8 个油橄榄品种扦插苗苗期耐寒性评价
2024-01-20吴文俊戚建莉金高明姚玉芳马成艳姜成英
吴文俊,戚建莉,金高明,张 荣,姚玉芳,马成艳,姜成英
(甘肃省林业科学研究院 国家林业和草原局油橄榄工程技术研究中心 甘肃省油橄榄工程技术研究中心,甘肃 兰州 730020)
油橄榄(Olea europaeaL.)是木樨榄属(Olea)常绿乔木,世界四大木本油料作物之一,野生油橄榄起源于小亚细亚,主产区在地中海区域[1],但南、北半球的40 多个亚热带国家都有引种栽培,植物油产量居世界第六位[2]。油橄榄与其他亚热带常绿经济林树种一样,冷胁迫一方面影响其地理分布,另一方面造成产量的巨大损失[3],希腊和意大利就曾在2004 年和2008 年遭遇寒冷天气,导致油橄榄园受灾,鲜果减产[4]。我国西秦岭南坡的油橄榄主产区,位于北亚热带北缘,低温是限制该区域油橄榄地理分布和鲜果减产的重要因素[5],而引种和筛选耐低温品种能一定程度上解决低温对油橄榄栽培带来的不利影响[6]。
植物可以通过在低温条件下激活抗氧化酶系统抵御超氧自由基的毒害[7];通过调节组织中可溶性蛋白、非结构性碳水化合物以及游离脯氨酸含量来提高细胞液渗透势,提高对寒冷的忍耐力[8];低温下植物净光合速率、气孔导度、蒸腾速率和胞间CO2浓度等指标的变化能够反映植物光合器官受损程度,也与耐寒能力间有一定关系[9]。因此耐寒生理指标、植物光系统在低温下的破坏程度都可以作为耐寒性评价的方法,但由于各生理指标对低温敏感性差、实验数据可重复性低等因素,使植物耐寒能力定量化评价变得尤为复杂。在油橄榄适应性育种工作中,有许多科研人员致力于研发可靠的油橄榄耐寒性评价技术,用于筛选耐低温品种。地中海油橄榄主产区,M.Gómez 等人通过对10 个油橄榄品种越冬(-10.5 ~ -6.5 ℃)表现进行观测,发现油橄榄嫩枝抗冻性与发芽能力呈线性相关[10],品种间抗冻性存在显著差异,品种‘Cornicabra’和‘Arbequina’低温后嫩枝损伤率低,再生能力强,而‘Empeltre’、‘Frantoio’和‘Hojiblanca’耐寒性差。Cansev 等人研究了9 个油橄榄品种抗氧化酶的变化规律与耐寒性的关系,发现品种‘Domat’、‘Lecquest’、‘Ascolona’、‘Hojoblanca’和‘Gemlik’的组织通过增加过氧化氢酶(CAT)、抗坏血酸过氧化物酶(APX)和还原型辅酶II(NADPH)等酶的活性来激活抗氧化系统和脱氢酶的表达,增强了低温下细胞膜结构的稳定性,具有较强的耐寒能力[11]。在我国油橄榄作为引进树种,许多研究人员期望通过耐寒性评价来获得适合北亚热带高纬度、高海拔区域发展的品种,来扩大适生区。80 年代,在油橄榄适应性研究过程中,就发现叶片节间短、叶片厚而浓绿、叶缘反卷等叶片形态与耐寒性呈正相关[12]。近几年,令凡等人也通过低温胁迫试验,对6 个油橄榄品种1 年生扦插苗耐寒能力的大小进行了评价,由强到弱依次为:‘Picholine’>‘Pendolino’>‘Leccino’>‘Ascolana’>‘Ez-8’>‘Frantoio’,但是采用油橄榄离体植物叶片进行低温胁迫试验所评价耐寒能力大小与活体样本有所偏差[5]。此外,低温胁迫对光系统破坏与植物耐寒性的关系在甘蔗、油菜等植物中已有研究报道[13-14],低温不耐寒品种的光系统损伤较大,叶绿素荧光参数、叶绿素含量均明显较耐寒品种低,但利用叶绿素荧光参数和气体交换参数结合评价油橄榄耐寒性的研究方法比较少见。试验以活体植株进行冷胁迫,以期揭示不同品种油橄榄幼苗膜系统、渗透调节物质、抗氧化酶系统、光系统对低温胁迫的响应,评价不同油橄榄品种幼苗耐寒性,为筛选耐寒品种提供方法和依据。
1 材料与方法
1.1 材料
植物材料为2 年生油橄榄无性系扦插苗,由油橄榄国家林木种质资源库提供,地点位于甘肃省陇南市武都区两水镇段河坝村,地理坐标(104°47′59″~104°49′02″ E,33°23′45″~33°24′57″ N),属于北亚热带半湿润气候,年平均气温14.9 ℃,绝对最高温40 ℃,极端低温-8.6 ℃,年日照1 911.7 h,无霜期150~270 d,年降水量474~900 mm,年平均湿度61%~71%,供试品种见表1(木质化程度基本一致)。

表1 供试油橄榄材料基本情况Table 1 Information of the test materials of olive seedlings
1.2 试验处理
2021 年3 月,每个供试品种挑选健壮且长势基本一致的30 株2 年生无性系扦插苗,盆栽于3 L容器中(肥水条件一致),至11 月中旬大田温度降至20 ℃左右时,参照Ben-Ari[15]、Davide Guerra[16]等人冷胁迫方法,每个品种挑选21 株置于人工气候室中,在22 ℃/18 ℃条件下适应14 d(光照时间为8:00—18:00,光照强度800 μmol·m-2·s-1、湿度50%~60%,温度控制误差为 ± 0.5 ℃),3 株作为对照(CK),其余18 株保持与之前相同的光强和光周期,以1 ℃·h-1降温速度进行低温处理,分别在达到16、12、8、4、0、-4 ℃时维持24 h,在各温度胁迫结束前3 h 进行采样和测试。
1.3 方法
胁迫结束前3 h 每个品种随机选取3 株,每株选取枝条顶端至基部方向第2~4 对叶片中叶形、大小较为一致的3 片叶,进行光合和荧光参数测定。随机摘取枝条顶端至基部方向第2~4 对叶片中5~6 片,根据李玲等[13]人的方法进行叶绿素含量(Chl)、可溶性糖(SS)、可溶性蛋白(SP)、脯氨酸(Pro)、丙二醛(MDA)、超氧化物歧化酶(SOD)、过氧化物酶(POD)、过氧化氢酶(CAT)等指标参数测定。
光合参数测定利用LI-6 400 便携式光合作用分析系统;叶绿素荧光参数测定采用CF-imager 叶绿素荧光图像分析仪进行离体测定;电导率用DDS-11A 型电导率仪测定。
1.4 数据处理
采用Excel 2010 整理数据和SPSS 23.0 进行数据处理,用Duncan 多重比较检验不同处理间的差异显著性(ɑ=0.05),用origin2019 作图。
2 结果与分析
2.1 相对电导率及半致死温度(LT50)
如图1a 所示,温度从22 ℃降到0 ℃时,所有供试品种相对电导率逐渐增高,说明细胞膜透性缓慢增加,电解质在缓慢渗出。当温度降低至-4 ℃时,‘柯基’、‘佛奥’、‘阿尔波萨纳’和‘鄂植8 号’叶片相对电导率分别升高至76.63%、72.50%、73.27%和73.00%,说明其叶片细胞膜受损严重,而‘豆果’、‘皮瓜尔’、‘科拉蒂’和‘莱星’相对电导率均小于70%,细胞膜受到伤害较小。通过Logistic 方程对各品种不同处理温度和叶片相对电导率进行拟合,确定了LT50。发现8 个油橄榄品种叶片相对电导率与半致死温度间呈较强正相关,供试品种的LT50介于-16.359~-9.715 ℃之间表2,耐寒性排序如下:‘豆果’>‘皮瓜尔’>‘科拉蒂’>‘莱星’>‘佛奥’>‘鄂植8 号’>‘阿尔波萨纳’>‘柯基’。
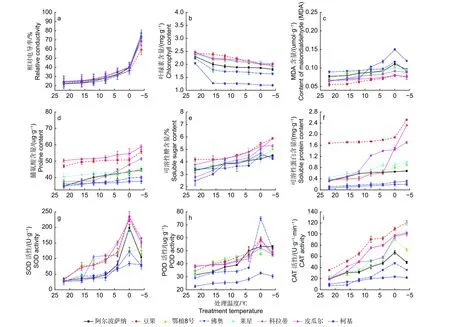
图1 低温胁迫过程中8 个油橄榄品种生理指标变化Fig.1 The change characteristics of physiological indices in eight olive seedlings under low-temperature stress
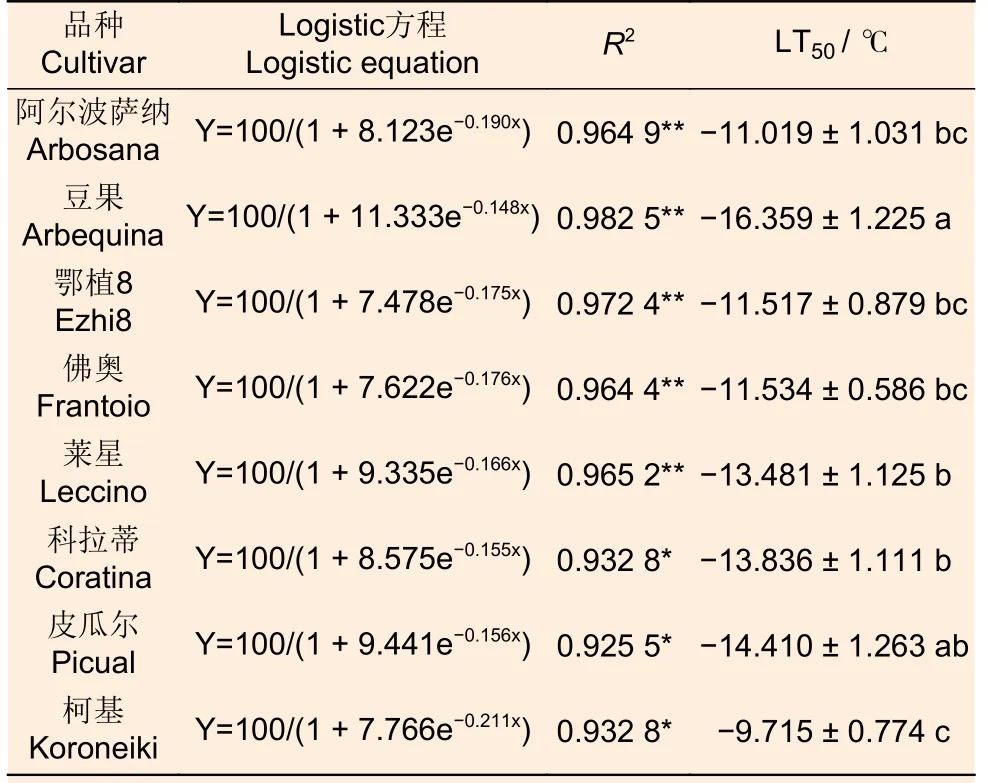
表2 参试品种半致死温度Table 2 Semi-lethal temperature of tested varieties
2.2 低温胁迫下各品种油橄榄生理响应及耐寒性排序
2.2.1 生理指标响应
2.2.1.1 叶绿素含量的变化 8 个油橄榄品种叶片叶绿素含量随温度的下降而减少。从22 ℃降至-4 ℃,‘豆果’和‘皮瓜尔’的叶片叶绿素含量降幅较小,分别为18.31%和18.21%,而‘佛奥’和‘柯基’降幅最大,分别为26.88%和41.37%(图1b)。
2.2.1.2 丙二醛含量的变化 ‘豆果’、‘皮瓜尔’、‘科拉蒂’和‘莱星’叶片丙二醛(MDA)含量总体低于其他品种,在低温胁迫加剧的过程中呈先升高后降低的趋势,都在0 ℃达到高峰。0 ℃降低至-4 ℃的过程中,‘柯基’和‘佛奥’降幅度最大,为20.60%和22.61%(图1c)。
2.2.1.3 低温胁迫下脯氨酸含量的变化 供试品种叶片脯氨酸含量随着低温胁迫加剧总体呈升高的趋势,在-4 ℃均达到峰值,与0 ℃相比,‘皮瓜尔’增幅较大,为7.58%,而‘柯基’和‘佛奥’增幅较小,分别为0.46%和1.05%(图1d)。
2.2.1.4 低温胁迫下可溶性糖含量的变化 叶片可溶性糖含量随着胁迫加剧总体呈升高趋势,在-4 ℃达到峰值,‘豆果’可溶性糖含量显著高于其他品种。在0~-4 ℃,‘柯基’和‘佛奥’的可溶性糖含量均有所下降,‘柯基’降幅明显(图1e)。
2.2.1.5 低温胁迫下可溶性蛋白含量的变化 ‘豆果’可溶性蛋白含量显著高于其他品种,叶片可溶性蛋白含量随温度降低而升高。-4 ℃胁迫下,‘皮瓜尔’的增幅最大,为54.14%,‘豆果’次之,而‘柯基’的增幅仅为1.67%,说明其受低温影响较大(图1f)。
2.2.1.6 SOD 活性的变化 随着处理温度的降低,各油橄榄品种叶片的SOD 活性总体呈先升高后降低的趋势,同一品种在不同低温胁迫下均存在显著差异(P<0.05)。在0 ℃均达到峰值,其中‘豆果’的SOD 活性最高,为235.19 U·g-1,‘科拉蒂’次之。与室温CK相比,-4 ℃时,‘莱星’和‘科拉蒂’的增幅最大,‘豆果’次之,而‘柯基’和‘佛奥’增幅最小,分别为197.46%和193.70%(图1g)。
2.2.1.7 POD 活性的变化 8 个油橄榄品种叶片的POD 活性随着处理温度的降低总体呈先升高后降低的趋势。除‘鄂植8 号’在 -4 ℃达到峰值,而其余品种均在0 ℃达到峰值,0~-4 ℃活性降低,其中‘豆果’的POD 活性总体高于其他品种,而‘柯基’最小(图1h)。
2.2.1.8 CAT 含量的变化 随着处理温度的降低,‘豆果’、‘皮瓜尔’、‘科拉蒂’和‘莱星’的CAT 活性均呈上升趋势,在-4 ℃达到峰值,而‘鄂植8 号’、‘阿尔波萨纳’、‘佛奥’和‘柯基’均呈先升高后降低的趋势,在0 ℃达到峰值(图1i)。
2.2.2 基于生理响应数据的耐寒性排序 以相对电导率、叶绿素含量、可溶性糖含量和可溶性蛋白质含量等9 个参数,计算各参数的平均隶属函数值,并进行耐寒能力综合评价,‘豆果’、‘皮瓜尔’、‘科拉蒂’和‘莱星’的平均隶属函数值差异不显著,但与其它品种之间存在显著差异(P<0.05)。8 个品种油橄榄耐寒性由强到弱见表3,依次为:‘豆果’>‘皮瓜尔’>‘科拉蒂’>‘莱星’>‘鄂植8 号’>‘阿尔波萨纳’>‘佛奥’>‘柯基’。
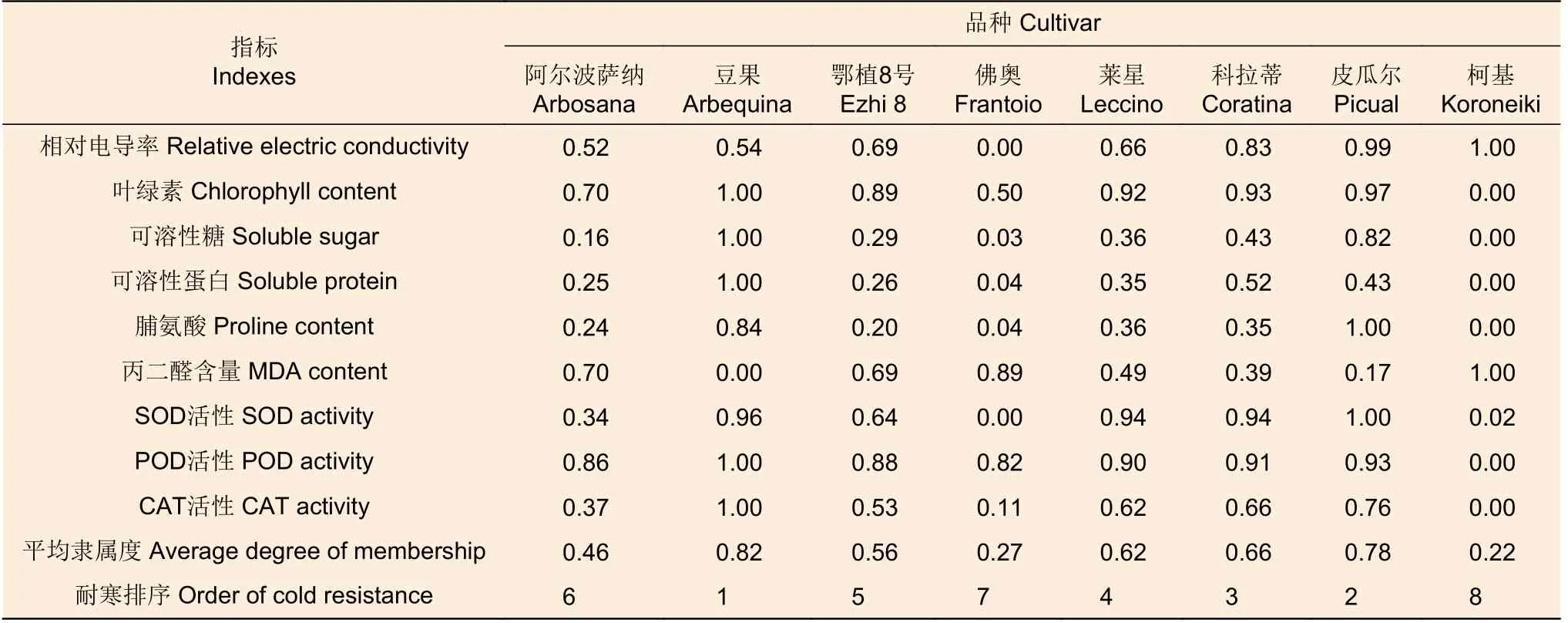
表3 低温胁迫下8 种油橄榄幼苗测定指标的隶属函数值Table 3 Membership function values of the eight olive seedlings under low-temperature stress
2.3 低温胁迫对油橄榄光合荧光特性的影响
2.3.1 低温胁迫对油橄榄幼苗光合气体交换参数的影响 参试油橄榄品种在室温下(22 ℃),各光合参数就有所差异(图2)。随着低温胁迫加剧,参试油橄榄品种叶片净光合速率(Pn)整体呈下降趋势,-4 ℃与对照(CK,22 ℃)相比,‘皮瓜尔’降幅最小为84.79%,其他品种降幅都在90%以上。低温胁迫过程中,气孔导度(Gs)在4 ℃前整体呈下降趋势,4 ℃与CK 相比,‘皮瓜尔’和‘科拉蒂’的降幅最小,分别较CK 下降了49.57%和78.37%;‘阿尔波萨纳’降幅最大,达到96.02%。胞间CO2浓度(Ci)在8 个品种中呈现出不同的变化规律,-4 ℃时‘阿尔波萨纳’和‘柯基’较CK 降低了5.36%和12.33%,其他品种都有所上升,其中‘佛奥’增幅最大为119.35%。随着低温胁迫的加剧,蒸腾速率(Tr)在-4 ℃出现明显下降,相较于CK 各品种降幅基本一致,介于93.76%~96.80%。低温加剧过程中气孔限制值(Ls) 呈现先升后降再升的趋势,增加时峰值集中出现在16 ℃和12 ℃, ‘皮瓜尔’和‘阿尔波萨纳’增幅最大,分别为153.49%和137.04%(12 ℃)。水分利用效率( WUE )呈现先升后降再升的趋势,再次上升的拐点温度出现在4℃,相比对照‘阿尔波萨纳’降幅最大为56.04%。

图2 低温胁迫过程中8 种油橄榄幼苗气体交换参数的变化特征Fig.2 The change characteristics of gas-exchange parameters in eight olive seedlings under low-temperature stress
2.3.2 低温胁迫对油橄榄叶绿素荧光参数的影响
由图3 可见,在低温胁迫下,8 个供试油橄榄品种的最大光化学效率(Fv/Fm)整体呈下降趋势。-4 ℃时,‘鄂植8 号’较CK 降幅较小为33.26%,而‘柯基’较CK 降幅最大为50.55%。从16 ℃到4 ℃胁迫过程中,‘佛奥’和‘莱星’变化趋势一致且较为缓和。
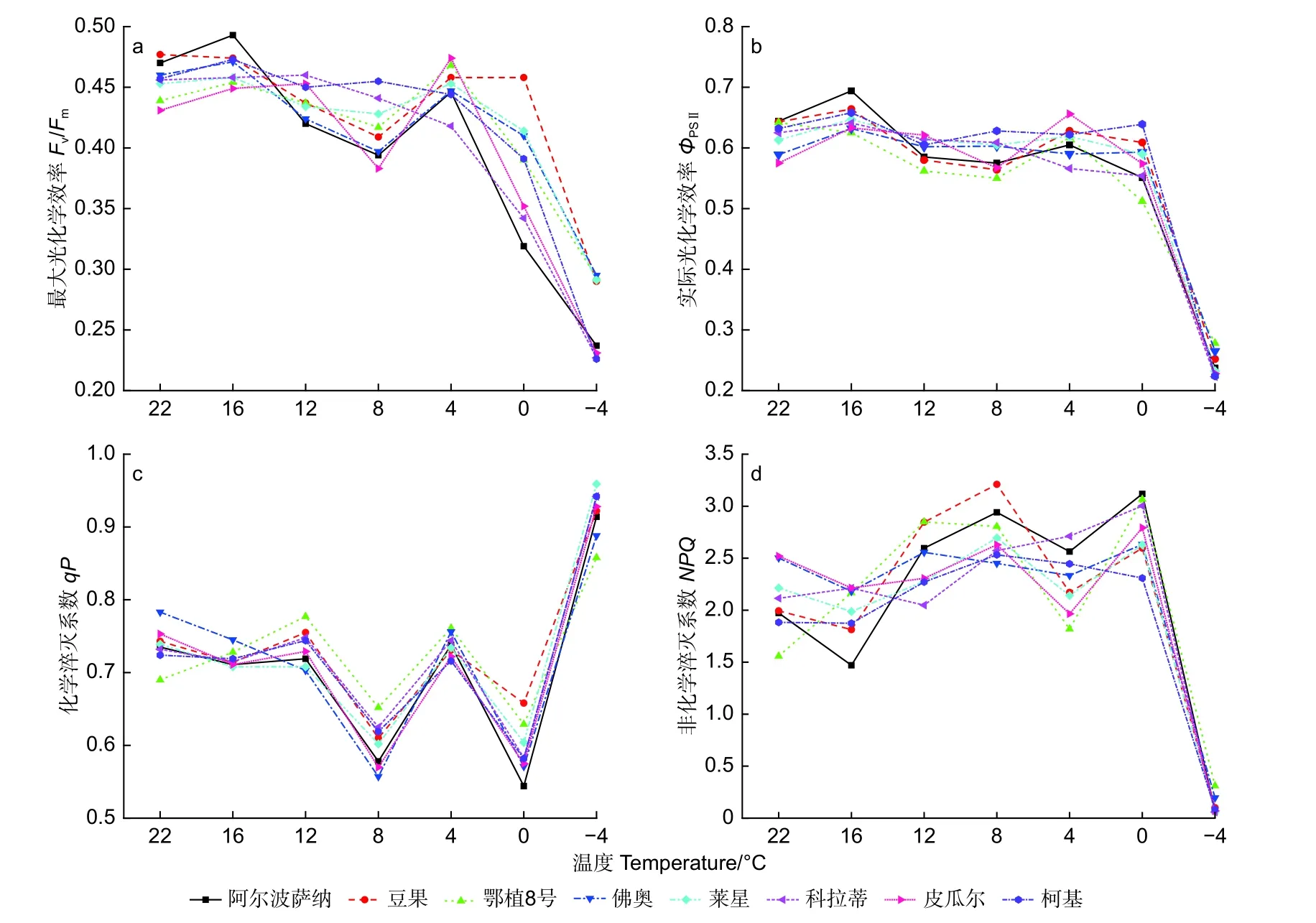
图3 低温胁迫过程中8 种油橄榄幼苗叶绿素荧光参数的变化特征Fig.3 The change characteristics of chlorophyll fluorescence parameters in eight olive seedlings under lowtemperature stress
经低温胁迫,实际光化学效率(ΦPSⅡ)在0 ℃前变化趋势较为缓和,0 ℃至-4 ℃出现明显下降,‘阿尔波萨纳’、‘豆果’、‘鄂植8 号’、‘佛奥’、‘莱星’和‘科拉蒂’、‘皮瓜尔’和‘柯基’降幅分别为63.20%、60.81%、56.70%、55.01%、62.15%、64.32%、60.35%、64.56%。
各参试品种的化学淬灭系数(qP)整体呈“W 形”趋势,最低值出现在8 ℃和0 ℃,4 ℃时有上升趋势,在-4 ℃时骤升出现峰值。‘阿尔波萨纳’、‘豆果’、‘鄂植8 号’、‘佛奥’、‘莱星’和‘科拉蒂’、‘皮瓜尔’和‘柯基’增幅分别为24.35%、24.09%、24.35%、13.41%、29.77%、28.69%、23.24%、30.11%。
随温度胁迫的加剧,各供试油橄榄品种的非化学淬灭系数(NPQ)值在22 ℃至0 ℃变幅度不大,介于1.558~3.118,0 ℃至-4 ℃出现急剧下降,降低至0.05 左右。
2.3.3 基于光合及叶绿素荧光参数的耐寒性排序
采用隶属函数法对8 个供试品种光合和叶绿素荧光参数的10 个指标进行耐寒性综合评价。由表4可知,8 个油橄榄品种的平均隶属度在0.41~0.71 之间,从隶属函数综合评价值可得出耐寒性依次为‘科拉蒂’>‘豆果’>‘柯基’>‘鄂植8 号’=‘佛奥’=‘莱星’>‘皮瓜尔’>‘阿尔波萨纳’。‘科拉蒂’的综合评价值高达0.71,耐寒性最强;‘阿尔波萨纳’的综合评价值仅为0.41,耐寒性最差。
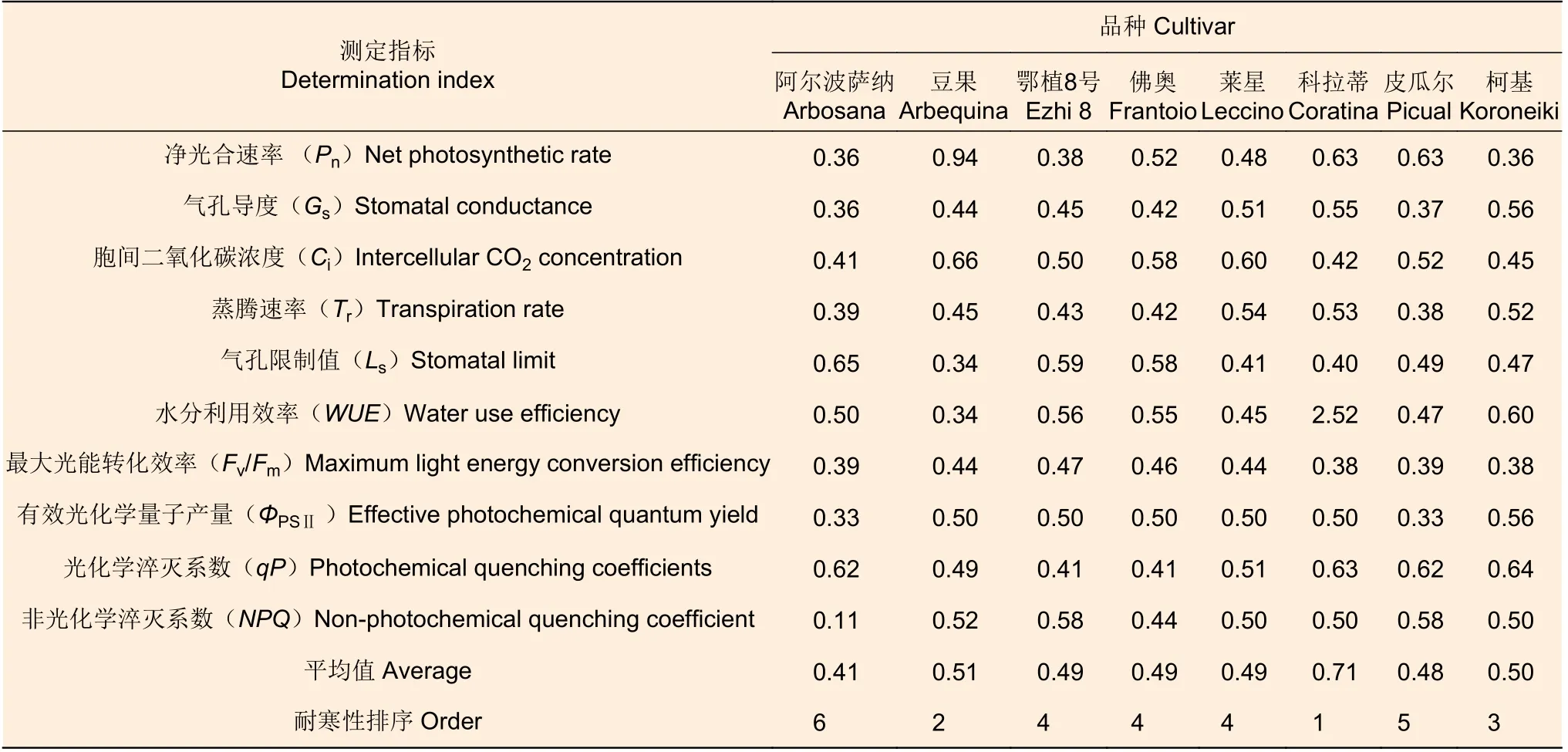
表4 低温胁迫下8 种油橄榄幼苗测定指标的隶属函数值(光合参数)Table 4 Membership function values of the eight olive seedlings under low-temperature stress (photosynthetic parameters)
3 讨论
3.1 膜系统对低温的响应
冷胁迫时,植物细胞中细胞膜最先受到伤害,导致细胞膜电解质外渗,且随着伤害程度的增加,离子渗出率升高,利用相对电导率结合Logistic 曲线方程获得植物的半致死温度是一种有效且简便的耐寒性评价方法,该方法已在金光杏梅、花椒、葡萄等诸多植物的耐寒性评价中得到应用[17-19]。本试验中,随着温度从22 ℃降到0 ℃时,油橄榄叶片电解质在缓慢渗出(相对电导率35% 左右),当温度从0 ℃降低到-4 ℃时,‘柯基’、‘佛奥’、‘阿尔波萨纳’和‘鄂植8’号叶片相对电导率分别升高至76.63%、72.50%、73.27% 和73.00%,说明这4 个品种细胞膜受到伤害较大,而‘豆果’、‘皮瓜尔’、‘科拉蒂’和‘莱星’叶片相对电导率均在70%以下。通过Logistic 方程对不同品种的处理温度和叶片相对电导率进行拟合,试验中8 个油橄榄品种,半致死温度(LT50)介于-16.359~-9.715 之间,‘豆果’耐寒性最好,‘柯基’最差,供试品种中‘莱星’、‘鄂植8 号’、‘佛奥’的耐寒性排序与令凡等人的研究结果一致[20]。
MDA 是植物脂膜过氧化的重要产物之一,积累会使脂膜在逆境中的损伤加剧[21],通过比较MDA 含量及其变化程度,可以反映出8 个油橄榄品种在低温中的受损程度。本研究中,8 ℃之后‘柯基’MDA 含量上升幅度显著高于其它7 个品种,说明‘柯基’细胞膜氧化程度更大,符合半致死温度以及耐寒性综合评价结果。
3.2 渗透调节物质和抗氧化酶活性对低温胁迫的响应
Chl、SS、SP、Pro、POD、SOD、CAT 等指标虽然都与植物耐寒性关系密切,但是由于植物是一个有机整体,遭受低温胁迫而引起生理生化指标变化也是错综复杂的,因此用单一指标来评价植物耐寒性太过片面。当然,想要准确评价某一种植物耐寒性所应用的指标体系也有所不同,例如细胞膜系中不饱和脂肪酸的变化就不能用于评价苹果(Malus pumilaMill.) 、 菜 蓟 (Cynara scolymus)、西蕃莲属(PassifloraL.)植物的耐寒性;脯氨酸含量的累积无法用来评价柑橘属植物的抗冷性。冷胁迫条件下,植物体内会积累大量超氧自由基,从而破坏植物膜系统,抗氧化酶系统可以清除超氧自由基,从而避免植物膜系统在冷害下遭到破坏[22]。本研究中,8 个油橄榄品种抗氧化酶活性随着温度的降低呈先上升后下降的趋势,说明抗氧化酶系统在油橄榄耐低温过程中作用明显,但当温度降低到0 ℃以下时,其抗氧化酶系统逐渐失去作用,从而导致活性降低。从本研究中抗氧化酶活性的上升幅度看,‘柯基’上升幅度均最小,说明抗氧化酶系统对低温的响应最不敏感,这可能是导致‘柯基’耐寒性表现不佳的原因之一。相反,耐寒性表现良好的‘豆果’和‘皮瓜尔’等抗氧化酶活性的上升幅度更大。为克服单一指标在耐寒性鉴定中的不足,试验采用8 个生理指标利用隶属函数法对其耐寒性进行综合评价,耐寒性强弱顺序为:‘豆果’>‘皮瓜尔’>‘科拉蒂’>‘莱星’>‘鄂植8 号’>‘阿尔波萨纳’>‘佛奥’>‘柯基’。
3.3 光系统对低温胁迫的响应
光系统的破坏程度也可以用来判定植物的耐寒性。随着低温胁迫加剧,‘鄂植8 号’和‘莱星’的qP呈下降趋势,NPQ呈上升趋势,说明低温胁迫严重破坏了‘鄂植8 号’和‘莱星’的光合电子传递途径,使得光合电子传递能力不断降低,用于光化学反应的光能部分减少,而以热形式耗散掉的光能部分不断增加。但其热耗散增加缓解了光抑制对其光合功能产生的破坏,增强了光合反应中心的稳定性,从而能减轻低温胁迫对其叶片光合器官的损伤。本研究通过测定8 种油橄榄幼苗的光合及叶绿素荧光等相关指标,并利用隶属函数法对其耐寒性做了初步评价,结果为:‘科拉蒂’>‘豆果’>‘柯基’>‘鄂植8 号’=‘佛奥’=‘莱星’>‘皮瓜尔’>‘阿尔波萨纳’。
3.4 耐寒性评价体系对比
低温是限制油橄榄在我国北亚热带扩区发展的主要因素。研究油橄榄幼苗膜系统、渗透调节物质、抗氧化酶系统、光系统对低温胁迫的响应,是低温适应性品种选育至关重要的一步。从评价结果来看,电导率拟合半致死温度评价方法和耐寒相关生理指标隶属函数法评价结果接近,更符合各品种采样区越冬情况。而以光合和叶绿素荧光指标体系的耐寒性评价方法与前2 种方法评价结果差异较大,可能与油橄榄叶片近轴表皮角质膜较厚,蜡质结晶致密,气孔密度小等叶片重要形态特征有关[23]。因此评定油橄榄耐寒性时,需多方面综合评价,同时结合田间观察,才能使结果更加准确、科学。
4 结论
通过Logistic 方程拟合半致死温度,渗透调节物质、保护酶活性参数,光合和叶绿素荧光参数等指标分析,对8 个油橄榄品种进行耐寒性评价,发现油橄榄幼苗膜系统、渗透调节物质、抗氧化酶系统、光系统对低温胁迫的响应并不一致。渗透调节物质、保护酶活性参数隶属函数法所得出耐寒性综合排序与应用电导法配合Logistic 方程求出半致死温度的排序基本一致,且符合各品种田间越冬表现,该方法更适合作为油橄榄苗期耐寒性评价方法。利用光合及叶绿素荧光参数10 个指标的隶属函数评价方法,很难准确评价油橄榄苗期耐寒性,尤其是难以区分耐寒性差异较小的品种。
