南亚热带乡土阔叶林与马尾松林土壤细菌群落多样性和结构
2024-01-20滕金倩肖指柔明安刚席守鸿朱茂锋
滕金倩,肖指柔,明安刚,朱 豪,席守鸿,何 江,朱茂锋,陶 怡,覃 林*
(1.广西大学林学院,广西森林生态与与保育重点实验室,广西 南宁 530004;2.中国林业科学研究院热带林业实验中心,广西 凭祥532600;3.广西友谊关森林生态系统定位观测研究站,广西 凭祥 532600)
为了满足木材、燃料和其它林产品的需求,将天然阔叶林改造成速生丰产的人工针叶林是过去几十年里中国南方普遍采用的经营策略[1]。然而,大面积多代连栽的针叶林极易造成一系列严重的生态环境问题(如土壤侵蚀、肥力衰退和生态系统稳定性降低等),最终导致人工林生产力的不断下降[2]。如何科学培育人工林,有效提高其生产力并维持生物多样性,是人工林多目标经营亟需解决的关键问题。有研究表明,将现存的大面积低质低效针叶纯林转化为经济价值、生产力和物种多样性高的乡土阔叶树种人工林,将会改变土壤理化性质和养分循环,有助于提高土壤肥力[3]。将针叶林改造成乡土阔叶林,是近年来我国最有前景的人工林经营模式之一。
土壤细菌群落是陆地生态系统中最多样化和最主要的微生物群落,对生态系统营养物质循环、分解有机质及维持土壤肥力等具有关键效用[4]。凋落物与根系分泌物能为土壤细菌生命活动提供所需要的碳源,因而不同树种可直接引起土壤细菌群落组成多样性的变化[5]。土壤细菌群落多样性包括分类和系统发育两方面。传统的分类多样性仅仅量化物种名称和丰度变化,没有传达有关物种的生态和系统发育的关键信息,无法解释群落动态,因此系统发育多样性的变化越来越受到人们的关注[6]。系统发育多样性反应了群落内物种之间的系统发育距离和作用,有助于估计生物多样性对生态系统功能的影响[7]。群落结构是物种在不同空间尺度上相互作用的结果,量化物种分布模式的空间差异可以深入了解调控物种多样性形成的机制以及群落趋同、转换率等群落状态[8]。有研究表明,中国南方地区的针叶林和阔叶林的土壤细菌群落分类结构差异显著[9]。目前,生态位理论和中性理论分别代表了群落构建的确定性过程(环境过滤和竞争排斥)和随机性过程(不可预测的干扰、扩散、出生和死亡)[10]。有学者使用群落分类结构的Raup-Crick相异指数(βRC)探究群落构建过程,发现环境过滤是中国华北亚高山林线区域土壤细菌群落构建驱动因素[11]。同时,通过系统发育结构也可以反映群落构建过程中物种的进化历史,若由环境过滤主导,则筛选出适应能力相似、亲缘关系较近的物种,导致系统发育聚类;若由竞争排斥主导,则生态位相似的物种将无法共存,导致系统发育过度分散[12]。至今关于森林土壤细菌群落系统发育结构的研究结果不一。有研究认为,森林土壤(0~10 cm)细菌群落系统发育结构是聚集的,群落构建由确定性过程决定,主要受环境过滤影响[13-14];但也有研究指出,确定性过程和随机性过程共同作用于森林土壤(0~20 cm)细菌群落构建,其中确定性过程占主要作用[15]。另外,土壤细菌对生物地球化学循环过程的影响在整个土壤剖面中一直存在[16],且不同土层深度的土壤理化性质不同[9],或许会导致不同的细菌群落分布模式。然而,现有的研究仅仅考虑了森林表层土壤(0~20 cm)细菌群落构建机制,对于人工林深层土壤(>20 cm)的细菌群落如何构建知之甚少。
我国亚热带地区人工林的面积和蓄积均居全国之首[17]。用红锥(Castanopsis hystrixMiq.)、米老排(Mytilaria laosensisLec.) 和火力楠(Michelia macclureiDandy)等珍贵乡土阔叶林更替杉木(Cunninghamia lanceolata(Lamb.)Hook.)和马尾松(Pinus massonianaLamb)等针叶林是目前该地区常见的营林模式。针叶林改建成阔叶林后,其凋落物数量与质量的改变将会对土壤理化性质产生作用,进而会影响土壤细菌群落多样性和结构。本研究以南亚热带地区杉木林采伐迹地种植的马尾松、红锥、米老排和火力楠等人工林为对象,基于各林分不同土层(0~20、20~40 和40~60 cm)细菌16S rRNA 高通量测序数据,旨在通过差异性分析推测未来在马尾松林采伐后种植乡土阔叶林情况下土壤细菌群落的分类与系统发育多样性和结构依土层的变化特征,以及导致变化的主要土壤理化因子。研究结果不仅对针叶林改建乡土阔叶林的树种选择具有指导作用,还为阐明亚热带人工林土壤剖面细菌群落构建机制提供科学依据。
1 材料与方法
1.1 研究区概况及样地设置与样品采集
2017 年2 月在该区域选取具有立地条件(土壤类型、海拔、坡度、坡向)和经营措施相似的马尾松、红锥、米老排和火力楠等4 个乡土树种人工林为对象。4 个人工林均是在杉木采伐迹地上种植(密度为2 500 株·hm-2);其中,马尾松林和红锥林于1983 年营造并经过了3 次间伐,而米老排林和火力楠林于1981 年营建并经历2 次间伐,间伐强度约30%。在各林分内,随机设置3 个20 m × 20 m 的样地(样地间距20 m 以上),在每个样地内沿左对角线任意选择3 个采样点,用土钻(内径5.0 cm)分别采集表层(0~20 cm)、中层(20~40 cm)和底层(40~60 cm)土样,除去动植物残体与石块等杂质后,将同层土样混合成一个土样装入无菌采样袋,置于内含生物冰袋的泡沫箱中并带回实验室。新鲜混合土样过2 mm 钢筛后分为3 份:1 份置于-80 ℃的冰箱内冷藏,用于土壤DNA 提取;1 份置于冰箱4 ℃保存,用于测定土壤硝态氮和铵态氮含量;1 份经自然风干后过0.25 mm 筛保存用于测定土壤基本化学性质。各林分的立地条件和基本特征见表1。
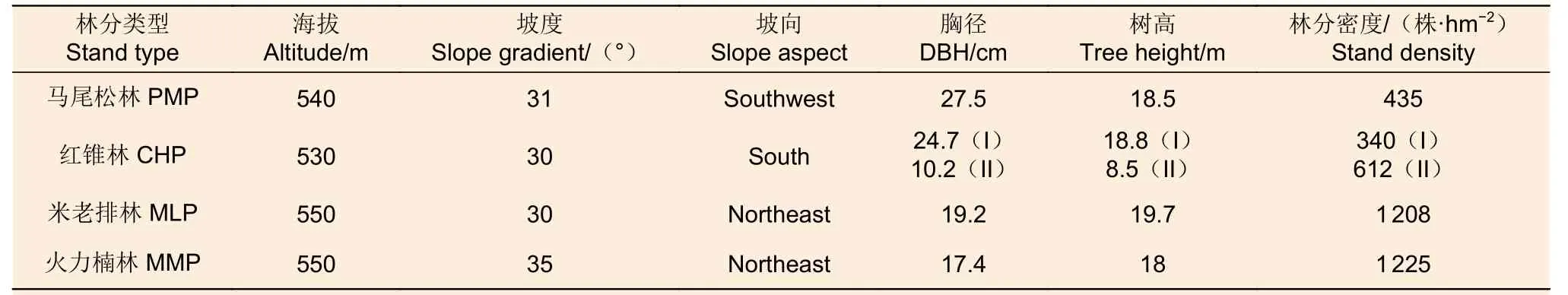
表1 4 个人工林的立地条件和林分特征Table 1 Site conditions and stand characteristics of the four planted forests
1.2 土壤理化性质测定与高通量测序
参照鲁如坤[19]测定如下指标:烘干法测定土壤含水量(SWC);pH 计(Prtavo 907 MULTI pH,德国)测定土壤pH 值(土∶水=1∶2.5);重铬酸钾法测定土壤有机碳(SOC)含量;酚二磺酸比色法测定硝态氮()含量;扩散法测定铵态氮()含量;双酸浸提-钼锑抗比色法测定速效磷(AP)含量。经H2SO4与HClO4消煮后,使用全自动间断化学元素分析仪SmartChem200(Alliance,法国)测定总氮(TN)和总磷(TP)含量;土壤有机碳与总氮含量的比值即土壤碳氮比(C/N)。
土壤DNA 使用 PowerSoil®DNA Isolation Kit 试剂盒(美国MoBio 公司)提取。提取到的土壤DNA 纯度和浓度通过紫外微量分光光度计(Thermo Nano Drop 2000)进行测定。以515F( 5 ′-GTGCCAGCMGCCGCGGTAA-3 ′ )/926R(5′-CCGTCAATTCMTTTGAGTTT-3′)为引物[20],通过PCR 扩增技术进行细菌16S rRNA V4-V5 区基因片段的扩增。两次PCR 扩增的条件和反应体系见彭雯等[21]。对PCR 产物进行2%琼脂糖凝胶电泳检测,使用美国AXYGEN 公司的AxyPrepDNA凝胶回收试剂盒对凝胶进行回收纯化,通过FTC-3000TM real-time PCR 仪实现胶回收产物的荧光定量,均一化处理后测序文库构建结束。由上海微基生物科技有限公司通过Illumina Miseq 2x300 bp完成高通量测序。
根据barcode 信息对高通量测序得到的原始序列进行拼接、质量控制和过滤。有效序列拼接由FLASH 软件完成,质控和过滤由Trimmomatic软件实现,优化序列通过Mothur V.1.33.3 软件获得。对优化序列基于97% 相似水平的聚类由UPARSE 软件完成, 生成OTU (operational taxonomic unit)的特征序列,用UCHIME 软件检测和去除嵌合体序列,使用Mothur(classify.seqs)软件比对OTU 特征序列与Silva(Release 123, http://www.arb-silva.de)数据库后进行物种注释(置信度阈值设置为0.8)。
以最低样本测序深度17 291 为基准进行抽平,结果用于后续分析。采用最大似然法构建系统发育树,在QIIME2 2 021.8 软件中的FASTTREE程序完成[22]。
1.3 土壤细菌群落分类与系统发育多样性、结构及转换率
1.3.1 细菌群落分类与系统发育多样性 采用Chao1、Shannon 和Simpson 指数反映细菌群落的分类多样性;PD 指数[23]表征系统发育多样性,其数值大小取决于物种共同祖先的多寡(即系统发育树的节点数量)。
1.3.2 细菌群落分类与系统发育结构 利用基于Bray-Cruits 距离的主坐标分析(PCoA)和置换多元方差分析(PERMANOVA)检测不同林分土壤细菌群落分类结构之间的差异,并用Monte Carlo检验理化性质的显著性。用Raup-Crick 相异指数(βRC)表示两个群落之间的分类结构相异程度,其取值范围为[-1,1],可表征群落的构建过程[24]。使用净亲缘关系指数(NRI)和最近亲缘关系指数(NTI)量化系统发育结构[25],从而揭示群落构建机制。
1.3.3 细菌群落分类与系统发育转换率 为了测量不同林分土壤细菌群落分类和系统发育的空间动态变化,使用Bray-Cruits 距离表征群落分类转换率,使用丰度加权的平均成对系统发育距离(βMPD)和平均最近相邻系统发育距离(βMNTD)量化群落系统发育转换率[26]。以上分析均在R 软件中进行。
总的来说,初中语文的作文教学是提升学生写作能力的重要关键,教师应当重视自身教学中存在的问题,在解决这些问题与困境的基础上,多让学生以自己的视角进行人与事的分析,这样才能够让学生的写作更有内涵,同时也能够提升他们的实际写作感悟,进而保障他们写作能力的提升。
1.4 统计分析
采用单因素方差分析(One-way ANOVA)及多重比较(Tukey)判定相同土层不同林分之间土壤理化性质、细菌群落的分类与系统发育多样性以及分类与系统发育转化率的差异。采用Pearson相关方法,探讨土壤理化性质与分类和系统发育多样性之间的相关性。利用SPSS 26.0 软件(SPSS, Inc, Chicago, IL)完成计算。
2 结果与分析
2.1 马尾松林与乡土阔叶林土壤理化性质的比较
方差分析表明(表2),就相同土层不同林分而言,在土壤表层,马尾松林的SWC 显著低于米老排林和火力楠林;在中层,马尾松林的SWC 显著低于米老排林和火力楠林以及TN 显著低于火力楠林,但其pH 和均显著高于红锥林和米老排林;在底层,马尾松林的pH 显著高于火力楠林,而其显著高于红锥林和米老排林(P<0.05)。土壤SOC、、TP、AP 及C/N 在相同土层不同林分间均无显著差异(P>0.05)。
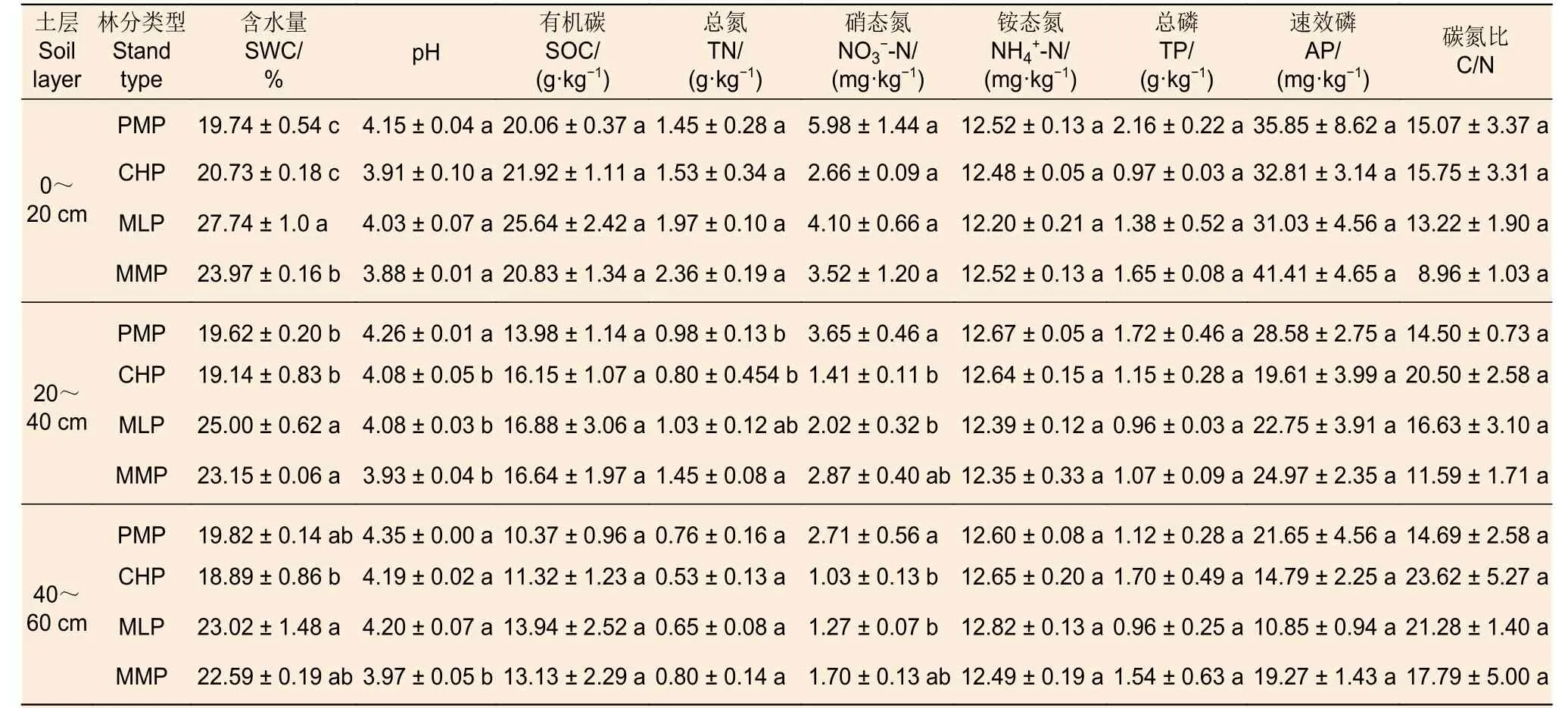
表2 4 个研究林分的土壤理化性质(均值 ± 标准误,n=3)Table 2 Physical and chemical properties of soil in four plantations (mean ± SE, n=3)
2.2 马尾松林与乡土阔叶林土壤细菌群落多样性的差异
方差分析表明,在土壤表层,马尾松林的Chao1、PD 指数显著低于米老排林和火力楠林(P<0.05),而与红锥林差异不显著(P>0.05)(图1A、D),其中,相较于马尾松林,米老排林的Chao1、PD 指数分别提高了20.5%和20.7%,火力楠林则提高了23.8%和20.6%;马尾松林与3 乡土阔叶林的Shannon 和Simpson 指数均无显著差异(图1B、C)。在中层和底层,马尾松林与3 个乡土阔叶林的分类和系统发育多样性指数均无显著差异(P>0.05)(图1)。
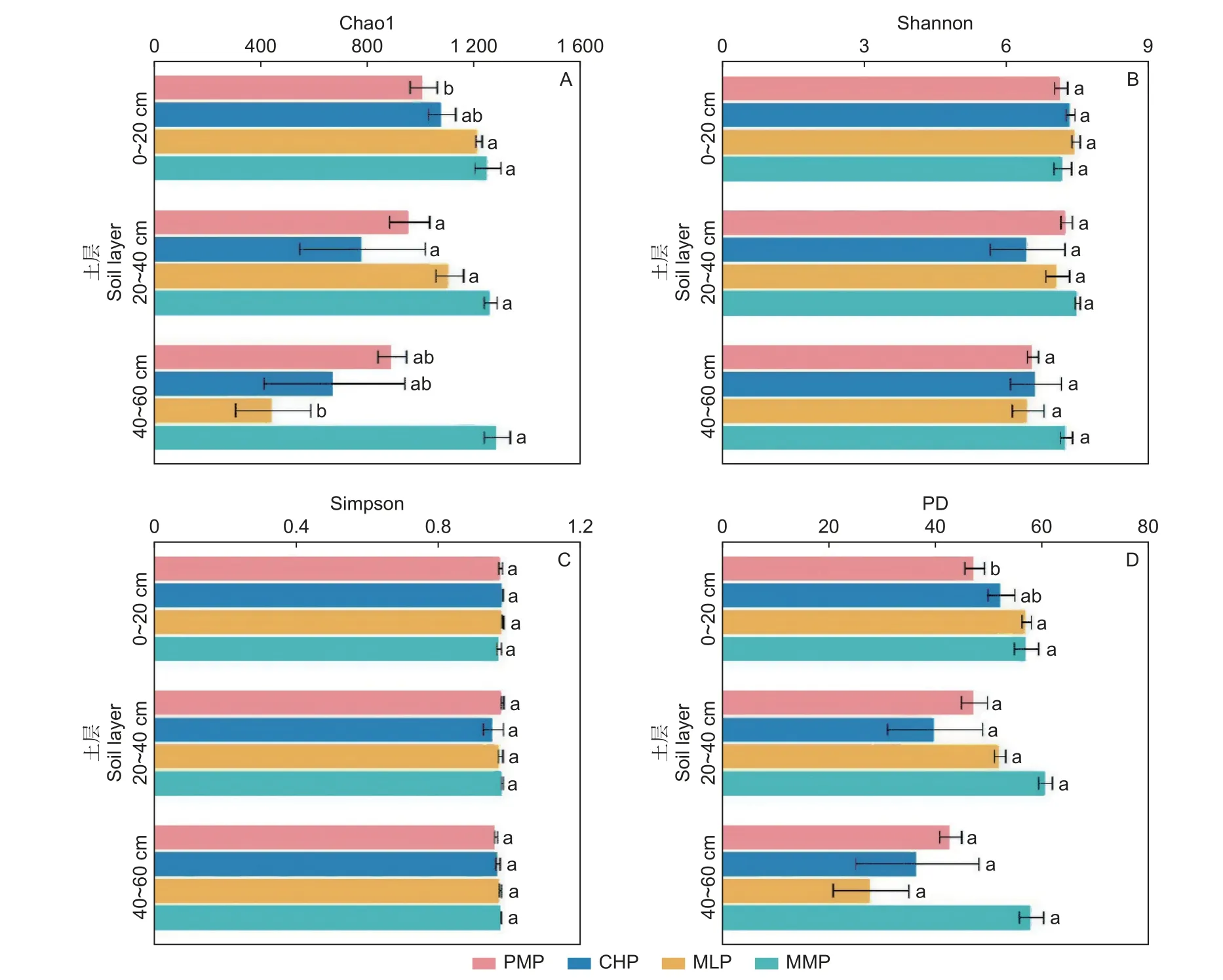
图1 4 个人工林土壤细菌群落的分类与系统发育多样性(均值 ± 标准误,n=3)Fig.1 Taxonomic and phylogenetic diversity of soil bacterial communities in four plantations (mean ± SE, n=3)
土壤理化性质与土壤细菌群落多样性指数的Pearson 相关性分析表明(图2),Chao1 和PD 在表层分别与SWC 显著正相关;在中层分别与TN 显著正相关且与C/N 显著负相关;在底层均与9 个理化因子无显著相关性。Shannon 和Simpson 在表层与呈显著负相关,且Simpson 还与C/N 显著正相关;在中层分别与C/N 呈显著负相关;在底层,Simpson 与pH 显著负相关(P<0.05),而Shannon 与9 个理化因子均无显著性相关。
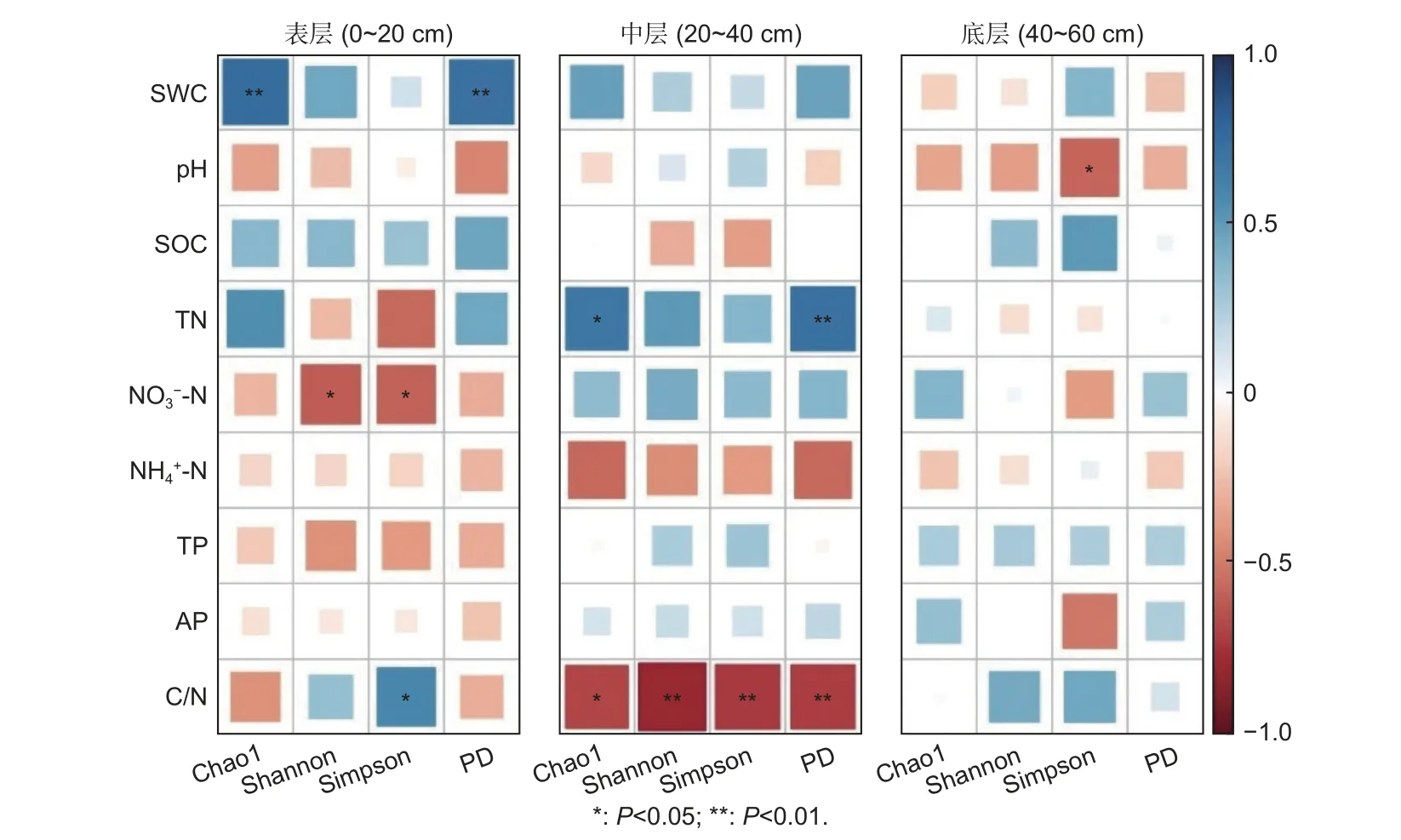
图2 全部研究林分土壤理化性质与细菌群落分类及系统发育多样性的相关性热图Fig.2 Correlation heatmap of the soil physicochemical properties and bacterial community diversity(taxonomic and phylogenetic) in all plantations
2.3 马尾松针叶林与乡土阔叶林土壤细菌群落结构的变化
2.3.1 细菌群落分类结构 在土壤表层,主坐标分析(PCoA)细菌群落分类结构总解释度为54.4%(图3),PERMANOVA 分析显示马尾松林分别与红锥林(Fpseudo=3.1,P=0.017)、米老排林(Fpseudo=2.9,P=0.025)、火力楠林(Fpseudo=2.0,P=0.039)有显著差异。dbRDA 结果表征(图4),前两轴累计解释了58.2% 的样品细菌群落分类结构与土壤理化性质因子的关系;马尾松林土壤样品位于RDA2 的负方,而红锥林、米老排林和火力楠林土壤样品几乎位于RDA2 的正方;Monte Carlo 检验进一步表明,土壤TP、SWC、TN 是制约马尾松林与红锥林、米老排林、火力楠林土壤细菌群落分类结构差异的环境因素(P<0.05)。
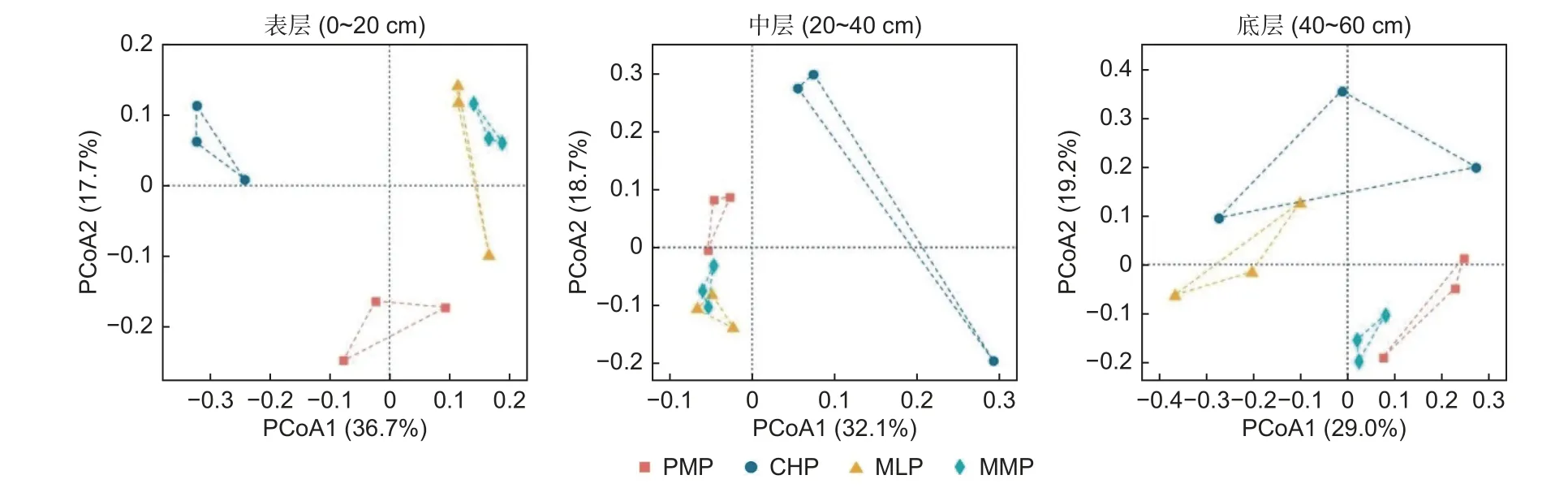
图3 马尾松林与3 个乡土阔叶林土壤细菌群落分类结构的主坐标分析(PCoA)Fig.3 Principal Coordinate Analysis (PCoA) of soil bacterial community taxonomic structure between Pinus massoniana plantation and three broadleaved plantations

图4 土壤理化性质对4 个人工林细菌群落分类结构影响的冗余分析(RDA)Fig.4 Redundancy analysis (RDA) on effects of soil physical and chemical properties on bacterial community taxonomic structure of four plantations
在中层,PCoA1 和PCoA2 的解释度分别为32.1%和18.7%(图3),且PERMANOVA 分析表明马尾松林仅与红锥林有显著差异(Fpseudo=2.3,P=0.012)。dbRDA分析表明(图4),dbRDA1解释度为36.3%,dbRDA2 解释度为21.3%;马尾松林土壤样品处于RDA1 的负侧,而红锥林土壤样品处于RDA1 的正侧;Monte Carlo 检验揭示土壤C/N 是制约马尾松林与红锥林土壤细菌群落分类结构差异的环境因素(P<0.05)。
在底层,PCoA 分析的前两轴累计解释度为48.2%(图3),PERMANOVA 发现马尾松林与3 个阔叶林均无显著差异(P>0.05)。dbRDA 前两轴的总解释度为51.3%(图4),Monte Carlo检验表明土壤和SWC 是影响不同土壤样品细菌群落分类结构差异的主要因素(P<0.05)。
另外,相同土层相同林分的两两土壤样品之间的βRC均为-1,意指环境过滤在4 个人工林不同土层的细菌群落构建中均起主导作用。
2.3.2 细菌群落系统发育结构 4 个林分在各土层的细菌群落净亲缘关系指数NRI(图5A)和最近亲缘关系指数NTI(图5B)均大于1.96,表现为系统发育聚集,说明确定性过程在土壤细菌群落构建中起重要作用,环境过滤导致系统发育聚类。

图5 4 个人工林在不同土壤深度的细菌群落系统发育结构指数(均值 ± 标准误,n=3)Fig.5 Phylogenetic structure index of bacterial communities in the four plantations at different soil depths (mean ± SE, n=3)
2.4 马尾松林改建成乡土阔叶林的土壤细菌群落分类与系统发育转换率
2.4.1 细菌群落分类转换率 马尾松林转换为3 个阔叶林的土壤细菌群落分类转换率的差异因土层不同(图6A)。在0~40 cm 土层,马尾松林转换为红锥林的转换率显著高于转换为米老排林和火力楠林;在40~60 cm,马尾松林转换为红锥林和米老排林的转换率均显著高于转换为火力楠林(P<0.05)。
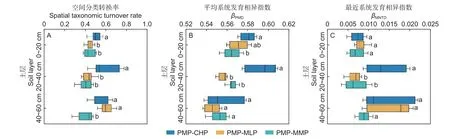
图6 马尾松林转换为3 阔叶林的土壤细菌群落分类和系统发育转换率Fig.6 Taxonomic and phylogenetic turnover rates of soil bacterial communities from PMP to three broadleaved plantations
2.4.2 细菌群落系统发育转换率 土壤细菌群落平均系统发育相异指数βMPD(图6B)和最近系统发育相异指数βMNTD(图6C)的方差分析表明,在土壤表层,马尾松林转换为红锥林的βMPD显著大于转换为火力楠林(P<0.05),而马尾松林分别转换为3 个阔叶林的βMNTD无显著差异(P>0.05);在中层,马尾松林转换为红锥林的βMPD和βMNTD都显著大于转换米老排林和火力楠林(P<0.05);在底层,马尾松林转换为3 个阔叶林的βMPD、βMNTD均无显著差异。
3 讨论
3.1 针叶林与阔叶林土壤细菌群落多样性
土壤细菌群落分类多样性是表征细菌在生态系统中功能的重要指标[27],分类多样性越高,则越有利于土壤的可持续利用和抵抗力[28]。有研究指出,树种变化是影响土壤细菌群分类多样性的原因[29-30]。由于针叶与阔叶树种的凋落物和根系对土壤传输的养分不同,改变了土壤理化性质,从而影响土壤细菌群落分类多样性。本研究发现,米老排林和火力楠林相比于马尾松林,土壤表层的细菌群落Chao1 指数有显著提高(图1A),而中层和底层土壤细菌群落的分类多样性(Chao1、Shannon 和Simpson 指数)没有明显差异,说明树种转换导致土壤细菌群落分类多样性的变化也会因土壤深度而异。米老排林和火力楠林表层土壤细菌群落Chao1 指数高于马尾松林,其原因在于其表层土壤SWC 显著高于马尾松林(表2),而表层土壤细菌群落Chao1 指数与土壤SWC 显著正相关(图2A)。细菌在相对较高的水势环境下活性更高[31],SWC 高的土壤能够为土壤细菌的生长提供了一个稳定而潮湿的环境促进其生长繁殖。
系统发育多样性可以表示物种的差异性和生态位[32],系统发育多样性越高的生态系统越稳定,且群落中的细菌相互作用越弱[33]。细菌以非常小的尺度(very small scale)栖息在土壤生态位中,其系统发育多样性通常与土壤理化性质密切相关[34]。本研究发现,系统发育多样性与分类多样性呈相同的分布规律,即相较于马尾松林,米老排林和火力楠林表层土壤细菌群落系统发育多样性有显著增加(图1D),而中层和底层无明显变化。Taketani等[35]研究指出,土壤SWC 对细菌群落系统发育多样性具有至关重要的作用,土壤SWC 降低会直接引起系统发育多样性减少。本研究也发现4 个林分土壤表层SWC与PD极显著正相关(P<0.01,图2A),因此,于马尾松林而言,米老排林和火力楠林表层土壤SWC 的显著增加是导致细菌群落系统发育多样性显著提高的主要缘由。尽管土壤pH 值被普遍认为是影响土壤细菌群落系统发育多样性的重要因素[36],但由于本研究中红锥林、米老排林、火力楠林和马尾松林表层土壤的pH 值无显著差异(3.88~4.15),因而未发现土壤pH 值对土壤细菌群落系统发育多样性产生明显影响。
3.2 针叶林与阔叶林土壤细菌群落结构
针叶林与阔叶林的土壤细菌群落分类结构不同[30],这是多种环境因子综合作用的结果。Gunina等[37]研究表明,造林树种不同导致其土壤理化性质差异,最终影响其土壤细菌群落分类结构。土壤TN[38-39]、TP[40]和C/N[41]是调节森林土壤细菌群落分类结构的重要因素。本研究的RDA 分析结果较好的反映了理化因子与土壤细菌群落分类结构的关系表层土壤SWC 增加、TN 增加和TP 减少是驱动马尾松林分别与3 个阔叶林土壤细菌群落分类结构具有显著差异的重要因素;而中层土壤C/N 增加是导致马尾松林与红锥林土壤细菌群落分类结构差异显著的主要因子;但在底层马尾松林与3 个阔叶林土壤细菌群落分类结构都没有显著差异(图3、图4)。可见,马尾松林与3 个乡土阔叶林土壤细菌群落分类结构的差异因土层而异,这是因为人工林凋落物累积在土壤表层,且针叶林与阔叶林的凋落物分解率不同[42],表层土壤理化性质差异较大,但随着土层加深,针叶林与阔叶林的理化性质差异缩小,使得细菌群落分类结构逐渐趋同。
微生物群落构建过程一直是群落生态学关注的热点[43],然而由于多种生态过程的作用使因果关系的确定复杂化。βRC可以作为定量估计随机与确定性过程在微生物群落构建中贡献的指数[44],系统发育结构的变化也已被证明可以用于评估微生物群落构建过程中环境过滤、竞争排斥和随机过程的重要性[45]。本研究的βRC、NRI 和NTI 指数表明,马尾松林与3 个阔叶林不同土层的土壤细菌群落构建均受环境过滤主导,这表明针叶与阔叶树种的更替不会造成土壤细菌群落构建机制的改变。这与华北亚高山林线区域土壤[11]、不同林龄的杉木人工林土壤(0 ~10 cm)[13]及不同时间历史的阔叶糖枫(Acer saccharumMarshall)天然林土壤有机质层[46]细菌的研究结果一致,认为是环境过滤调控细菌群落构建。此外,在土壤表层和中层,马尾松林转换为红锥林的物种组成和系统发育结构转换率高于转换为米老排林和火力楠林(图6),本文推测未来将马尾松林改造成红锥林后的土壤细菌种类更替比转换成米老排林和火力楠林更频繁,红锥林的土壤细菌群落稳定性较差,细菌间的相互作用更强烈[33]。
4 结论
本文推测南亚热带地区的马尾松林改建成阔叶林后,米老排林和火力楠林土壤表层细菌群落的分类与系统发育多样性显著提高了20.5%~23.8%,土壤SWC 是影响的主导因子;3 个阔叶林土壤细菌群落分类结构都在表层产生了显著的变化,而在中层只有红锥林有明显改变,主要受土壤SWC、TP、TN 和C/N 影响;不同土层的细菌群落构建主导因素没有发生改变,均是环境过滤;在土壤表层和中层,马尾松林转变为米老排林及火力楠林之间的分类和系统发育转换率都低于转变为红锥林,米老排林和火力楠林的土壤细菌群落比红锥林更稳定。基于土壤细菌群落分类与系统发育的多样性及转换率角度,该地区将马尾松林改建成米老排林或火力楠林比红锥林更能提高土壤细菌群落的稳定性。未来可开展针叶林转换乡土阔叶林后土壤细菌群落功能变化的研究,有助于深入理解林分转换介导土壤生态功能改变的调控机制。
