不同时长的睡眠剥夺对注意警觉功能的影响
2023-12-27王家涛周前祥
殷 瑛,王家涛,周前祥
(北京航空航天大学生物与医学工程学院,北京 100191)
1 引言
警觉度指标在航天、交通安全领域和军事行动领域均发挥重要作用,而警觉度的降低可能导致任务绩效下降,甚至危及人员安全[1-2]。随着载人航天技术的不断发展,航天员在轨工作的时间越来越长,维持警觉状态对于个体保证工作绩效十分重要[3-4]。在轨人员睡眠不足会影响认知表现[5-6],从而引发安全事故[7]。因此,探讨睡眠不足对认知功能影响的机制具有重要的现实意义。完全急性睡眠剥夺(Total Sleep Deprivation,TSD)作为操纵睡眠不足状态的重要范式,被广泛用于探索睡眠不足情况下,认知行为和神经功能变化的重要理论问题。睡眠剥夺期间参与者需要一直保持清醒,时长从24 h 到72 h 不等[8]。
警觉性注意(Vigilant Attention)反映了一定时间内将注意力维持在简单反应任务上的能力[9-10]。作为一种基础认知功能,警觉性注意的降低会影响执行功能的表现[11-12]。在睡眠剥夺影响注意的研究中,精神运动警觉性测试(Psychomotor Vigilance Test,PVT)因其灵敏性高,个体差异较小成为测量警觉性注意的首选范式[10]。
Mao 等[11]发现睡眠剥夺会严重影响多种认知过程,包括警觉性注意、工作记忆、执行功能[13-14]。这些发现支持了警惕注意力在睡眠不足后维持整体神经行为功能的关键作用。一晚的睡眠剥夺后个体完成go/no-go 任务和PVT 任务的行为表现显著下降,且go/no-go 任务中增加的脱漏次数与PVT 任务中下降的反应时存在相关关系。睡眠不足后警觉注意力的损害部分解释了睡眠剥夺对抑制控制的影响,并在一定程度上证实了警觉假说。警觉性注意对睡眠剥夺尤为敏感,具体表现在随睡眠剥夺时间增加,PVT 任务的反应时和正确率都显著下降,而受昼夜节律影响,在深夜和凌晨警觉性注意受损最严重[10]。在神经层面,睡眠剥夺后认知功能受损及大范围α功率激活和功能连接减弱有关,其中默认网络、顶枕网络、感觉运动网络和丘脑是影响认知功能表现的重要区域[15-17]。但睡眠剥夺对认知功能影响的时间累积效应并不一致。Skurvydas 等[18]研究发现,24 h 的睡眠剥夺不影响执行功能的表现,而在48 h 后表现显著下降。同样,30 h 睡眠剥夺后的工作记忆表现显著低于24 h[19]。Stenson 等[20]发现参与者在39 h 的完全睡眠剥夺期间,警觉性注意的任务表现呈波动下降。
Lowe 等[21]分析表明,睡眠剥夺对跨认知领域的认知处理有显著的负面影响,包括警觉性、简单注意力、持续注意力。由于警觉注意力对于高级认知功能至关重要,并且最容易受到睡眠剥夺的影响,因此研究人员提出,警觉注意力可能是解释睡眠剥夺后其他认知缺陷变化的基本因素[22]。从认知加工的角度分析,行为决策等认知过程是以个体对相关信息的前注意加工为基础的。个体之所以在睡眠缺失后表现出操作失误的行为,可能在于他们在做出认知决策之前,选择性地注意了无关信息。尽管这一假设已经存在多年,但很少有研究明确地指出警觉注意力在整个睡眠剥夺中是如何变化的。目前还不清楚警觉注意力的受损随着剥夺时间的增加线性下降是否动态变化。
本文以事件相关电位技术构建不同时长睡眠剥夺模型,明确不同时长的睡眠剥夺对警觉注意加工的损害特征。研究结果可为科学管控睡眠、科学应对失眠等睡眠障碍提供参考。
2 方法
2.1 志愿者
实验共招募30 名志愿者,年龄22.79±2.27岁。志愿者均为右利手,视力或矫正视力正常,之前未参加过心理生理相关实验,无精神神经疾病史,无焦虑和抑郁症状,近期无急性感染史或感染症状,近期无药物治疗史,体内无植入金属。经数据筛查后,保留27 名志愿者的数据。志愿者智商处于人群平均水平以上(IQ>110 分)[23]。使用匹兹堡睡眠质量指数问卷测验(Pittsburgh Sleep Quality Index,PSQI)对志愿者睡眠情况进行筛查(PSQI<5)。在实验前所有志愿者详细了解实验流程及试验中注意事项,并填写了知情同意书。实验经过了北京航空航天大学伦理委员会的批准。
2.2 实验流程
研究采用单因素组内设计。志愿者在测试前一天到达实验室,确保当晚在实验室有至少8 h的睡眠。第二天8:00 睡眠剥夺开始,志愿者完成了第一次3 min 的闭眼静息态数据采集和PVT 任务脑电数据的采集,作为基线测量(Baseline)。第三天8:00(记作24 h-TSD 值),志愿者完成了第二次静息态脑电数据采集和PVT 任务脑电数据。第三天20:00(记作36 h-TSD 值),志愿者完成了第三次静息态脑电数据采集和PVT 任务脑电数据,然后整个实验结束。每次实验同时有2名志愿者参与,整个睡眠剥夺过程中有2 名医护人员和一名主试人员陪同监督。实验流程如图1所示。
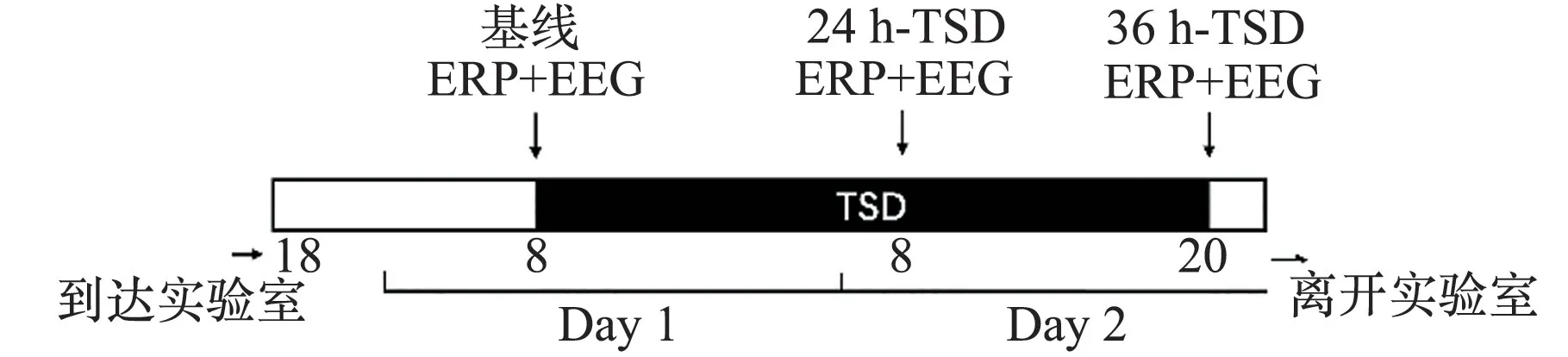
图1 实验流程示意图Fig.1 Flow chart of the experiment
2.3 检测方法
2.3.1 精神运动警觉性任务
在睁眼静息状态期间,志愿者目光集中到屏幕中央固定的十字上。在PVT 任务中,开始时屏幕出现一个固定在中央的十字,呈现时间为随机2~10 s 的间隔时间随后转换到一个红色圆点,志愿者被要求用右手食指尽快按空格键,按键后屏幕显示反应时间,马上再次转换到十字显示。任务示意图见图2。

图2 PVT 任务示意图Fig.2 Schematic diagram of PVT task
2.3.2 脑电记录
验程序采用E-prime 2.0 编写和呈现。EEG信号则通过64 通道电极帽(10-20 系统标准位置)结合SynAmps2 放大器(Compumedics Neuroscan,Charlotte,NC,USA)通过Neuroscan 在线采集。采集过程中,采样率为1000 Hz,电极阻抗保持在5 kΩ 以下,双侧乳突作为在线参考电极[23]。
2.4 数据处理
2.4.1 ERP 分析
ERP (Event Related Potential) 数 据 采 用SCAN4.3 软件进行预处理和统计参数分析。经脑电预览后,采用回归分析去除眼电伪迹,经滤波(带通为0.05~17 Hz)、脑电分段(ERP 分析窗选用900 ms,以刺激前100 ms 用于基线校正)、基线校正后,对所有正确反应的脑电诱发电位进行叠加平均。
选取F3、Fz、F4、C3、Cz、C4、P3、Pz、P4 9 个电极点,提取P2、P3 成分的波幅和潜伏期[23]。选取刺激呈现后150 ~250 ms 作为提取P2 成分的时间窗,选取刺激呈现后250 ~400 ms 作为提取P3 成分的时间窗。计算9 个点的平均波幅和潜伏期纳入到后续统计处理中。
2.4.2 EEG 频谱分析
脑电相对功率的计算结果可以用于分析脑电信号在不同频段内的能量分布情况,本文用此来探究特定频段内的脑活动对于注意加工过程的贡献。
使用MATLAB 中的EEGLAB 工具包进行预处理和相对功率计算。考虑到在实验刚开始时被试可能还没有进入状态,因此删除前30 s 的数据。为了提高数据信噪比,将剩余的脑电数据分为2 s 一段进行分析。鉴于脑电信号是一种非平稳信号,为避免在每段边缘处出现信号衰减和畸变,每段数据均加上一个等长的汉明窗(Hamming window)函数(重叠率为50%),以进行基于Welch算法的功率谱估计。对分段后的EEG 数据进行傅里叶变换。对α(8~14 Hz)频段范围内的EEG功率谱进行平均,得到该条件下该频段的平均绝对功率。将该频段的绝对功率除以整个目标频率范围的总功率,计算α 频段的相对功率,以消除两种条件下功率基线不同的情况。
2.4.3 统计学处理
3 结果
3.1 行为学结果
对于每个志愿者在睡眠剥夺前后完成PVT任务的平均反应时进行统计分析(表1)。结果显示,时间主效应显著(P<0.001)。事后比较结果表明,与基线相比,24 h 完全睡眠剥夺后,被试完成任务的平均反应时显著延长(P<0.001),36 h完全睡眠剥夺后,被试完成任务的平均反应时显著延长(P<0.001)。24 h 完全睡眠剥夺后的反应时显著大于36 h 完全睡眠剥夺后(P=0.007)。
表1 不同时长睡眠剥夺下被试的任务反应时()Table 1 Reaction time to task in subjects with different lengths of sleep deprivation()

表1 不同时长睡眠剥夺下被试的任务反应时()Table 1 Reaction time to task in subjects with different lengths of sleep deprivation()
注:***P<0.001 vs 基线;#P<0.01 vs 24 h-TSD
参数 基线 24h-TSD 36h-TSD反应时/ms 313.137±36.25 355.96±8.33***337.03±2.036***#
3.2 ERP 结果
3.2.1 波幅
对3 个时间点的P2、P3 成分波幅分别进行单因素重复测量,结果表明,时间主效应对P2 波幅(P=0.010)和P3 波幅(P=0.006)影响显著。即不同睡眠剥夺时长下,P2 和P3 成分的波幅存在显著差异。
事后比较结果表明,与基线相比,24 h 完全睡眠剥夺后,志愿者的P2 波波幅和P3 波波幅均显著下降(P=0.012;P=0.002),36 h 完全睡眠剥夺后,志愿者的P2 波波幅和P3 波波幅均显著下降(P=0.040;P=0.037)。24 h 完全睡眠剥夺后的P2 波幅和P3 波幅与36 h 完全睡眠剥夺后无显著差异。不同时长睡眠剥夺下脑电成分的波幅和潜伏期见表2,不同时长睡眠剥夺下被试完成警觉任务的脑电波形图见图3。
表2 不同时长睡眠剥夺下脑电成分的波幅和潜伏期(,n=27)Table 2 Amplitudes and latencies of electroencephalographic components under different durations of sleep deprivation(,n=27)

表2 不同时长睡眠剥夺下脑电成分的波幅和潜伏期(,n=27)Table 2 Amplitudes and latencies of electroencephalographic components under different durations of sleep deprivation(,n=27)
注:#P<0.05 vs 基线;##P<0.01 vs 基线
参数基线24h-TSD 36h-TSD P2 P3 P2 P3 P2 P3波幅 6.30±3.98 8.34±5.35 4.21±2.83# 5.90±3.70## 4.72±2.27# 6.59±4.98#潜伏期 222.72±32.58 424.05±31.49 225.36±36.67 436.32±38.65 218.47±26.64 439.15±27.13

图3 不同时长睡眠剥夺下志愿者完成警觉任务的脑电波形图Fig.3 ERP waveforms under different durations of sleep deprivation
3.2.2 潜伏期
对3 个时间点的P2、P3 成分潜伏期分别进行单因素重复测量,结果表明,时间主效应对P2潜伏期(P>0.05)没有显著影响。时间主效应对P3 潜伏期(P=0.058) 有显著影响。事后比较结果表明,与基线相比,24 h 完全睡眠剥夺后,P3 潜伏期显著延长(P=0.026)。
3.3 EEG 频谱分析结果
对3 个时间点的α 频段的相对功率进行单因素重复测量,结果表明时间主效应对α 频段相对功率值有显著影响(P=0.022)。相较于基线和24 h 完全睡眠剥夺,36 h 睡眠剥夺后α 频段相对功率显著降低(P=0.014)。不同时长睡眠剥夺下被试静息态α 频段相对功率见表3、图4。

表3 不同时长睡眠剥夺下α 频段相对功率值Table 3 Relative power values in the alpha band for different durations of sleep deprivation

图4 不同时长睡眠剥夺下被试静息态α 频段相对功率Fig.4 Relative power in the resting alpha band under different durations of sleep deprivation
4 讨论
本文比较了不同睡眠剥夺时长下个体的行为学指标、脑电成分以及脑电频谱活动。研究发现,睡眠剥夺后,受试者任务反应时显著下降,睡眠剥夺影响了个体的警惕性、动作准备和执行[24]。24 h 睡眠剥夺后,个体的反应时和脑电P2、P3 成分的波幅呈现出断崖式下降,相较于基线状态,大幅度降低。而与后续36 h 睡眠剥夺的结果相比,个体的注意警觉水平并未呈现出显著差异。这说明警觉注意力并非会随着剥夺时间的增加而线性下降,睡眠剥夺24 h 后,个体的认知能力进入一个平台期。经过一定时间的睡眠剥夺后,个体可能已经适应了这种状态,或者在某种程度上恢复了一些认知功能。这种适应可能表现为生理上的调整,例如改变神经化学物质的水平,以帮助维持警觉性。此外,心理适应也可能发挥作用,个体采取更积极的策略来应对疲劳和困倦,从而保持注意警觉水平。
α 频段的功率是用来反映警觉能力的良好脑电指标[25],该频段的静息态活动在闭眼过程中进行采集信号更好[26]。此外,α 频段和抑制控制、注意力等功能密切相关[27-28]。Boonstra 等[29]研究表明,24 hTSD 额叶皮质的α 活性降低,高α 活性可能表明抑制任务无关的大脑区域或总体大脑活动的降低。本文结果表明,36 h 睡眠剥夺后大脑的自发α 活动显著降低,而基线与24 h 睡眠剥夺后的α 频段活动无显著差异。结合行为学和ERP 分析的结果,24 h 睡眠剥夺后,个体认知能力急速下降,大脑采取了代偿的活动模式以保证正常的认知活动。这一研究结果可为未来选取睡眠剥夺期间进行恢复性干预措施的时间点提供一定的实证证据。
P2 成分反映了注意力和视觉加工的早期过程,与早期信息加工和选择性注意相关,反映了知觉过程中的早期判断[30]。睡眠剥夺后,注意力资源的分配会发生变化。相较于清醒状态,睡眠剥夺后大脑的显著性网络会将更多的注意力资源分配给默认模式网络,以维持个体的警觉性[31-32]。睡眠剥夺后,个体感觉门控功能受损,信息滤出存在障碍,影响个体的认知功能[33]。本文的结果显示,在经过24 h 睡眠剥夺后,志愿者的P2 波幅显著增加,说明志愿者睡眠剥夺后辨别和加工刺激信息的速度和能力降低,大脑对信息的选择、注意及抗干扰能力减弱[34]。睡眠剥夺后缩短的P2 潜伏期可能也说明了可支配注意力资源的减少[19]。以往研究表明,睡眠剥夺后最一致的负面影响是持续注意力的恶化,其对于高级认知功能的影响目前仍在讨论中[35]。
本文虽然采取了简单警觉性任务,但简单的反应任务仍然包含着早期感知觉、警惕性、注意、动作执行与相应等一系列心理机制[36]。研究发现睡眠剥夺后,志愿者的P3 成分的波幅显著下降。P3 成分在信息整合,传输有意注意、注意调节、反应后适应中起着重要的作用[37-38]。睡眠剥夺影响了后续的认知控制控制阶段。
已有不少研究表明,睡眠剥夺对PVT 任务和其他持续注意力任务存在强烈而一致的负面影响[20]。因此有研究人员提出假设,即睡眠剥夺对认知功能的负面影响是睡眠不足后个体警觉注意力不稳定的结果[13]。换而言之,警觉注意最容易受到睡眠剥夺的影响。
本文研究发现,24 h 睡眠剥夺后的个体注意警觉水平就已经大幅度受损,表明了注意警觉功能的敏感性。任何需要获取信息的认知任务的表现都可能因睡眠不足而受损;任务越需要保持警惕,睡眠不足造成的损害就越大[20]。警觉注意是一种会受到睡眠剥夺持续影响的认知成分[38]。注意模型强调警觉和持续性注意是进行高级认知加工时不可或缺的,并且如果个体不能很好地保持警觉水平时,其高级认知功能也会受到损害[39]。睡眠剥夺对工作记忆、执行功能的影响可能存在时间累积效应,只有当睡眠剥夺的时间足够长时,负面效应才足够明显。个体维持清醒的能力却是受到自上向下的皮层控制系统调节,尤其是前额叶的执行控制区域[40-41]。当睡眠压力较低时,这个系统处于松弛状态,清醒状态的警觉表现也处于稳定状态。当个体持续维持24 h 的清醒状态时,逐渐增加的渴望睡眠的身体内稳态压力和维持清醒的动机之间的冲突就会增加,从而导致了警觉系统的不稳定性和不可靠性。
本研究的不足是实验采用的都是男性志愿者,增加女性样本和功能磁共振成像的研究对于睡眠剥夺影响警觉注意的神经机制阐述和群体差异性有重要的帮助作用。
5 结论
1)完全睡眠剥夺会影响个体的注意警觉性,警觉认知任务表现出对睡眠剥夺的高敏感性。
2)警觉注意力并非会随着剥夺时间的增加而线性下降,睡眠剥夺24 h 后,个体的认知能力进入一个平台期,个体认知能力急速下降,大脑采用了代偿活动模式以保证正常的认知活动。
3)在日常生活中,健康人在安静觉醒的状态下,警觉状态的维持相对稳定,但平衡状态一旦遭到破坏,个体的注意警觉状态就会受到影响。
