金银花黑斑病病原菌番茄匍柄霉的分离与鉴定
2024-01-20刘端冲杨金库林若竹姚艳霞淮稳霞赵文霞
刘端冲,杨金库,林若竹,姚艳霞,淮稳霞*,赵文霞*
(1.国家林业和草原局森林保护学重点实验室 中国林业科学研究院森林生态环境与自然保护研究所,北京 100091;2.廊坊市农林科学院,河北 廊坊 065000)
金银花学名忍冬(Lonicera japonicaThunb.),是一种重要的药用植物,具有广泛的药理作用,如抗菌、抗炎、抗病毒、抗内毒素、减脂、解热等[1-2]。金银花在国内分布十分广泛,其种植区域主要集中在山东、河北和河南等省份,其中河北省巨鹿县为全国金银花三大主产地(河北巨鹿、河南封丘、山东平邑)之首,成为当地的支柱产业,获得“中国金银花之乡”的称号[3-4]。
金银花叶部病害是田间生产的巨大障碍,主要有褐斑病(Cercospora rhamniFuckel)、白粉病(Microsphaera liniceraeWint.inRabenh)等。其中,金银花褐斑病对金银花的产量与质量有严重影响。此前,有报道认为金银花褐斑病的病原菌是鼠李尾孢(C.rhamni)[5-6],也有研究认为是多主棒孢霉(Corynespora cassiicola(Berk.& M.A.Curtis) C.T.Wei)[7-8];2014 年,de Miranda 等人的研究表明假尾孢属真菌(Pseudocercospora lonicerigenaU.Braun & Crous)是引起巴西金银花褐斑病的病原菌[9];2016 年张永信等人的研究发现拟茎点霉属真菌(Phomopsissp.)也可以使金银花产生褐斑病[10]。
2014 年到2018 年期间,笔者在研究河北金银花褐斑病病原过程中,总能分离到一种产棕黄色或红色色素的真菌,其分出率约为17%。利用离体叶片致病性接种时,该菌产生的病斑与褐斑病不符,并依据ITS 序列和形态特征将其初步鉴定为匍柄霉(Stemphyliumsp.)。针对这种金银花新型叶斑病,本文依照科赫氏法则对其病原菌进行了致病性测定,并结合形态学和多基因片段分析对其进行了分类鉴定,并将该病害命名为金银花黑斑病,以期为该病害的有效防控提供理论基础。
1 材料与方法
1.1 样品采集及病原菌的分离纯化
本项研究所用金银花病叶样采集自河北省巨鹿县和廊坊市。样本利用自封袋包装并按野外样本采集要求记录采集时间、地点、采集人、环境条件等要素后,带回实验室用于病原菌分离鉴定。
病原菌的分离采用常规组织分离法。首先将采集的叶片用无菌水冲洗干净,用剪刀将病叶的病健交界处剪为4 mm × 4 mm 的小组织块,于超净工作台上用75%酒精浸泡30 s;随后用无菌水淋洗,再用0.1%升汞溶液浸泡2 min,无菌水洗3 次;用无菌滤纸吸去叶组织块表面的水分后,再将其摆放至PDA 培养基(马铃薯200 g、琼脂20 g、葡萄糖20 g、蒸馏水1 L)平板上进行分离培养。每个平板均匀摆放6 个组织块,于25 ℃下恒温培养24 h 后,挑取已长出菌落的边缘菌丝进行菌株的纯化,从而获得纯菌株。
1.2 形态学观察
将纯化后的菌株在PDA 培养基上培养4 d后,分别转接到新的PDA 和PCA(马铃薯和胡萝卜各100 g, 葡萄糖20 g,琼脂20 g,蒸馏水1 L)培养基上,于25 ℃下恒温培养7 d,观察并记录菌落的形状、颜色、光泽度、质地等培养特征。分别采用番茄快速诱导产孢法和双玻片插片诱导产孢法诱导该菌株产孢[11-12],菌株的显微特征主要包括菌株分生孢子的大小、形状,分生孢子梗的形状等特征,并拍照。
番茄快速诱导产孢法:将接种于PDA 上的匍柄霉菌在恒温培养箱中按照12 h 光照12 h 黑暗光周期在温度25 ℃下培养至菌丝长满培养皿;取菌丝生长边缘的匍柄霉菌片接种到经过催芽处理新长出的番茄叶片上,在温度25 ℃、相对湿度60%~80%与12 小时光照12 小时黑暗光周期的条件下培养3 d,挑取匍柄霉菌片表面在显微镜下观察。
双玻片插片诱导产孢法:于25 ± 1 ℃培养箱中恒温培养3~4 d 后,取3~5 个平板于菌落边缘1~2 cm 处以30°~45°角斜插入洁净的双层重叠的盖玻片,然后放置 25 ± 1 ℃培养箱恒温培养3~4 周左右。用镊子取出诱导培养3~4 周后的双层玻片,并轻轻分成2 个单层玻片,单层玻片的正面载有菌体,在显微镜下观察。
1.3 DNA 提取、PCR 扩增及系统发育学分析
所获得的纯菌株于PDA 培养基上培养10 d后,刮取少量菌丝置于2 mL 离心管中,采用改良的CTAB 法提取本研究所用菌株的DNA[13],利用引物ITS1(5′-TCCGTAGGTGAACCTGCGG-3′)/ITS4(5′-TCCTCCGCTTATTGATATGC-3′)、引物gpd1(5′-CAACGGCTTCGGTCGCATTG-3′)/gpd2(5′-GCCAAGCAGTTGGTTGTGC-3′)和引物EF446F(5′-TCACTTGATCTACAAGTGCGG TGG-3′)/EF1473R(5′-CGATCTTGTAGA CAT CCTGGAGG-3′)分别扩增核糖体内转录间隔区(ITS)、3-磷酸甘油脱氢酶(gpd)和蛋白延伸因子(EF-1α)基因片段[14-16]。ITS 基因片段反应体系为(25 μL):12.5 μL 2 × Taq PCR Master Mix(天根生化科技(北京)有限公司),9.5 μL ddH2O, 1 μL ITS1, 1 μL ITS4 和1 μL DNA 模板。gpd基因片段反应体系(25 μL):12.5 μL 2 ×Taq PCR Master Mix,9.5 μL ddH2O, 1 μLgpd1,1 μLgpd2 和1 μL DNA 模板。EF-1α 基因片段反应 体 系 为(25 μL):12.5 μL 2 × Taq PCR Master Mix,9 μL ddH2O, 1 μL EF446F, 1 μL EF1473R 和1.5 μL DNA 模板。ITS 序列PCR 反应程序为:94 ℃预变性5 min,94 ℃变性30 s,54 ℃退火 30 s,72 ℃延伸 30 s,35 个循环,最后72 ℃延 伸10 min,4 ℃保 存。gpd序列PCR 反应程序为:94 ℃预变性3 min,94 ℃变性30 s,57 ℃退火 1 min,72 ℃延伸 1.5 min,35 个循环,最后72 ℃延伸10 min,4 ℃保存。EF-1α 序列反应程序为:94 ℃预变性1 min,94 ℃变性30 s,60 ℃退火 1 min,72 ℃延伸 3 min,35 个循环,最后72 ℃延伸5 min,4 ℃保存。PCR 扩增产物于1.2% 琼脂糖凝胶电泳检测后,送至北京擎科生物公司进行测序。
测序所获得的拼接序列在NCBI 中进行比对,确定菌株分类地位,并从GenBank 中下载相似性高的序列或近缘物种序列作为参考序列(表1)。使用MEGA 10.2.2 软件[17]进行序列对齐和剪切,以ITS-gpd-EF-1α 的顺序进行序列串联,并分别利用最大似然法(ML)、贝叶斯法(BI)和最大简约法(MP)进行系统发育树的构建,外群为互隔链格孢菌Alternaria alternataEGS34-016。
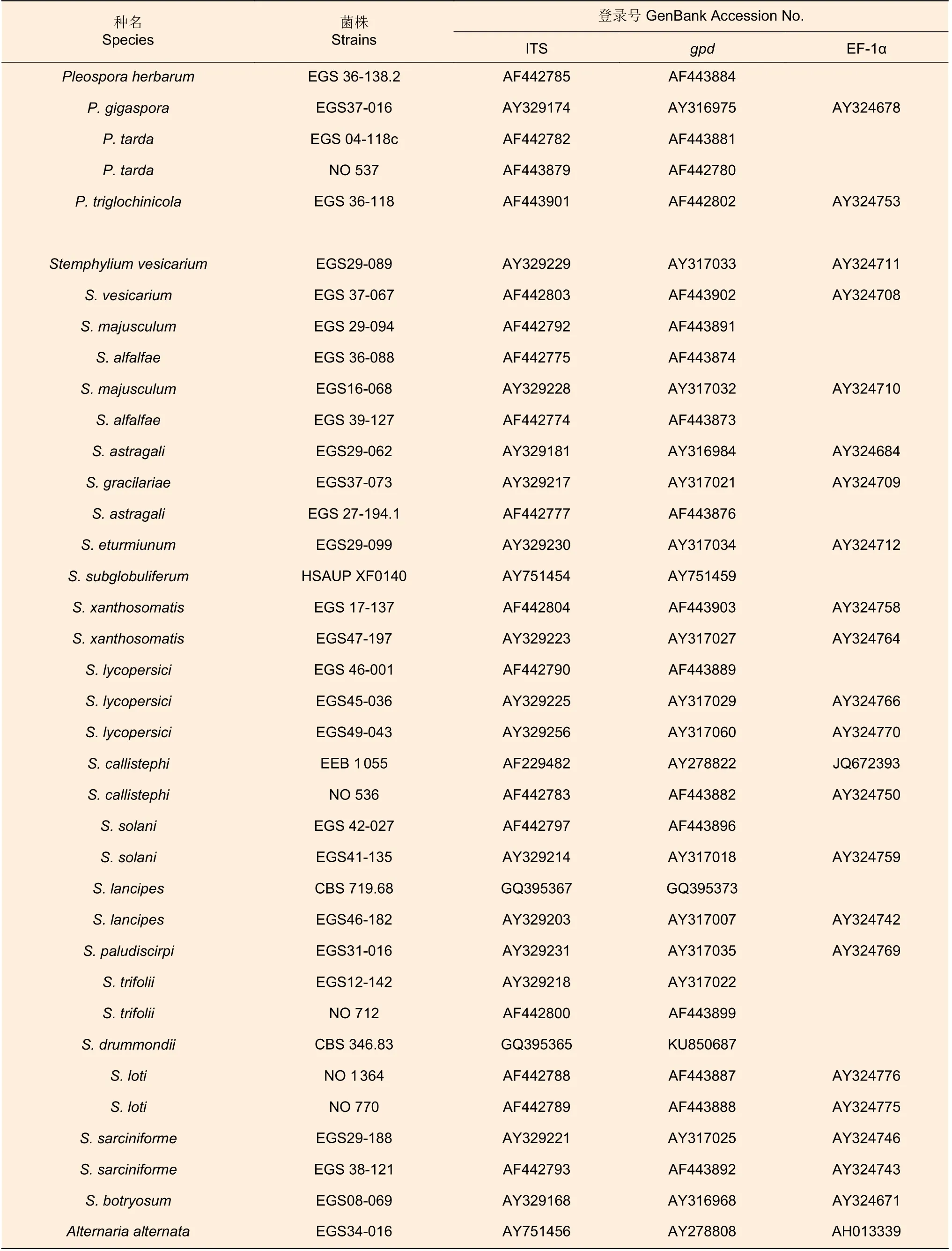
表1 用于分子系统发育分析的参考菌株信息Table 1 The information of strains used for phylogenetic analysis
1.4 致病性检测
本研究分别对金银花和番茄的叶片进行致病性接种实验。
金银花叶片接种:采用刺伤接种菌丝块的方法对健康的金银花活体叶片进行致病性测定。首先将纯菌株培养4~5 d 后打取菌落边缘直径为5 mm的菌饼;用75%酒精对叶片进行表面消毒,用接种针刺伤接种部位,菌饼接种于刺伤处,以无菌PDA 琼脂块接种作为对照,每组处理设10 次重复。接种后于25 ℃下恒温放置9 d,观察并记录发病状况。随后,通过常规组织分离法,取病健交界处进行病原菌的再次分离和纯化;通过菌株菌落形态结合ITS 序列,确定菌株是否与接种菌株相同。
番茄叶片接种:采用刺伤接种菌丝块的方法对健康的番茄离体叶片进行致病性测定,具体步骤同上。接种后于25 ℃下恒温放置48 h,观察并记录发病情况,并进行病原菌的再次分离、纯化和复核鉴定。
2 结果与分析
2.1 病害症状
在田间调查和实验中,该病害主要危害金银花的叶片,发病初期(5—8 月份),通常在叶片边缘产生芝麻粒大小的灰黑色小斑点。9 月份进入发病后期,黑色病斑逐渐扩大,有时多个小斑扩大后融合在一起,呈不规则状。不同于褐斑病,该病斑能横跨叶脉,后期数个小斑融合在一起,发病严重时,叶片枯黄脱落(图1)。

图1 金银花黑斑病不同时期的危害症状Fig.1 Symptoms of the black spot disease in different periods on Lonicera japonica
2.2 病原菌的分离
对采集到的金银花叶片样品进行分离培养,分离纯化后得到的纯菌株在PDA 培养基上呈现出相同的菌落,经过ITS 序列比对,初步确定所分离得到的菌株属于同一种。挑选其中1 株(YP4)作为代表菌株进行后续形态学观察和系统发育学分析。
2.3 病原菌形态学鉴定
供试菌株YP4 在PDA 培养基上培养7 d 后,形成表面为灰白色或黄白色、棉絮状的菌落。菌落呈同心环状,中心为灰白色,外围白色发黄,并在PDA 培养基上产生棕黄色至深红色色素。经过番茄快速诱导产孢法和双玻片插片诱导产孢法诱导后,在显微镜下未观察到其分生孢子及分生孢子梗,仅观察到其菌丝。该菌株的菌丝光滑,直或弯曲,有分隔,具叉状分枝,直径约为1.8~2.5 μm(图2)。
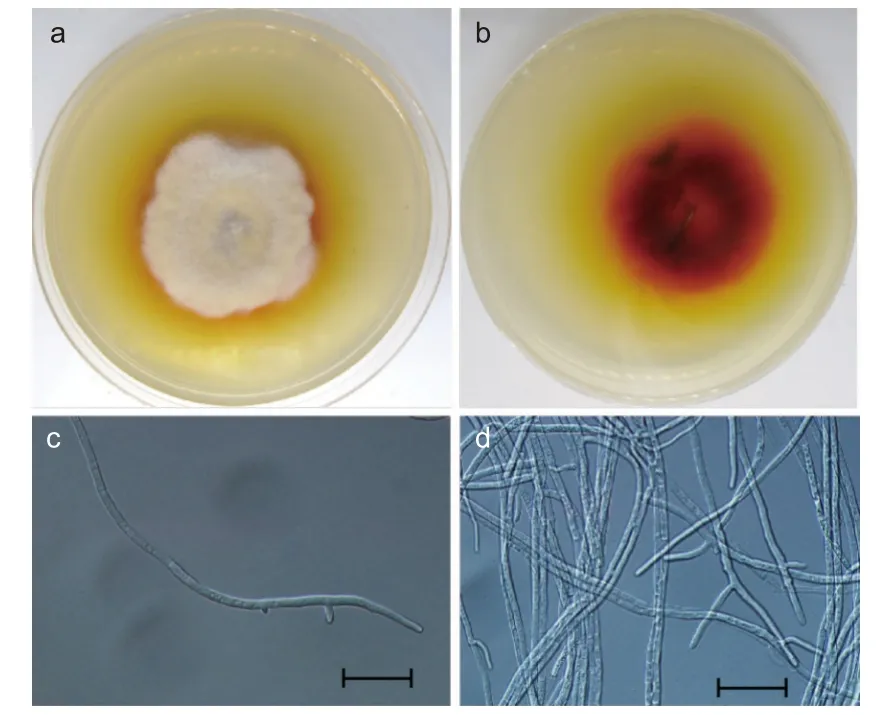
图2 菌株YP4 的形态特征Fig.2 Morphological characteristics of the strain YP4
2.4 病原菌分子鉴定
本研究基于ITS、gpd和EF-1α 3 个基因片段利用最大似然法(ML)、贝叶斯法(BI)和最大简约法(MP)分别构建了匍柄霉属的系统发育树(图3~5),3 个树中包含1 个供试菌株YP4 和36 个从Genbank 库中下载的参考菌株,以及1 个外类群菌株Alternaria alternata。ITS、gpd和EF-1α 3 个基因获得扩增片段大小分别为530、533和 674 bp(GenBank 登 录 号OQ157590、OQ186451 和OQ185385),串联后总长度为1 737 bp, 菌株YP4 与番茄匍柄霉Stemphylium lycopersici(EGS45-036、EGS45-043) 的参考序列进行多重比对时的一致性均为100%。并在3 种方法构建的系统发育树(图3~5)中,菌株YP4均与番茄匍柄霉S.lycopersici同源性最高,在ML树中以86% 的支持率聚为一支,在BI 树中以0.92 的支持率聚为一支,在MP 树中以85%的支持率聚为一支。

图3 基于最大似然法(ML) ITS、gpd 和EF-1α 联合基因系统发育树构建Fig.3 Maximum likelihood phylogenetic tree based on combined datasets of ITS,gpd and EF-1α
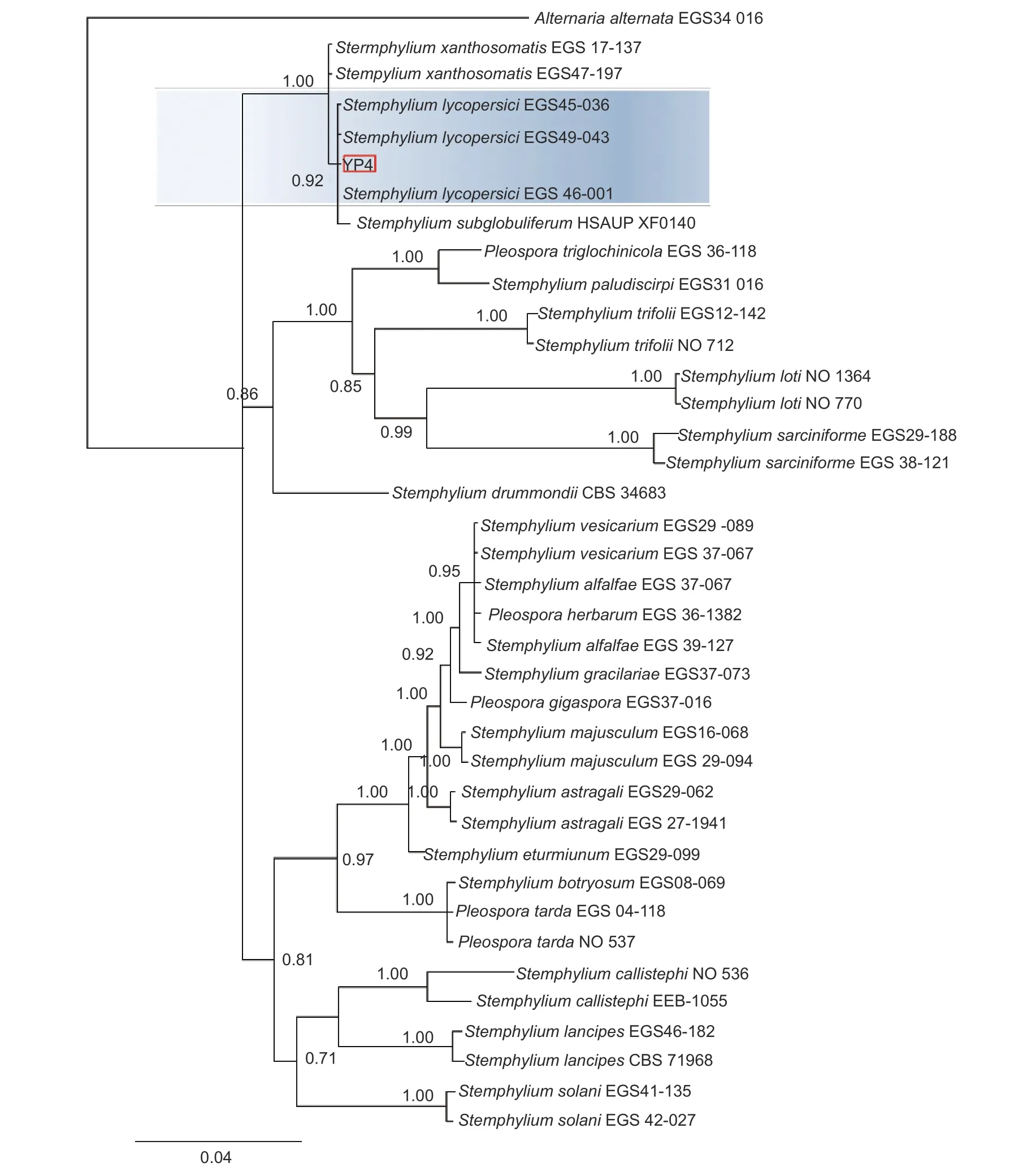
图4 基于贝叶斯法(BI)ITS、gpd 和EF-1α 联合基因系统发育树构建Fig.4 Bayesian phylogenetic tree based on combined datasets of ITS,gpd and EF-1α
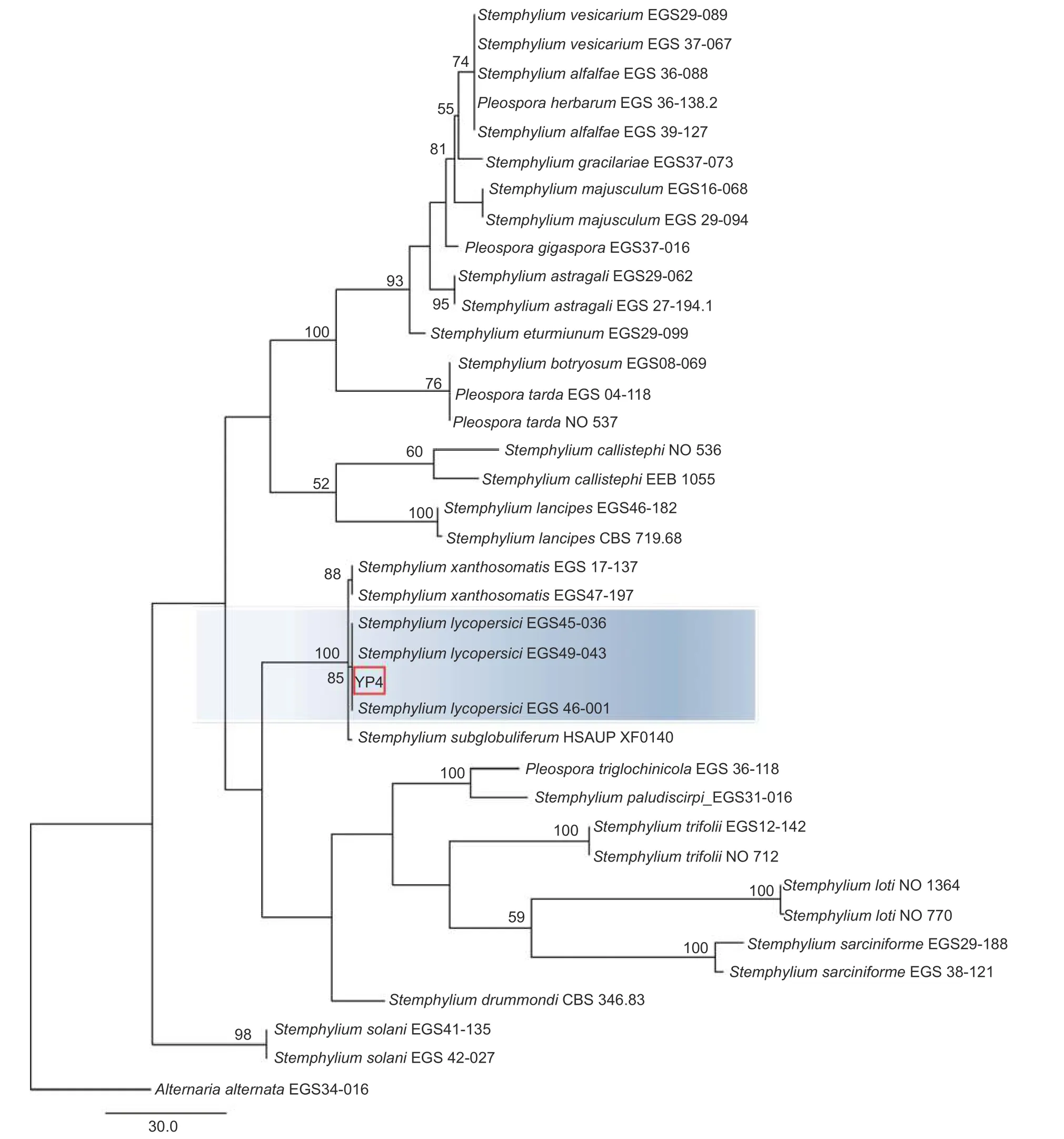
图5 基于最大简约法(MP)ITS、gpd 和EF-1α 联合基因系统发育树构建Fig.5 Phylogenetic tree (MP) based on combined datasets of ITS,gpd and EF-1α
2.5 致病性测定
在对金银花活体叶片接种9 d 后,叶片上产生病斑,呈不规则状,边界不清晰,横跨叶脉,并产生灰白色霉层,而对照组叶片无明显症状(图6)。根据柯赫氏法则,将接种叶片进行再分离,分离得到的菌株的形态特征与接种所用菌株相同。

图6 致病性接种实验金银花叶片症状Fig.6 Symptoms of leaves of Lonicera japonica after inoculation of the strain YP4
在对番茄离体叶片接种48 h 后,接种叶片上产生黑色病斑,呈不规则状,边界不清晰,横跨叶脉。而对照组叶片无明显症状(图7)。从表现症状的番茄离体叶片病斑上均能再次分离得到所接种的真菌菌株。

图7 致病性接种实验番茄叶片症状Fig.7 Symptoms of leaves of Solanum lycopersicum after inoculation of the strain YP4
3 讨论
本研究中所获得的菌株在系统发育学上与番茄匍柄霉聚为一支,在形态学上虽未能观察到该菌株的分生孢子和分生孢子梗,但其菌落形态与番茄匍柄霉相同,并且将该菌株接种到金银花叶片和番茄叶片上均可引起明显症状。因此,本研究结合形态学、系统发育学以及致病性分析,将引起河北省金银花黑斑病的病原菌鉴定为番茄匍柄霉(Stemphylium lycopersici(Enjoji) Yamamoto)。
由番茄匍柄霉引起的金银花黑斑病与金银花褐斑病相比有很大的不同:褐斑病的症状为发病初期在叶上形成褐色小点,后扩大成褐色圆斑或不规则病斑,病斑边缘受叶脉限制呈多棱状[17]。而这种黑斑病初期为芝麻状小黑斑,后期扩大为不规则状黑斑,并且能够横跨叶脉。
番茄匍柄霉隶属匍柄霉属(StemphyliumWallr.),该属是1883 年由Wallroth 以簇孢匍柄霉(S.botryosum)为模式种创立的。本属内大部分种类通过产生分生孢子进行无性繁殖,有一些种有性型为格孢腔菌属 (PleosporaRabenh.ex Ces.&De Not.),隶属于格孢腔菌目 (Pleosporales),格孢腔菌科 (Pleosporaceae)。匍柄霉属包括多种腐生真菌及病原真菌,其中病原真菌可危害包括多种农作物在内的寄主植物。匍柄霉属在形态上与其相近属链格孢属(AlternariaNees)相似,它们的分生孢子都是多隔且着色的,形成于由菌丝产生的分生孢子团上,而两属的区别在于匍柄霉属可在菌丝顶端的分生孢子团上产生连续的分生孢子[16,18-19]。匍柄霉属中的番茄匍柄霉也是一种世界性分布的重要植物病原菌,能够引起番茄、辣椒、茄子、莴苣和人参果等多种寄主植物产生叶斑病,于1931 年被Enjoji 从日本的番茄叶片上被首次分离出来[20-23]。有研究表明,该种匍柄霉通过合成和释放植物毒性次生代谢物破坏植物细胞,从而导致植物发病[24]。在中国,番茄匍柄霉于2001 年首次在山东省泰安市被发现,其寄主植物为番茄[18];随后,在我国多次发现番茄匍柄霉引起番茄产生灰叶斑病[25-27];此外,在国内番茄匍柄霉也可危害茄子、辣椒、柑橘、向日葵等经济作物的叶部[28-30],还有研究发现甜瓜根部病害也与番茄匍柄霉有关[31]。而本研究是番茄匍柄霉在我国危害金银花的首次报道,进一步拓宽了该病菌的寄主范围。
4 结论
本研究对采自河北省金银花样品进行了分离与纯化,结合形态学和多基因片段分析对其病原菌进行了分类鉴定,并依照科赫氏法则进行了致病性测定,发现了一种由番茄匍柄霉(Stemphylium lycopersici)引起的金银花新型叶斑病,并将该病害命名为金银花黑斑病,为金银花病害的科学防控提供了一定的理论依据。
