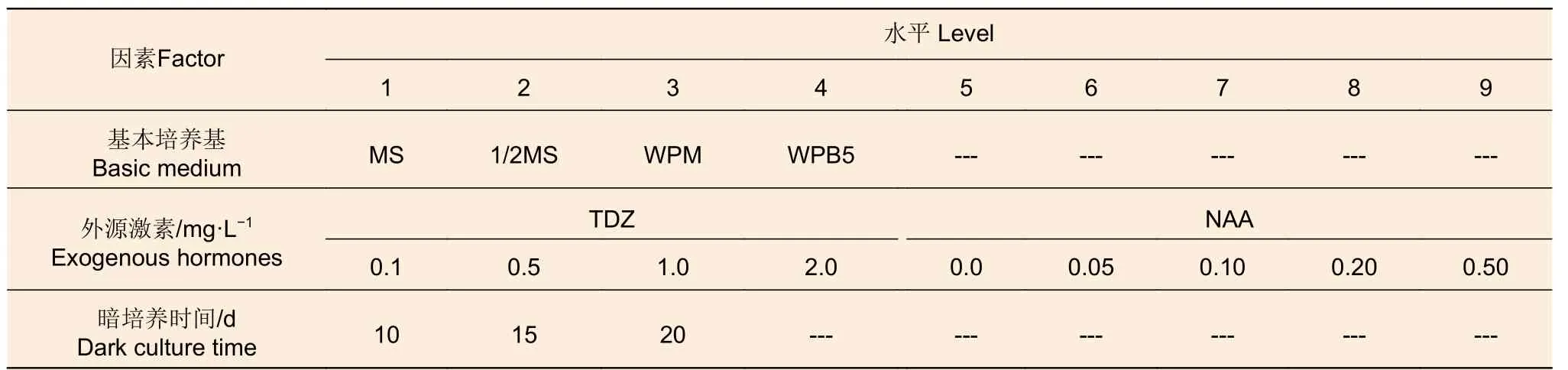
表1 愈伤组织诱导阶段的单因素试验Table 1 Single-factor experiments at callus induction stage
基本培养基试验中,各处理的激素添加均为1.0 mg·L-1TDZ 和0.1 mg·L-1NAA;外源激素浓度梯度试验以WPB5 为基本培养基,选用TDZ 和NAA 的不同浓度(表1)进行试验;暗培养时间优化试验按表1 中的3 个时长进行培养,所用培养基为WPB5 + 1 mg·L-1TDZ + 0.1 mg·L-1NAA +20 g·L-1蔗糖 + 5.8 g·L-1琼脂 (pH5.8)。统计愈伤诱导率后将表1 中所有试验处理的愈伤组织转接至培养基WPM + 1 mg·L-16-BA + 0.5 mg·L-1GA3+30 g·L-1蔗糖 + 5.8 g·L-1琼脂 (pH5.8)中进行后续分化培养,在正常光照条件下培养60 d 后统计发生分化的愈伤组织数与每个茎段分化的不定芽数(高度5 mm 以上),期间30 d 继代1 次,并计算分化率、净增殖系数和总增殖系数。
(2)愈伤组织分化不定芽阶段 以WPM 为基本培养基,采用6-BA(A)、GA3(B)和NAA(C)3 种激素(3 因素),每因素包括2 水平,构成完全随机试验(表2),分析各激素组合的不定芽诱导效果,以筛选分化培养基。选择在上述最佳愈伤组织诱导培养基上培养出的大小相近的愈伤组织,分别接种于8 种组合处理,每个处理3 次重复,每次重复接种10 块愈伤组织。每隔30 d 转接(继代)1 次,60 d 后观察统计不定芽诱导情况,包括发生分化的愈伤组织数、每个茎段分化的不定芽数、分化率、净增殖系数和总增殖系数。
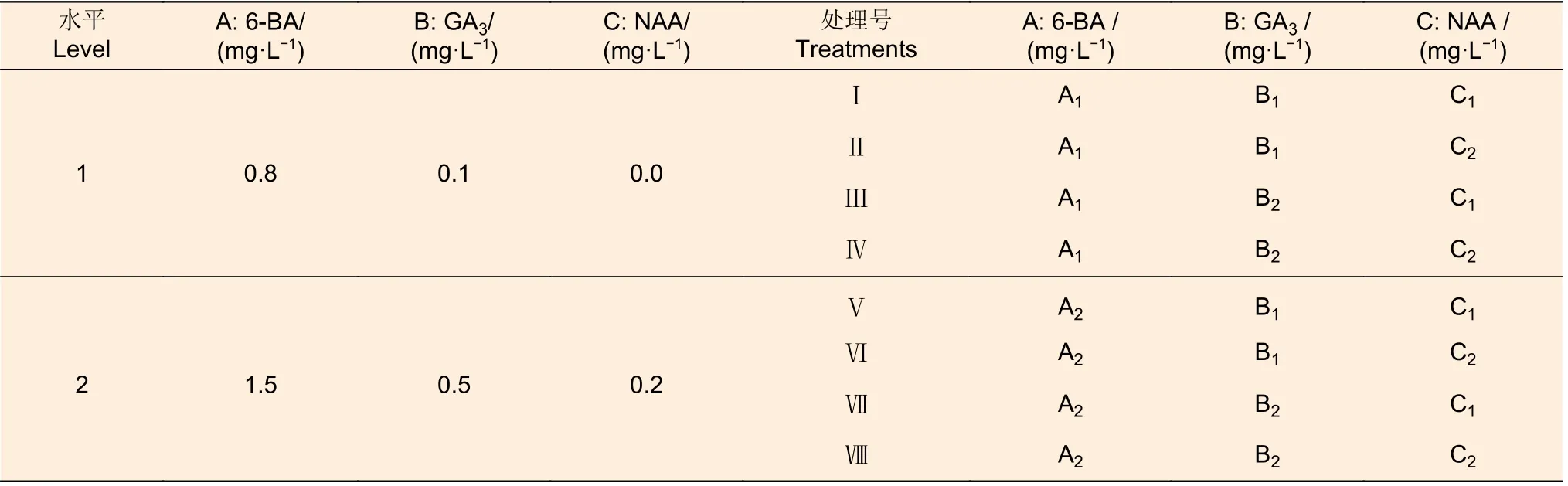
表2 愈伤组织分化阶段激素组合试验Table 2 Hormone combination experiment at callus differentiation stage
(3) 生根培养阶段 待分化的不定芽长至2~3 cm,剪取单芽接种至1/2MS 或WPM 培养基,附加NAA(0.1 和0.2 mg·L-1)或IBA (0.1 和0.2 mg·L-1)、20 g·L-1蔗糖和 5.8 g·L-1琼脂进行生根诱导,以筛选生根培养基。试验设计详见表3。每个处理3 次重复,每次重复12 株。培养过程中追踪统计各组合生根起始时间,于12 d 和30 d 后统计生根率和生根条数,并测定30 d 时根长。

表3 生根诱导试验设计Table 3 Experimental design for rooting induction
1.2.2 预培养条件的优化 因研究过程中发现外植体的状态影响各阶段的试验效果,于是针对预培养条件进行如下优化。选取继代培养30 d 的TC2 试管苗,剪取其带顶芽茎段,接种于培养基WPM + 0.15 mg·L-1NAA + 20 g·L-1蔗糖 + 5.8 g·L-1琼脂(pH5.8),于8 种条件下进行培育(表4)。每个处理3次重复,每次重复10 株,30 d 后统计株高和节间长度。分别取8 个处理的节间茎段,同时统计每株可取平均茎段数,然后接种于最佳愈伤组织诱导培养基(WPB5 + 1.0 mg·L-1TDZ + 0.2 mg·L-1NAA + 20 g·L-1蔗糖 +5.8 g·L-1琼脂 );暗培养15 d 后转至最佳愈伤组织分化的培养基(WPM + 0.8 mg·L-16-BA + 0.5 mg·L-1GA3+ 30 g·L-1蔗糖 + 5.8 g·L-1琼脂)。每个处理3次重复, 每次重复24 个茎段,75 d 后统计不定芽分化情况,统计指标同(2)。

表4 预培养条件筛选试验Table 4 Screening test on preculture conditions of internode
1.3 数据分析
统计各试验数据,计算以下指标:
Y=(n/N) × 100%,F=(m/M) × 100%,R=(q/Q) × 100%,Tt=Na/m,Tn=Na/M
Y为愈伤诱导率,n为愈伤组织诱导数,N为接种茎段数;F为愈伤分化率,m为分化愈伤组织数,M为接种愈伤组织数;R为生根率,q为生根株数,Q为接种数;Tt为总增殖系数,Tn为净增殖系数,Na为不定芽总数。使用Microsoft Excel 2010 软件进行数据整理,使用IBM SPSS Statistics13.0 软件进行方差分析和Duncan 多重比较。
2 结果与分析
2.1 愈伤组织诱导阶段各因素对后续不定芽诱导和增殖的影响
2.1.1 基本培养基 方差分析结果(表5)表明:除净增殖系数外,4 种基本培养基处理间产生的不定芽数(10 个接种茎段不定芽数量)及其分化率和总增殖系数均呈现显著差异(P<0.05),其中,WPB5 的所有指标均最高,WPM、1/2MS和MS 间不定芽数、分化率和总增殖系数依次显著地降低,净增殖系数差异不显著,说明采用WPB5作为基本培养基,可显著地促进西南桦茎段再生不定芽的形成,提高其增殖系数,因此,WPB5 是西南桦茎段再生不定芽培养的优选培养基。
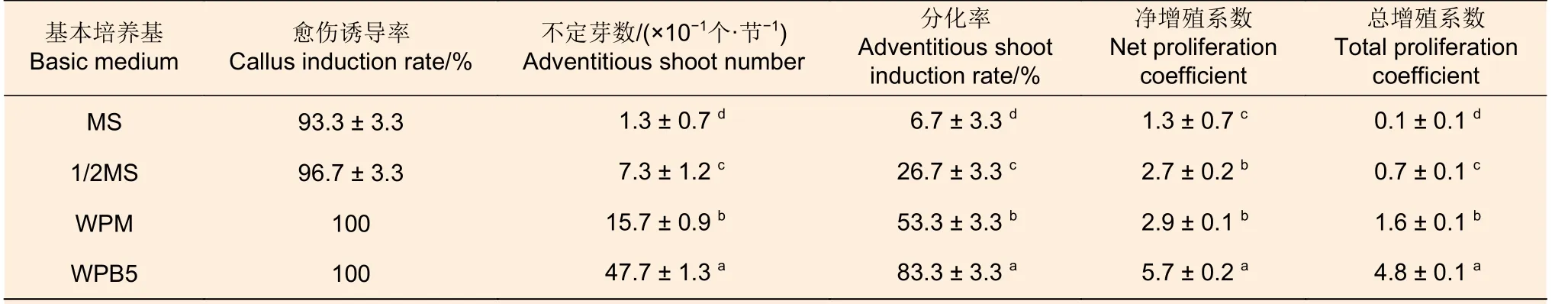
表5 愈伤组织诱导阶段基本培养基对后续西南桦不定芽分化的影响Table 5 Effect of basic medium at callus induction stage on subsequent adventitious bud differentiation of Betula alnoides
2.1.2 外源激素 由表6 可知:随着外源激素TDZ 或NAA 浓度的增加,不定芽数、分化率和总增殖系数均呈现先增加后下降的趋势,净增殖系数在两种激素中表现不同,随着TDZ 浓度增加呈递增趋势,而随NAA 浓度增加则呈现先增加后下降的趋势; TDZ 或NAA 不同浓度处理间4 个指标均存在显著差异。TDZ 的4 个浓度处理中,不定芽数、分化率和总增殖系数以1 mg·L-1处理为最高,分别为49、83.3%和4.9,显著地高于其它处理;净增殖系数最高的TDZ 浓度为2 mg·L-1,但与1 mg·L-1处理差异不显著。NAA 的5 个浓度处理中, 0.2 mg·L-1处理的不定芽数、分化率和总增殖系数均最高,分别为48.3、86.7%、4.8;净增殖系数最高的NAA 浓度为0.1 mg·L-1;除分化率在0.1 mg·L-1和0.2 mg·L-1处理间差异显著之外,其余指标在二者间均差异不显著。总的来看,TDZ 或NAA 为西南桦茎段愈伤组织诱导的关键外源激素,其适宜浓度分别为1 mg·L-1和0.2 mg·L-1。
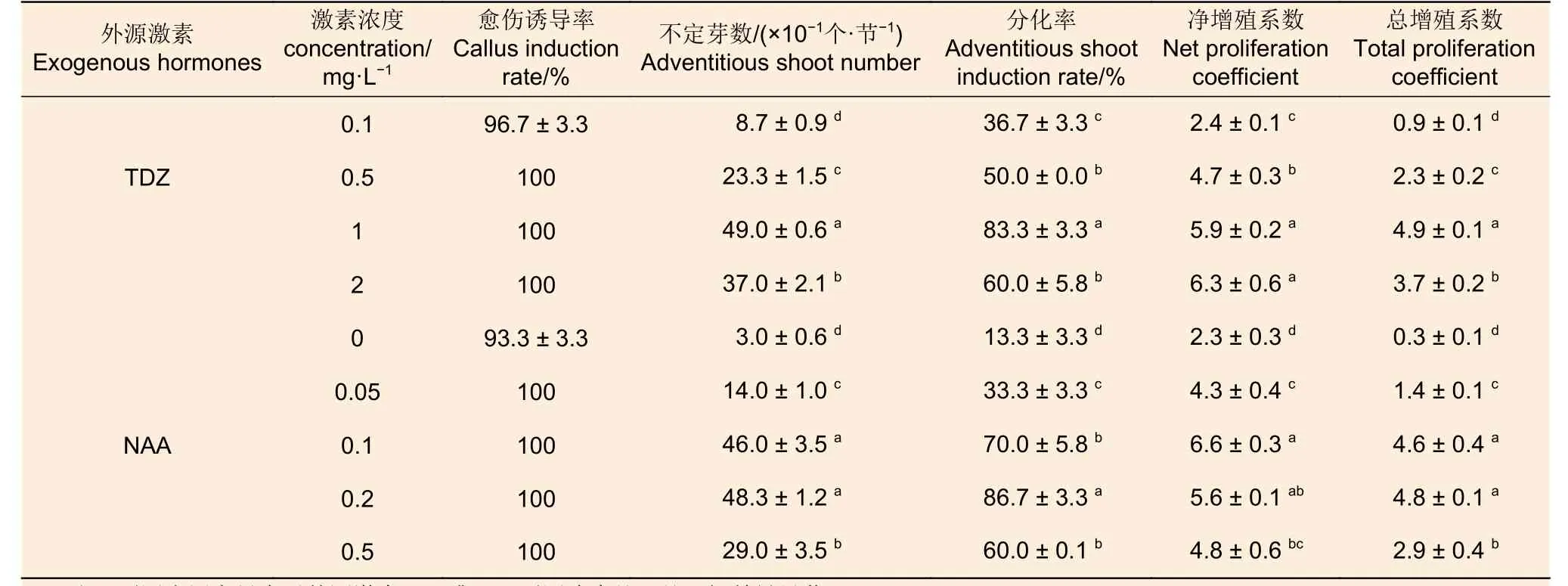
表6 愈伤组织诱导阶段外源激素及其浓度对后续西南桦不定芽分化的影响Table 6 Effects of exogenous hormones and their concentrations at callus induction stage on subsequent adventitious bud differentiation of Betula alnoides
2.1.3 暗培养时间 在暗培养时间筛选试验中,不定芽数、分化率、净增殖系数和总增殖系数随暗培养时间增加整体上呈现先增加后下降的趋势(表7)。暗培养15 d 和20 d 时4 个指标均显著高于10 d,且以15 d 暗培养处理效果最好;两者的分化率均在80.00%以上,净增殖系数和总增殖系数分别在5.5 和4.5 以上。由此可见,西南桦茎段愈伤组织诱导的适宜暗培养时间为15 d。
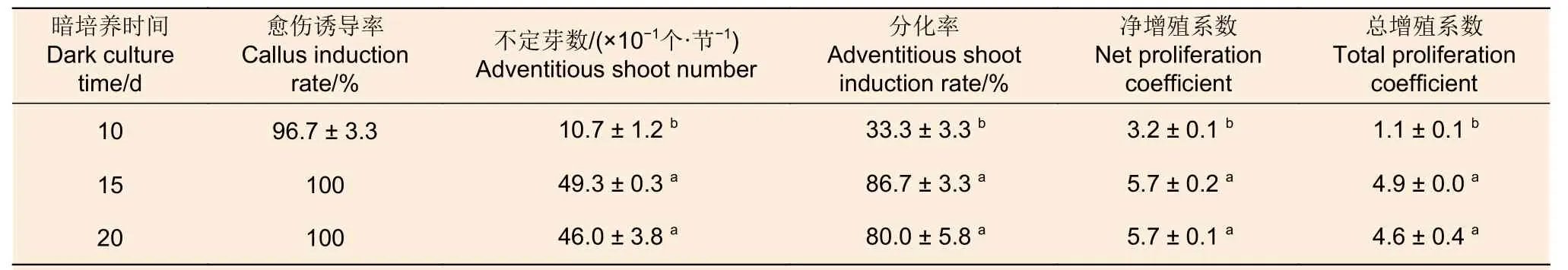
表7 愈伤组织诱导阶段暗培养时间对后续西南桦不定芽分化的影响Table 7 Effect of dark culture time at callus induction stage on subsequent adventitious buds differentiation of Betula alnoides
2.2 愈伤组织分化阶段不同激素组合对不定芽诱导和增殖的影响
GA3和NAA 两种因素各水平间不定芽数、不定芽分化率和总增殖系数存在显著差异,其净增殖系数无显著差异,6-BA 各水平间这些指标差异不显著;3 种因素间交互作用不显著(表8)。从表9可以看出:各激素组合处理的不定芽数、不定芽分化率、净增殖系数和总增殖系数分别为38.7~49.3、63.3%~86.7%、5.4 ~6.1 倍和3.9 ~4.9倍;组合处理间不定芽数、不定芽分化率、总增殖系数和净增殖系数均存在显著差异(P<0.05)。组合处理间比较,处理III 的不定芽数、不定芽分化率和总增殖系数最高,其净增殖系数也相对较高。对照表2 可知:0.8 mg·L-16-BA 和0.5 mg·L-1GA3的组合适宜应用于西南桦愈伤组织的不定芽分化和增殖。
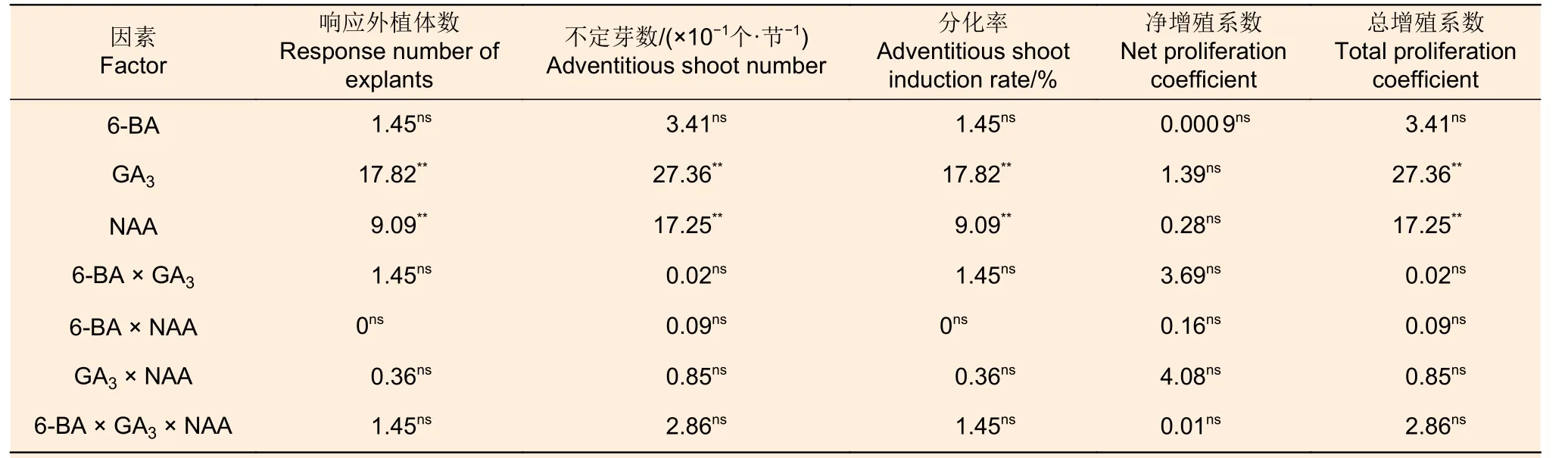
表8 西南桦不定芽分化指标的F 值和显著性分析Table 8 F-value and statistical significance of adventitious bud differentiation of Betula alnoides
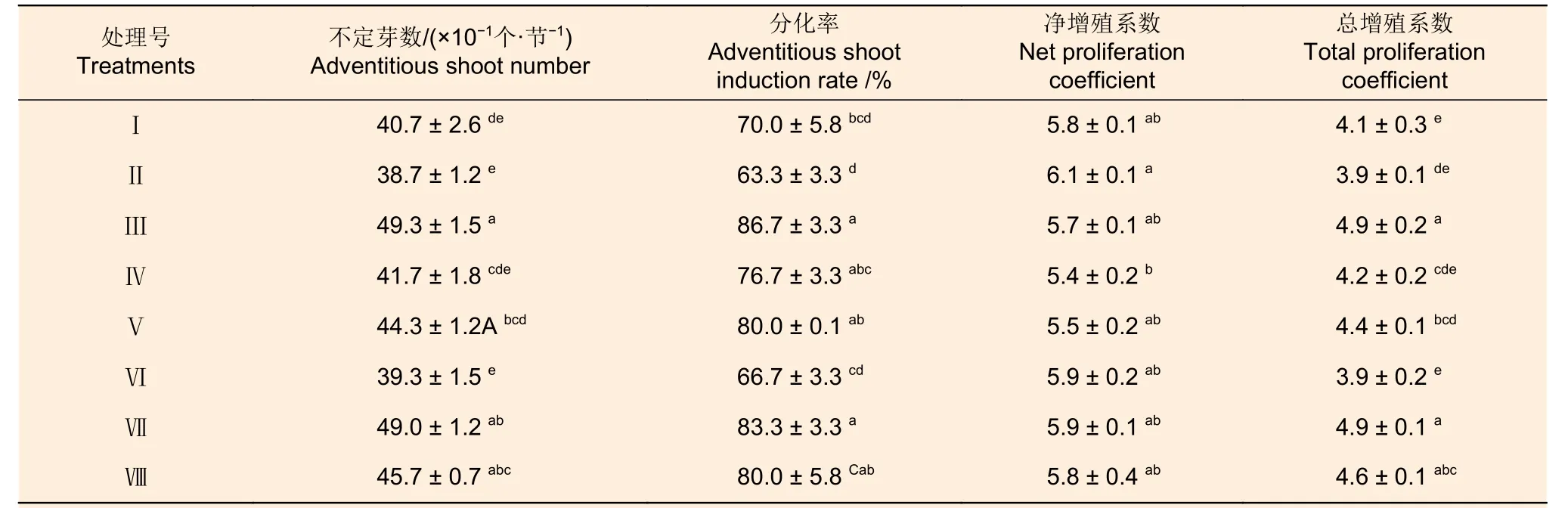
表9 不同激素组合对西南桦不定芽分化的影响Table 9 Effects of different hormone combinations on adventitious bud differentiation of Betula alnoides
2.3 不定芽生根诱导
培养12 d 和30 d 时,添加NAA 的处理(Ⅲ、Ⅳ、Ⅶ、Ⅷ)与添加IBA 的处理(Ⅰ、Ⅱ、Ⅴ、Ⅵ)间生根率和平均生根数均差异显著(表10)。处理Ⅱ和Ⅵ,基部出现大量致密的愈伤组织,30 d 时仍未见生根;处理Ⅰ和Ⅴ,基部出现较多愈伤组织,培养12 d 时未见生根,30 d 时虽有生根但生根率低,仅为22.2%~25.0%;其它4 种处理, 12 d 时生根率为33.3%~52.9%,平均生根数为2.4~6.7,30 d 时生根率均在90%以上,平均约10 条根,平均根长4.7 cm。对照表3 可知,无论采用1/2MS 还是WPM 作为基本培养基,添加0.1 mg·L-1或0.2 mg·L-1IBA 时不定芽不生根或生根率极低,而添加0.1 或者0.2 mg·L-1NAA的生根效果均较好。相对而言,适宜西南桦不定芽生根的基本培养基为WPM,外源激素及其浓度为0.1 mg·L-1NAA,在该条件下培养30 d 生根率可达100%。
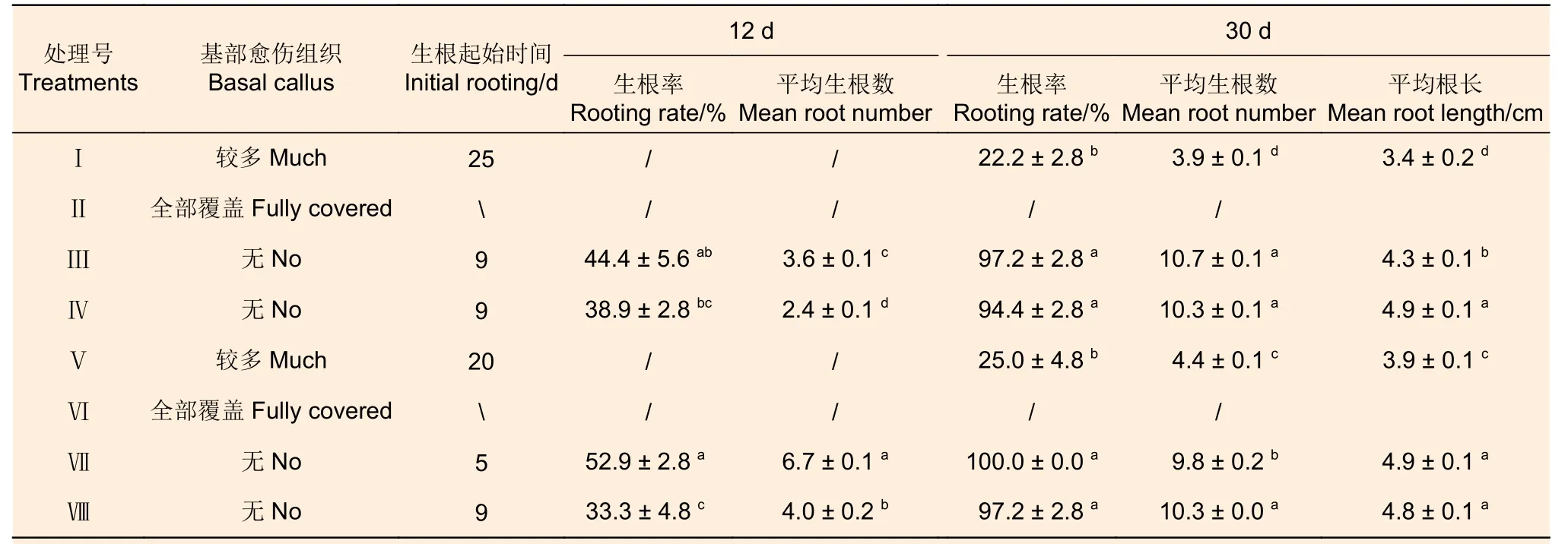
表10 不同培养基对西南桦不定芽生根的影响Table 10 Effects of different media on rooting of Betula alnoides adventitious buds
2.4 不同预培养条件对茎段待取苗生长和茎段再生的影响
方差分析结果(表11)显示:8 种预培养条件下,处理Ⅲ和Ⅵ的株高及每株可取平均茎段数显著高于其它处理,处理Ⅶ和Ⅷ的净增殖系数显著高于其它处理,8 个处理间分化率差异不显著。西南桦株高、节间长度以及每株可取茎段数均以处理Ⅵ表现最佳,其次为处理Ⅲ,两者的株高、节间长度以及每株可取茎段数分别在8 cm、3.5 cm 和12 以上,显著大于其它处理。分别采用各处理的茎段诱导愈伤组织并进一步分化培养,得到的分化率亦以处理Ⅵ为最高,但各处理间差异并不显著,均在86%以上;处理Ⅶ和Ⅷ的净增殖系数和总增殖系数均最高,净增殖系数在6.2 以上,总增殖系数约5.5,显著高于绝大多数其它处理。总而言之,处理Ⅶ的再生效率最高,平均每个预培养植株可产生56.8 个不定芽,整个再生过程见图1。因此,适宜西南桦茎段再生的预培养条件为处理Ⅶ,即弱光培养(1 000 lx)15 d 后转至暗培养7 d,再正常光照(2 000 lx)培养8 d。

图1 西南桦茎段再生各阶段的生长状态Fig.1 Growth status of each stage of regeneration from internodes of Betula alnoides
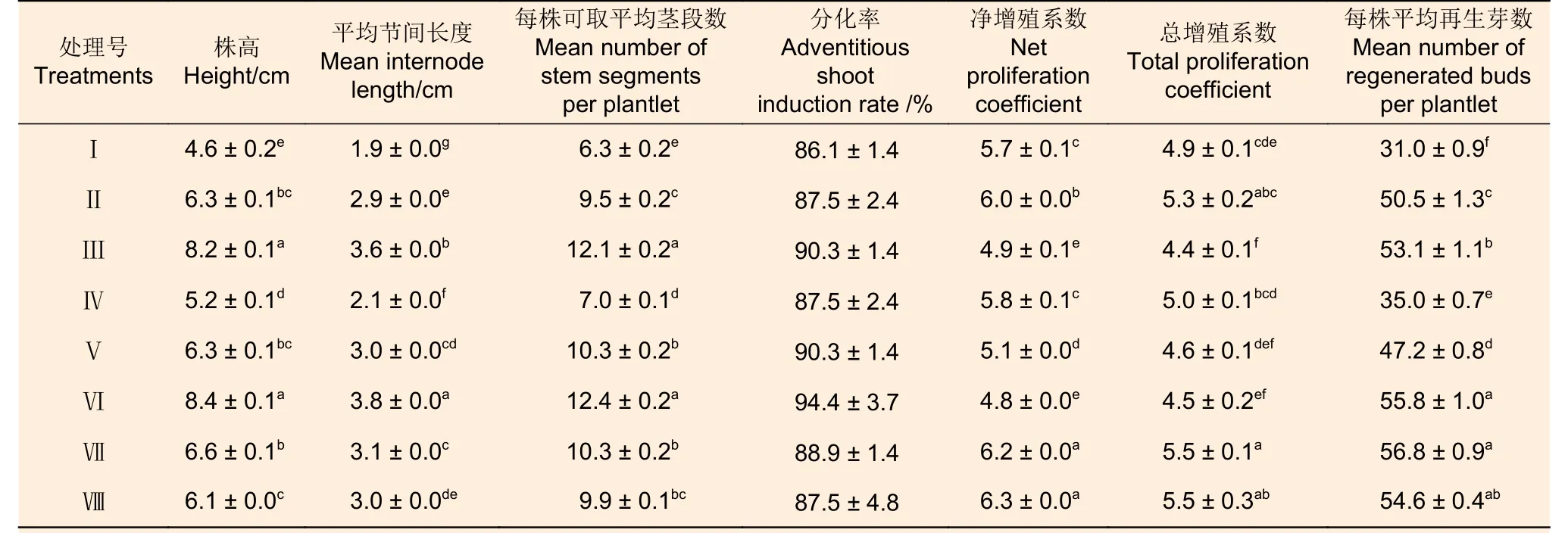
表11 不同预培养条件下西南桦植株生长表现和后续不定芽分化效果差异Table 11 Differences in growth performance and subsequent adventitious bud differentiation of Betula alnoides plantlets under various preculture conditions
3 讨论
外植体的选择是决定再生成效的第一步。幼态组织具有更高的形态发生能力,传统培养选取靠近形态学上端的材料更易成功[23-26],但每个植株可有效利用的材料较少,从而增加了取材时间和培养成本。对于节间茎段的取材而言,适当的黑暗培养可使试管苗生长加快、节间伸长、幼嫩程度增加[27],相对于传统培养仅能利用较短的顶端节间而言,提升其操作性的同时增加其茎段利用率。本研究通过设置8 种培养条件,筛选出再生效率和取材利用率均较高的组合,即弱光培养(1,000 lx)15 d 后,转至暗培养7 d,接着正常光照(2,000 lx))培养8 d。与常规培养(处理Ⅰ)相比,总增殖系数和净增殖系数分别提高8.8%和12.2%,每株平均再生芽数提高83.2%。究其原因,由于植物在黑暗中节间伸长加快,而随着光照时间的减少,光合作用变弱,光合产物也较少,相应地维管束和机械组织较不发达,更容易脱分化形成愈伤组织[28]。亦有研究表明,一段时间黑暗培养后的黄化苗,其生长素、可溶性糖、氨基酸和蛋白质等含量均比光培养苗高,即黄化培养后的幼茎生理代谢更活跃[29],更有助于脱分化形成胚性愈伤组织,从而提高其再生效率。
愈伤组织的诱导与其基本培养基、外源激素浓度、培养时间长短等因素密切关系。木本植物通常使用MS 或WPM 培养基作为基本培养基。本研究发现西南桦茎段在MS 培养基上不定芽诱导率极低,在WPM 培养基上也并不高,而在WPB5 培养基上表现较好。与WPM 培养基相比,WPB5 换用了B5 培养基的有机物,其有机物含量更高;相比MS 培养基,WPB5 替换了有机成分的同时还降低了氮和钾含量。由此可见,无机盐浓度低且有机物浓度高的基本培养基更适合西南桦茎段愈伤组织诱导。分析其原因,西南桦属于酚类物质较多的物种,培养过程中容易氧化形成醌类物质而产生褐变,从而影响外植体的脱分化和器官分化。而低盐培养基可以适当减轻此类褐变[30],从而诱导出愈伤组织。此外,在愈伤组织形成的起动期与分裂期,外植体细胞的合成代谢活动加强,需要迅速且大量地进行蛋白质和核酸的合成,适当添加有机物质可有效促进愈伤组织诱导[31]。
细胞分裂素和生长素是西南桦茎段器官发生途径愈伤组织诱导的必需因子,也是提高再生效率的重要因素。TDZ 具有生长素和细胞分裂素的双重功能,可使许多难再生的植物种实现高频分化[32-34]。如蒋淑磊等[34]对照山白杜鹃的茎段进行愈伤组织诱导研究发现,最佳TDZ 浓度为1.0 mg·L-1,愈伤组织诱导率可达96.3%,增殖系数为4.9。本研究通过设置0.1~2.0 mg·L-1的TDZ 浓度梯度开展试验,发现随着TDZ 浓度的增加,西南桦茎段的愈伤组织分化率和总增殖系数呈现先上升再降低的趋势,亦以1.0 mg·L-1浓度处理表现最好,过高则会导致愈伤组织逐渐变得疏松绵软,不利于不定芽分化。NAA 同样对愈伤组织的诱导至关重要[35],本研究发现,NAA 浓度为0.2 mg·L-1时,其愈伤组织分化出不定芽的概率与数量均比其它浓度高;浓度为0.5 mg·L-1时,其再生效果明显下降。关于暗培养时间,本研究结果显示,西南桦茎段愈伤组织诱导的最佳时间为15 d,培养20 d 效果仅次于15 d,但培养10 d 效果则明显降低。这可能与暗培养期间从培养基中吸收的TDZ 量有关,TDZ 具有快速诱导愈伤组织的效果,同时也具有剂量效应,因此外植体不宜长时间培养在该培养基上,但时间过短亦达不到相应效果。
分化率和增殖系数是决定不定芽分化效果的重要指标。本文对比研究了6-BA、GA3、NAA 3 种激素对西南桦不定芽分化的影响,发现GA3的影响最大,而且不同浓度对应的分化率和总增殖系数差异较大,浓度为0.1 和0.5 mg·L-1时,其分化率相差10%以上,总增殖系数相差0.5 以上。究其原因,GA3的使用有助于细胞伸长,加快不定芽的出芽速度,使得在统计时间段内出现较多的不定芽[36]。与之不同的是,低浓度6-BA 和NAA 更有利于不定芽分化。进一步分析发现,愈伤组织诱导阶段TDZ 的添加可能影响了分化阶段激素的添加。如前所述,TDZ 具有生长素和细胞分裂素的双重功能,既可以作为一种潜在的细胞分裂素物质,又可以诱导形成部分生长素类物质。愈伤组织诱导阶段高浓度TDZ 的添加使茎段愈伤组织积累了部分生长素和细胞分裂素类物质,从而较低浓度的添加即可满足愈伤组织分化的需求[37]。就6-BA 而言,虽然浓度越高,愈伤组织表面芽点越多,但过高的细胞分裂素/生长素比值容易导致这些芽点难以成芽,最终影响再生效率。本研究还发现,不添加NAA 时不定芽分化效果更好,亦可能与愈伤组织诱导阶段TDZ 和NAA 的添加积累了较多的生长素类物质有关,若在分化阶段继续添加生长素可能会导致激素配比不当,从而影响不定芽的分化。因此,综合考虑愈伤组织诱导和不定芽分化两阶段的激素使用对再生体系的建立至关重要。
4 结论
综上所述,本研究围绕西南桦节间茎段再生各阶段展开试验,建立了完整的西南桦间接器官发生途径再生体系。具体而言,增殖苗经弱光培养(1 000 lx) 15 d 后转至暗培养7 d,再正常光照(2 000 lx) 培养8 d 可获得适度黄化的植株,其平均株高和节间长度分别为6.6 cm 和3.1 cm;以其节间茎段为外植体,接种于添加1.0 mg·L-1TDZ和0.2 mg·L-1NAA 的WPB5 培养基上,暗培养15 d 后转接至含0.8 mg·L-16-BA 和 0.5 mg·L-1GA3的WPM 培养基上,正常光照培养2 个月,期间继代1 次,则可实现愈伤组织分化率和净增殖系数达88.9%和6.2 以上,平均每个植株可产生56.8个不定芽;不定芽接种于添加0.1 mg·L-1NAA 的WPM 培养基上,培养30 d 后生根率可达100%。本研究为西南桦的遗传改良提供了理论依据和技术支持,亦为其组培快繁开辟了一条行之有效的新途径。
