番茄素通过激活转录因子EB减轻脂多糖诱导的神经细胞损伤*
2024-01-17张卫钢毛嘉玥陈雨晴董明慧
张卫钢,汪 磊,毛嘉玥,张 杰,陈雨晴,董明慧,李 曙,王 林
(皖南医学院 基础医学院,安徽 芜湖 241002)
阿尔茨海默病(Alzheimer disease,AD)是常见的痴呆类型,其主要临床表现是学习和记忆等能力进行性丧失。AD 的典型病理性特征是老年斑(由淀粉样蛋白Aβ 沉积形成)和神经原纤维缠结(由过度磷酸化的τ 蛋白聚集形成)[1]。除此之外,神经炎症在AD 的发展进程中有着重要的作用[2]。在AD 患者脑组织中可观察到小胶质细胞过度激活、老年斑周围聚集大量促炎细胞因子,如白细胞介素1β(interleukin-1β,IL-1β)、肿瘤坏死因子α(tumor necrosis factor-α,TNF-α)等;同时AD患者的血浆中促炎细胞因子表达水平也升高[3]。AD病程的进展会随着炎症反应而加快,及时防治炎症可作为AD 的一种前瞻性治疗方法[4]。
异常聚集的Aβ和τ蛋白可促进神经炎症。自噬的主要功能是清除异常聚集的蛋白质[5];在AD 患者、AD 转基因鼠和细胞模型中均观察到自噬水平的降低可诱导炎症反应、Aβ和τ蛋白异常聚集;自噬途径和自噬相关蛋白在炎症的调节中起着核心作用[6]。转录因子EB(transcription factor EB,TFEB)在自噬进程中有着重要的作用,它可调节自噬体的形成、自噬体和溶酶体的融合以及溶酶体的生成和功能的正常发挥[7]。TFEB 在中枢神经系统中普遍表达,在神经元和胶质细胞中都有活性。在AD 患者的脑部中检测到细胞核中的TFEB 蛋白水平表达下降;在AD 模型鼠中,TFEB 激活后通过自噬溶酶体系统加速淀粉样前体蛋白(amyloid precursor protein,APP)的降解和减少Aβ 的产生[8]。TFEB 还可通过自噬溶酶体途径降解脑组织中的炎症小体和促炎细胞因子从而改善神经炎症。因此,TFEB 有望成为调控自噬和神经炎症的治疗靶标。
在不同AD 转基因鼠中,TFEB 的基因修饰已显示出神经保护作用,研究TFEB 激动剂在AD 中的作用将为新药开发提供依据[9]。番茄素(tomatidine,TA)是一种来源于茄科的甾体生物碱。番茄素通过增加抗氧化酶活性和减少细胞凋亡来消除氧化应激[10]。有研究报道番茄素可促进哺乳动物细胞或线虫中的自噬体的降解[11];N2a 细胞经氧糖剥夺/复氧损伤后,给予番茄素处理后TFEB 的活性增加,可促进溶酶体降解并减轻细胞损伤[12]。但是番茄素在脂多糖(lipopolysaccharide,LPS)诱导的神经炎症中的作用及机制尚未见报道。因此,本研究采用LPS 处理人神经母细胞瘤细胞(SH-SY5Y 细胞),模拟细胞水平的神经炎症模型,探究番茄素神经保护的可能机制。
材 料 和 方 法
1 细胞
SH-SY5Y 细胞购买于武汉普诺赛生命科技有限公司,货号为CL-0208。
2 主要试剂和仪器
LPS 和番茄素购自MedChemExpress;兔抗TFEB多克隆抗体购自武汉三鹰生物技术有限公司;兔抗Ser142 位点磷酸化TFEB(p-TFEB)多克隆抗体购自Affinity;兔抗P62、LC3 和cleaved caspase-3 多克隆抗体购自Cell Signaling Technology;兔抗β-actin 单克隆抗体、CCK-8 试剂盒等试剂均购自碧云天生物技术有限公司;Trizol、RNA 反转录试剂盒等均购自Vazyme;TNF-α 和IL-1β 引物购自生工生物工程(上海)股份有限公司;annexin V-FITC/PI凋亡试剂盒购自江苏凯基生物技术股份有限公司。SDS-PAGE 电泳仪及成像分析仪购自Bio-Rad (型号:ChemiDoc);流式分析仪购自BD(型号:Accuri);激光共聚焦显微镜购自Leica(型号为SP8)。
3 方法
3.1 细胞培养 SH-SY5Y 细胞复苏后用含有100 U/mL 青霉素、10 μg/mL 链霉素和10%胎牛血清的DMEM 培养液培养,置于37 ℃、5% CO₂培养箱中贴壁培养,待细胞增殖至融合度约80%左右时,用0.25%胰酶消化传代至另一培养瓶中,通过倒置光学显微镜观察细胞的生长增殖,待细胞状况稳定后用于后续实验。
3.2 细胞分组及给药 先观察不同浓度(1.25、2.5、5 和10 μg/mL)的LPS 对细胞活力和TFEB 活性的影响,并确定本研究的LPS 浓度为5 μg/mL。再观察不同浓度(1、5 和10 μmol/L)的番茄素对细胞活力和TFEB 活性的影响,结合相关文献确定用于本研究的番茄素浓度为5 μmol/L[13]。将SH-SY5Y 细胞分为对照(control,CON)组、LPS 组和LPS+TA 组。参考相关文献在LPS 组中加入5 μg/mL LPS 处理24 h,建立炎症模型[14],LPS+TA 组先加入5 μmol/L 番茄素处理24 h后再加入5 μg/mL LPS共培养24 h。
3.3 CCK-8 法检测细胞活力 将SH-SY5Y 细胞悬浮液以适当密度种入96 孔板,同时设置空白对照组和正常对照组,细胞贴壁后进行相应处理,LPS 处理24 h,番茄素处理48 h 后,每孔加入10 μL CCK-8 试剂,培养箱中孵育1 h,用酶标仪测量450 nm 波长下不同组别的吸光度(A)并算出细胞活力。
3.4 流式细胞术检测细胞凋亡 按凋亡试剂盒说明书,根据实验需求,将SH-SY5Y细胞处理完成后用不含EDTA的胰酶消化,停止消化后用PBS温和清洗细胞2 次,离心5 min,收集1×105~5×105个细胞,加入500 μL 试剂盒内缓冲液轻轻吹打混匀成单细胞悬液,再依序加入annexin V-FITC 试剂和PI 试剂各5 μL,吹打混匀,室温、避光反应5~10 min,随后在流式细胞仪上检测细胞凋亡率,时间控制在1 h内。
3.5 Western blot 检测蛋白水平 根据不同实验需求,细胞处理完成后,去掉培养上清液,PBS 温和清洗细胞2次,加入RIPA裂解液和蛋白酶抑制剂,静置20 min,使细胞充分裂解,收集细胞至EP 管,4 ℃、10 000×g离心20 min,弃沉淀留上清液,用BCA 法测定蛋白质浓度。根据不同目的蛋白分子量配制10%或12%聚丙烯酰胺凝胶,上样蛋白量为30 μg,电泳分离蛋白质后,采用湿式转移法转移到NC 膜。5%脱脂奶粉溶液封闭1 h,分别加入1∶1 000 稀释的兔抗TFEB、p-TFEB、P62、LC3 和β-actin 单克隆抗体,4 ℃孵育过夜,次日TBST清洗3次,每次10 min,清洗后加入辣根过氧化物酶标记的羊抗兔IgG 或羊抗鼠IgG(1∶1 000),室温摇床孵育1 h,再用TBST 清洗3次,每次10 min。洗膜后用ECL 化学发光液和凝胶成像分析仪曝光条带,ImageJ 软件定量分析蛋白灰度值,目的蛋白与内参照β-actin 之比代表蛋白相对表达量。
3.6 RT-qPCR 法检测mRNA 表达水平 使用Trizol试剂提取预处理后的细胞总RNA,逆转录成cDNA。以cDNA 为转录模板,每孔取2 μL 加入八联管中,再加上、下游引物(序列见表1)各0.4 μL,SYBR qPCR Master Mix 10 μL,DEPC 水7.2 μL。反应条件为:95 ℃预变性30 s;95 ℃变性10 s,60 ℃退火30 s,40个循环。根据产物的熔解曲线得到Ct 值,计算炎症因子mRNA的相对表达水平。

表1 RT-qPCR引物序列Table 1. The sequences of the primers for RT-qPCR
3.7 免疫荧光检测 各组细胞干预完成后,吸出培养基,沿着孔壁缓慢注入PBS 清洗,加入4%多聚甲醛,室温固定30 min,1% Triton-PBS清洗3次;免疫染色通透液破膜30 min,1% Triton-PBS清洗3次;5%牛血清白蛋白室温封闭1 h,加入免疫染色稀释液配制的Ⅰ抗(TFEB抗体,1∶200;cleaved caspase-3抗体,1∶250),4 ℃孵育过夜;1% Triton-PBS 清洗3 次,清洗后用3%牛血清白蛋白配制的Ⅱ抗(1∶500)室温避光孵育1 h,1% Triton-PBS 清洗后加入DAPI 染色液37 ℃避光孵育30 min,PBS清洗后滴加适量抗荧光淬灭封片液于载玻片,采用激光共聚焦显微镜拍照。
4 统计学处理
使用SPSS 17.0 统计软件处理数据。数据用均数±标准差(mean±SD)表示。多组间比较采用单因素方差分析(one-way ANOVA),组间两两比较采用最小显性差异法(LSD 法)。以P<0.05 为差异有统计学意义。
结 果
1 LPS对细胞炎症反应和凋亡的影响
不同浓度的LPS(1.25、2.5、5 和10 μg/mL)处理细胞24 h。CCK-8 实验结果表明,与CON 组相比,5 μg/mL 和10 μg/mL 的LPS 使细胞活力显著下降(P<0.05),见图1A。分别采用2.5 和5 μg/mL 的LPS 孵育细胞24 h,RT-qPCR 结果显示,与CON 组相比,IL-1β mRNA 水平在LPS 浓度为2.5 和5 μg/mL 组中均显著上升(P<0.05),TNF-α mRNA 水平只在LPS 浓度为5 μg/mL 组中表达显著上升(P<0.05),见图1B。流式细胞仪检测结果显示,2.5 和5 μg/mL 的LPS 引起细胞凋亡的数量显著增加(P<0.01),见图1C。免疫荧光染色检测结果表明,与CON 组相比较,cleaved caspase-3 表达水平在2.5 和5 μg/mL LPS 组中显著上升(P<0.05),见图1D。

Figure 1. Effects of LPS on the inflammatory response and apoptosis of cells. A: the CCK-8 assay was used to measure the viability of SH-SY5Y cells after LPS treatment; B: RT-qPCR was conducted to detect TNF-α and IL-1β mRNA levels in SH-SY5Y cells after LPS treatment; C: flow cytometry was performed to measure the apoptosis of SH-SY5Y cells after LPS treatment;D: representative cleaved caspase-3 fluorescence staining images of SH-SY5Y cells in each group (green represents cleaved caspase-3,and blue represents nuclei stained with DAPI; scale bar=50 μm). Mean±SD. n=3. *P<0.05,**P<0.01 vs CON group.图1 LPS对SH-SY5Y细胞炎症反应和凋亡的影响
2 LPS对细胞自噬和TFEB活性的影响
Western blot 实验结果显示,与CON 组相比,5 μg/mL LPS 处理组P62、LC3-II/LC3-I 和TFEB 表达水平均显著下降(P<0.05),但是p-TFEB 表达水平显著上升(P<0.01);2.5 μg/mL LPS 虽引起P62、LC3-II/I和TFEB 表达水平下降但无统计学意义,见图2A。免疫荧光染色结果显示,TFEB 在5 μg/mL LPS 处理组只定位在胞质中(图2B)。综合以上实验结果,本研究选择5 μg/mL的LPS浓度作为后续实验条件。
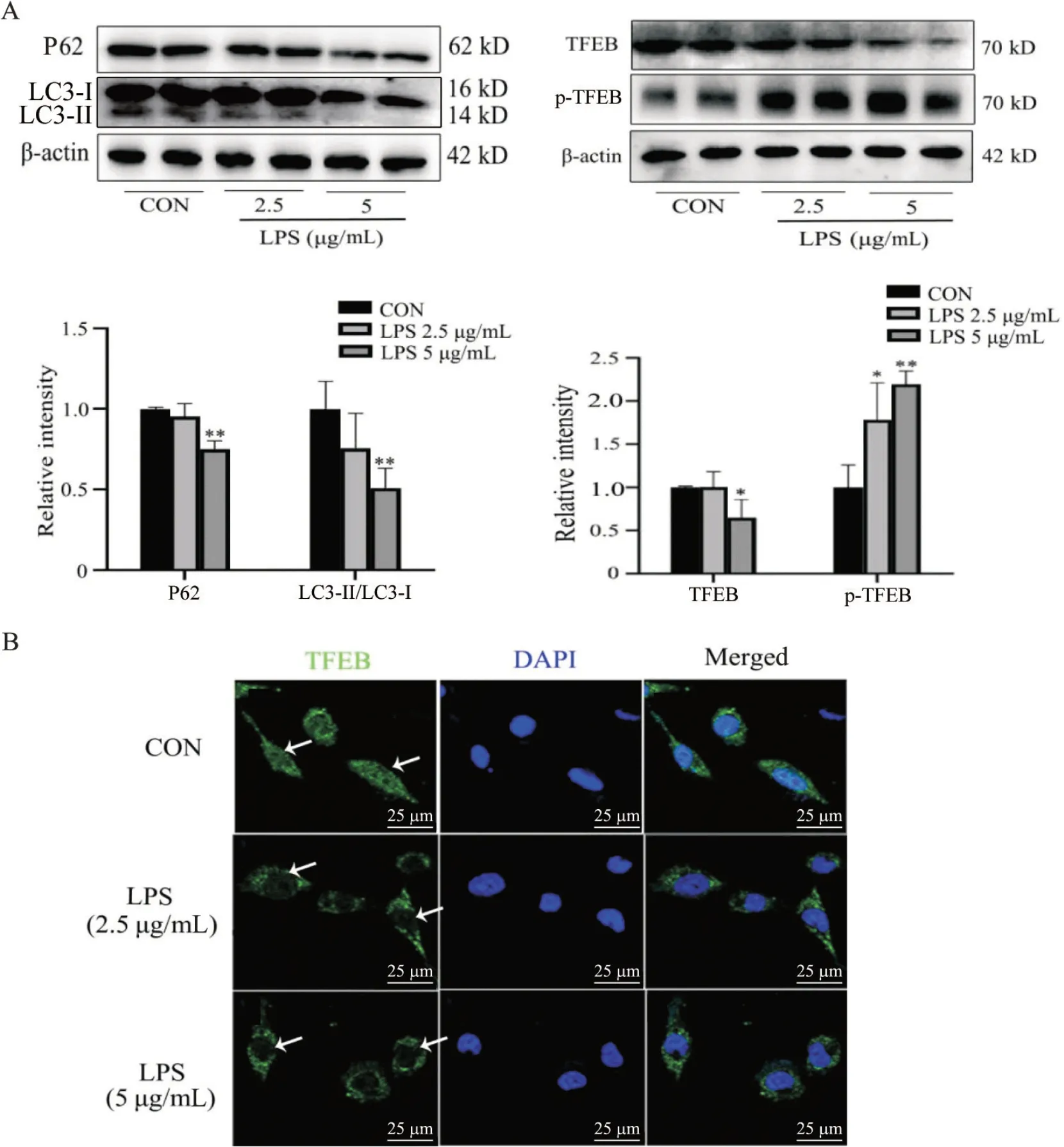
Figure 2. Effects of LPS on the autophagy and TFEB activity of SH-SY5Y cells. A: Western blot was performed to detect P62,LC3,TFEB and p-TFEB protein levels; B: representative TFEB fluorescence staining images of SH-SY5Y cells in each group(green represents TFEB,and blue represents nuclei stained with DAPI; scale bar=25 μm). Mean±SD. n=3. *P<0.05,**P<0.01 vs CON group.图2 LPS对SH-SY5Y细胞自噬和TFEB活性的影响
3 番茄素对TFEB的表达和定位的影响
分别用1、5 和10 μmol/L 的番茄素处理细胞48 h。CCK-8 结果表明,1、5 和10 μmol/L 的番茄素对细胞的活力无显著影响(图3A)。Western blot 实验结果显示,与CON 组相比,5 μmol/L 番茄素处理组,TFEB 表达水平显著上升(P<0.01),但p-TFEB 的表达水平显著下降(P<0.05),见图3B。免疫荧光染色结果显示,与CON 组相比,5 μmol/L 番茄素处理后TFEB主要定位于细胞核中(图3C)。

Figure 3. Effect of tomatidine (TA) on the expression and localization of TFEB. A: the CCK-8 assay was used to measure the viability of SH-SY5Y cells after tomatidine treatment; B: Western blot was used to detect TFEB and p-TFEB protein levels; C:representative TFEB fluorescence staining images of SH-SY5Y cells in each group (green represents TFEB,and blue represents nuclei stained with DAPI; scale bar=25 μm). Mean±SD. n=3. *P<0.05,**P<0.01 vs CON group.图3 番茄素对TFEB的表达和定位的影响
4 番茄素对LPS 处理的细胞TFEB 和自噬功能的影响
Western blot 实验结果表明,与CON 组比,LPS 组中P62 表达水平下降但差异无统计学意义(P=0.19),LC3-II/LC3-I 和TFEB 表达水平显著下降(P<0.05),而p-TFEB 表达水平显著上升(P<0.05);与LPS 组相比,LPS+TA 组中P62 有下降趋势但差异无统计学意义(P=0.08),p-TFEB 表达水平显著下降(P<0.01),而LC3-II/LC3-I 和TFEB 表达水平显著上升(P<0.05),见图4A。免疫荧光染色结果显示,与CON 组相比,TFEB 在LPS 组中主要定位于胞质,而在LPS+TA组中则易位到细胞核中,见图4B。

Figure 4. Effects of tomatidine(TA) on TFEB and autophagy in LPS-treated SH-SY5Y cells. A: Western blot was used to detect P62,LC3,TFEB and p-TFEB protein levels; B: representative TFEB fluorescence staining images of SH-SY5Y cells in each group (green represents TFEB,and blue represents nuclei stained with DAPI; scale bar=50 μm). Mean±SD. n=3.*P<0.05 vs CON group; #P<0.05,##P<0.01 vs LPS group.图4 番茄素对LPS处理的SH-SY5Y细胞TFEB表达和自噬功能的影响
5 番茄素对LPS 处理的细胞炎症反应和细胞凋亡的影响
RT-qPCR 结果表明,与CON 组相比,IL-1β 和TNF-α mRNA 表达水平在LPS 组中显著上升(P<0.01);与LPS 组相比,LPS+TA 组中IL-1β mRNA 表达有下降趋势但差异无统计学意义(P=0.16),TNFα mRNA 表达显著下降(P<0.01),见图5A。流式细胞仪检测结果显示,与CON 组相比,LPS 组细胞凋亡率显著上升(P<0.01);与LPS 组相比,LPS+TA 组细胞凋亡率显著下降(P<0.01),见图5B。免疫荧光染色结果显示,与CON组相比,LPS组cleaved caspase-3表达显著上升(P<0.01);与LPS 组相比,LPS+TA 组cleaved caspase-3表达显著下降(P<0.01),见图5C。

Figure 5. Effects of tomatidine (TA) on inflammatory response and apoptosis in LPS-treated SH-SY5Y cells. A: RT-qPCR was used to detect TNF-α and IL-1β mRNA levels in SH-SY5Y cells; B: flow cytometry was used to measure the apoptosis of SHSY5Y cells; C: representative cleaved caspase-3 fluorescence staining images of SH-SY5Y cells in each group (green represents cleaved caspase-3,and blue represents nuclei stained with DAPI; scale bar=50 μm). Mean±SD. n=3. **P<0.01 vs CON group; ##P<0.01 vs LPS group.图5 番茄素对LPS处理的SH-SY5Y细胞炎症反应和细胞凋亡的影响
讨 论
神经炎症可通过不同的方式诱导,其中LPS 诱导是常见的。LPS存在于革兰氏阴性菌外膜中,其主要受体是Toll 样受体,从而促进多种炎症介质的表达和神经退行性疾病的发展。本研究同样利用LPS处理SH-SY5Y 细胞模拟细胞神经炎症的微环境,探讨在细胞水平上番茄素能否发挥神经保护作用。与以往实验结果一致,本研究结果也显示LPS 可引起炎症因子IL-1β、TNF-α mRNA 表达水平和促凋亡蛋白cleaved caspase-3 的表达水平显著上升,凋亡细胞的数量也显著增多。番茄素主要存在于未成熟的绿色番茄的皮中,已被证明是耐药肿瘤细胞的化学增敏剂,可抑制肺癌、肝癌、结肠癌细胞的增殖和侵袭;番茄素还有抗炎、抗氧化、抑制病毒传播和保护心脏等功能[15]。有研究报道,在谷氨酸、双氧水处理的SH-SY5Y 细胞中,番茄素具有一定的神经保护作用[16],但番茄素能否在神经炎症中发挥保护作用尚未见报道。本研究结果显示给予番茄素处理后,SHSY5Y 细胞炎症因子的表达水平显著降低,凋亡细胞数量显著减少;该结果初步证明在LPS 处理的SHSY5Y 细胞中,番茄素具有一定神经保护作用,但其具体机制尚不明确。
TFEB 包含基本的螺旋-环-螺旋-亮氨酸拉链结构域(bHLHZip),通过与靶基因E-box (CANNTG)启动子元件紧密结合来调控靶基因的表达,从而参与溶酶体生物发生、自噬调节和应激适应等细胞过程[17]。在TFEB敲除小鼠脑组织中观察到Aβ沉积和磷酸化τ 蛋白的积累;在AD 小鼠模型中,TFEB 的过表达增强了对细胞脂滴和受损线粒体的降解[18]。但目前关于TFEB 在神经炎症中的作用尚无报道,本研究结果显示,在SH-SY5Y 细胞中,LPS 诱导TFEB 的表达水平显著降低,该结果表明细胞的神经炎症可降低TFEB 的表达。在缺血缺氧损伤的小鼠神经母细胞瘤N2A 细胞中,番茄素促进TFEB 入核和TFEB表达水平增加。与其结果一致,本研究中给予番茄素处理后,TFEB 的表达水平显著增加。生理情况下TFEB 主要位于胞质中,当饥饿或受刺激时TFEB 快速转移到细胞核并增强其有关靶基因的转录和靶蛋白的表达水平[19]。翻译后修饰调控TFEB 的亚细胞定位,磷酸化是调控TFEB 活性的中心驱动因素,去磷酸化后TFEB 从胞质易位至胞核[20]。本研究中,SH-SY5Y 细胞给予LPS 处理后p-TFEB 表达水平显著上升,TFEB 主要定位于胞质中;而番茄素处理后,p-TFEB 表达水平显著降低,TFEB 从胞质易位至胞核增强自噬功能。本实验结果证明在LPS 处理的SH-SY5Y 细胞中,番茄素可促进TFEB 的表达和核移位。
TFEB 可参与自噬体形成和自噬溶酶体降解,TFEB 表达水平降低虽引起自噬功能障碍[21],但是要明确具体哪一过程异常须结合自噬其他指标的变化来判定。LC3 是目前认可度最高的自噬标志物,贯穿于自噬体从形成到成熟的各个阶段;自噬初始阶段受到抑制,可降低自噬体的形成和LC3-II 分子的表达水平[22]。本研究的结果显示,给予LPS 处理后,LC3-II/LC3-I 表达水平显著下降,初步证实LPS 抑制自噬体的形成。自噬是一个动态的过程,单检测LC3-II/LC3-I 的表达水平无法完全充分反映细胞内的自噬状态。P62 是另一个常用的自噬标志物,P62和LC3-II/LC3-I 表达水平的变化可反映自噬通量的改变。在自噬过程中,P62与泛素化的蛋白质结合后与LC3-Ⅱ相互作用,靶向自噬体最终在自噬溶酶体内降解;当自噬体形成受抑制或溶酶体功能受损时,P62 会在胞质中逐渐累积[23]。P62 的活性受到磷酸化、乙酰化等修饰的调节[24]。在AD 患者脑中检测出P62蛋白水平的低表达,敲除P62抑制自噬功能会导致Aβ 积累、τ 蛋白过度磷酸化和神经退行性变[25]。本研究中初步证实LPS 抑制自噬体的形成,但实验结果却显示LPS处理后的P62的表达水平显著下降。已有大量文献报道LPS 会导致腺苷酸活化蛋白激酶(AMP-activated protein kinase,AMPK)的活性下降,而AMPK 可通过磷酸化水平调控P62 的表达[26]。本课题组其他实验结果也证实,在细胞水平上LPS 降低AMPK 的活性;因此,我们猜测在LPS 处理SHSY5Y 细胞中,P62 的表达水平显著下降是可能由于AMPK 活性降低引起,具体机制需要进一步验证。在LPS 处理的SH-SY5Y 细胞中,给予番茄素处理后LC3-II/LC3-I 表达水平显著上升,而P62 表达水平虽降低但差异无统计学意义,该实验结果初步表明在SH-SY5Y细胞中番茄素可促进自噬体的形成。
在AD 发生发展过程中,自噬、神经炎症和蛋白质异常聚集相互影响[27]。有研究报道AD 患者颞叶皮质中自噬蛋白P62和LC3阳性囊泡增加,与炎症小体、Aβ和过度磷酸化τ蛋白共定位,这可能是因为炎症小体的产生促进神经炎症,从而降低自噬功能,导致AD 患者颞叶处蛋白异常聚集[28]。自噬功能的增强可促进炎症因子的降解,抑制神经炎症和发挥神经保护的作用。在本研究中,LPS引起炎症因子表达水平增加,自噬体形成受到抑制;给予番茄素处理后增强自噬体的形成,促进炎症因子的降解。番茄素促进炎症因子的降解能否改善神经炎症引起细胞凋亡,本研究中检测细胞凋亡蛋白cleaved caspase-3 的表达水平和细胞凋亡数量的变化,实验结果显示番茄素处理后cleaved caspase-3 表达水平和凋亡细胞数量均减少。以上结果可表明番茄素可增强自噬功能,促进炎症因子的降解,从而减少SH-SY5Y 细胞凋亡。
综上所述,我们的研究结果显示,在细胞水平上,番茄素增强TFEB 表达并促进其入核,改善LPS引起的细胞自噬功能障碍和凋亡,从而发挥神经保护作用。但是本研究结果仅在细胞水平上有验证,该机制是否在动物水平有效仍需进一步研究。
