混交比例对西南桦和红锥早期生长与竞争的影响
2023-12-25邹扬东王春胜尹海锋贾宏炎
邹扬东,王春胜,尹海锋,贾宏炎,董 琼,曾 杰
(1. 西南林业大学 林学院,云南 昆明 650224;2. 中国林业科学研究院 热带林业研究所,广东 广州 510520;3. 中国林业科学研究院 热带林业实验中心,广西 凭祥 532600)
混交林相较于纯林,具有促进林木生长[1-2]、改善土壤养分状况[3]、提高生物多样性[4]以及林分稳定性[5]等优点,从而受到学者越来越多的关注,尤其对于混交林生长和产量探索已成为混交林研究的主要内容[6]。对于混交林而言,适宜的混交比例,是混交林成功营建的基础,对于提升林分生产力至关重要[7]。杨嘉麒等[8]基于4 种混交比例的桉树Eucalyptusspp.与红锥Castanopsis hystrix混交林生长和土壤养分分析,筛选出3 行桉树与3 行红锥的混交模式,其混交效果最佳;廖科等[9]通过调查分析栎类纯林以及不同混交比例的栎类与枫香、栎类与杉木混交林,发现7 栎类3 枫香和3 栎类7 枫香混交处理的栎类胸径和材积显著高于其他处理。由此可见,对于一个树种组合而言,开展其混交比例研究是构建合理混交模式的关键所在。
归根结底,林木生长及活力与林分内竞争强度呈负相关[10],合理混交配置可降低林分内树体间竞争,从而实现林分生产力的提升。林分内竞争强度主要以种内、种间以及单木竞争指数表征,而竞争指数通常依据林木胸径、树高、冠幅和与相邻树木的距离量化[11],林分内竞争强度随着林木径级增大而减小[12-14]。然而,有关混交比例对林木竞争的影响尚不明确。汪清等[15]通过对不同混交比例马尾松-木荷混交林研究发现,马尾松Pinusmassoniana的种内竞争强度随着木荷Schimasuperba比例的增加而减小。因此,研究不同混交比例林分内林木竞争状况,有助于增进对于混交林内林木竞争的理解,为混交林成功营建提供科学依据。
西南桦Betulaalnoides为热带、南亚热带地区的一种珍贵阔叶树种,具有速生、材质优良等特点。截至2015 年底,西南桦造林规模已达到150 000 hm2。然而,大面积纯林经营引发林分生长不良、蛀干害虫危害以及林木分化严重等诸多问题[16-17]。红锥Castanopsishystrix亦为该地区的一个重要阔叶用材树种,其适应性较强,生长较快,幼树喜阴[18-19]。有研究表明,西南桦与红锥混交不仅有助于降低西南桦的蛀干害虫为害风险,而且能促进林木生长[20],西南桦与红锥混交逐渐受到关注,在生产上得以广泛推广应用。目前西南桦与红锥大多采用为1∶1混交比例,而何种比例最佳尚不明确。因此,本研究以6 年生西南桦与红锥不同混交比例试验林为对象,探究西南桦和红锥生长以及林木竞争对不同比例混交的响应,明确西南桦与红锥合理的混交比例,为优化其混交林经营提供科学依据。
1 试验地概况
试验地位于广西壮族自治区凭祥市中国林业科学研究院热带林业实验中心伏波分场(22o02′N,106o52′E)。该地属于北热带季风气候,冬无严寒,夏无酷热。年均气温为21.0 ℃,1 月和7 月平均气温为11.0 和27.5℃,极端最高和最低气温为38.7 和-1.2 ℃;年均降水量1 550 mm,年均蒸发量1 400 mm,干湿季明显,其中5—9 月为雨季,其降雨量占全年的3/4;年均空气湿度为80%以上,年平均日照1 614 h,全年无霜期344 d。土壤为花岗岩发育的酸性红壤,立地类型属Ⅱ类,土层厚度>100 cm。试验地海拔为420 m。
西南桦与红锥混交试验林营建于2017 年3 月,共设置3 个混交比例处理(西南桦与红锥比例分别为1∶1、1∶3 和1∶5)以及西南桦和红锥2 个纯林处理。试验按随机区组设计,每个处理3 次重复,共15 个小区,每个小区面积为3 333.35 m2以上。1∶1、1∶3 和1∶5 处理分别按1 行西南桦1 行红锥、1 行西南桦3 行红锥以及1 行西南桦5行红锥在其小区内排列。西南桦采用优良无性系组培苗造林,红锥采用以来自当地母树林种子培育的实生苗造林,造林株行距为2 m×3 m。试验林的保存率为85%。
2 研究方法
2.1 样地布设与生长观测
2023 年4 月,于每个小区中间位置设置20 m×30 m 样地,调查每个样地内胸径(DBH)>1 cm的所有单株的生长情况。具体而言,采用胸径尺(精度0.1 cm)测量DBH,采用超声波测高仪(Haglof Vertex IV、瑞典;精度0.1 m)测定树高(H)、枝下高(HCB)和冠幅(CW)。冠幅为从树干到东、西、南和北4 个方向树冠边缘距离平均值的2 倍,并计算高径比(H/D)、冠长(CL)、树冠率(CR)、冠体积(CV)、冠表面积(CSA)[21]。
2.2 林木竞争测定
1)竞争范围确定
根据林分内胸径最大西南桦和红锥单株的树冠确定竞争影响范围[22]。据样地调查,混交试验林内西南桦和红锥的最大冠幅分别为5.7和3.8 m,因此,将西南桦和红锥竞争范围分别确定为6 和4 m,将竞争范围内的所有单株列为竞争木。
2)林木竞争指数计算
各样地内每个树种选取5 株作为对象木,对象木选择标准为:生长正常,且竞争范围内竞争木均正常存活。采用Hegyi 提出的与距离有关的竞争指数计算各林分内对象木种内和种间竞争指数,其计算公式参考文献[15]。
2.3 数据分析
采用SPSS 13.0 软件进行单因素方差分析、Duncan 多重比较以及Pearson 相关性分析。
3 结果与分析
3.1 林木生长对混交比例的响应
由表1 可知,混交对西南桦生长具有显著的促进作用,西南桦树高、胸径以及胸高断面积在3个混交处理间差异不显著(P≥0.05),均显著高于西南桦纯林处理(P<0.05)。混交林的西南桦冠幅、冠体积以及冠表面积均显著高于西南桦纯林,以1∶3 混交处理为最大,显著高于其他混交处理。而西南桦枝下高和树冠率以纯林最高,且均显著高于1∶3 和1∶5 处理;而其冠长则正好相反,以1∶3 和1∶5 处理为最高,显著高于纯林;1∶1 处理与其他处理间西南桦树冠率和冠长的差异均达显著水平。对于高径比而言,1∶1 和1∶5混交处理均显著低于纯林,而这些处理与1∶3 处理差异不显著(表1)。
红锥对混交的响应较西南桦稍弱,混交处理对红锥树高生长无显著影响,但显著增加了红锥胸径、冠幅、胸高断面积、冠长、冠体积和表面积,以1∶1 混交处理为最高,均显著高于纯林。与西南桦类似,混交也显著降低了红锥枝下高和树冠率,各混交处理红锥枝下高和树冠率均显著低于纯林,各混交比例处理间差异均不显著。此外,1∶1 和1∶3 混交处理的红锥高径比均明显低于纯林处理,而1∶5 处理与纯林间差异未达显著水平。
3.2 林木竞争对混交比例的响应
由图1 可知,林分内西南桦种内竞争指数随红锥比例的上升整体上呈现下降趋势,以纯林处理为最高,显著高于3 个混交处理;其次为1∶1处理,也显著高于其余两个混交处理。3 个混交比例处理比较,1∶3 处理的西南桦种间竞争指数最高,显著高于1∶1 和1∶5 处理。综合种内和种间竞争,西南桦在其纯林中的竞争指数也显著高于各混交比例处理,而各混交处理间无显著差异(图1)。
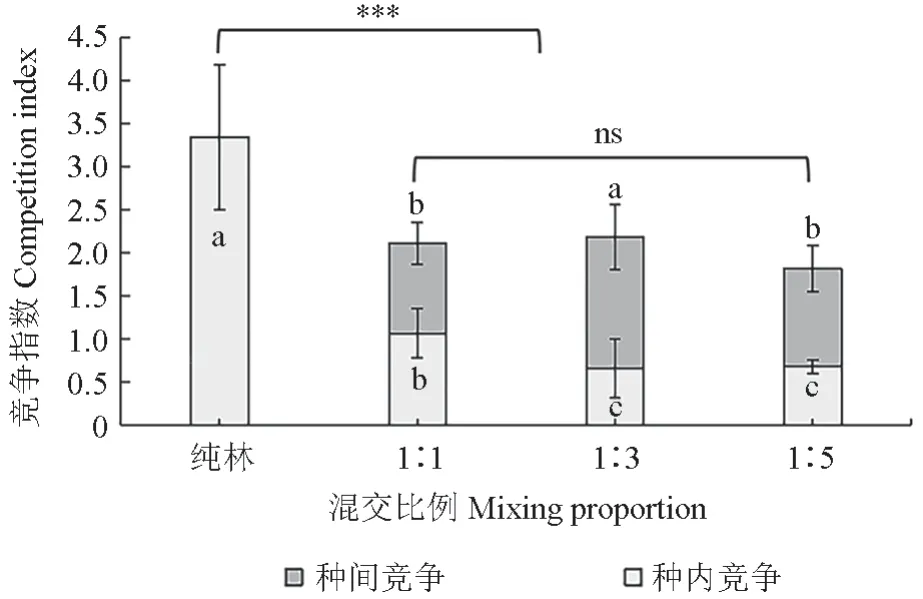
图1 西南桦和红锥不同比例混交林内西南桦的竞争指数Fig. 1 Competition index of B. alnoides in its mixture with different proportions of C. hystrix
与西南桦相反,林分内红锥种内竞争指数随红锥比例的升高呈现增大趋势,纯林内红锥种内竞争指数显著高于各混交比例处理,而1∶3 和1∶5处理间差异未达显著水平,均显著高于1∶1 处理。红锥种间竞争指数随红锥比例的升高呈现下降趋势,其中1∶1 处理种间竞争指数显著高于1∶3 和1∶5 处理。整体而言,1∶1 处理的红锥种内与种间竞争指数之和最高,显著高于红锥纯林和其他混交处理(图2)。

图2 西南桦和红锥不同比例混交林内红锥的竞争指数Fig. 2 Competition index of C. hystrix in its mixtures with different proportions of B. alnoides
3.3 林木生长与竞争指数间的相关性
由表2 可知,西南桦和红锥的种内竞争指数与其自身的胸径、树高、冠幅、胸高断面积、冠长、冠体积、冠表面积呈现极显著(P<0.01)或显著(P<0.05)负相关,与高径比呈极显著或显著正相关,与枝下高相关不显著(P≥0.05)。对于种间竞争指数与林木生长的相关性而言,西南桦与红锥间的差异较大。西南桦的种间竞争指数与其胸径、树高、枝下高、冠幅、胸高断面积、冠体积、冠表面积极显著或显著负相关,与树冠率、冠长相关不显著;而红锥的种间竞争指数仅与其树高呈显著负相关,与其他生长指标均相关不显著(表2)。
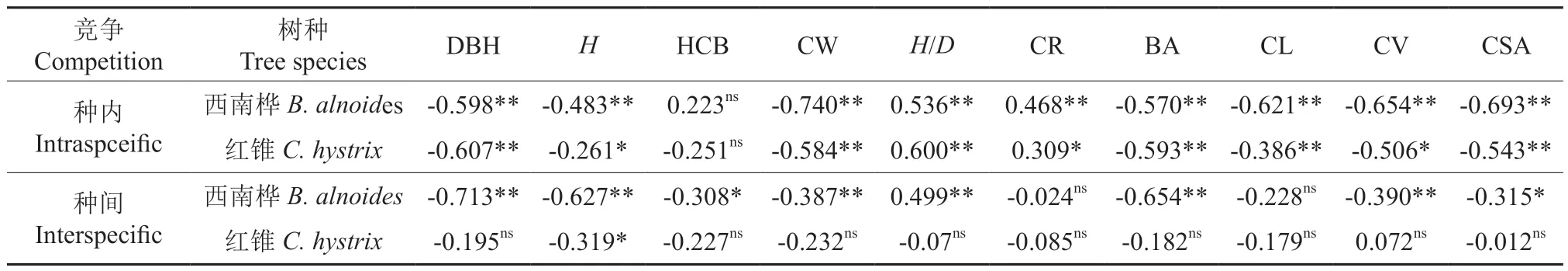
表2 西南桦和红锥混交林内林木生长表现与竞争指数的相关性Table 2 Correlation between growth performance and competition index for B. alnoides and C. hystrix in their mixtures
4 讨 论
在本研究中,混交显著促进了西南桦的生长,各混交处理的西南桦胸径、树高、冠幅、胸高断面积、冠长、冠体积以及冠表面积均显著高于纯林。究其原因,与西南桦和红锥混交显著降低了其在林分中所承受的竞争强度有关。一般而言,树种间生态习性、生态幅度差异越大,其竞争越小[23]。西南桦是一个速生的强阳性树种,而红锥早期生长相对较慢且耐阴,两者同龄混交后,西南桦迅速占据林冠上层;相对于纯林而言,混交林内的西南桦具有更大的生长空间以及较低的竞争强度,从而促进了树冠的扩展及其对光资源的获取,最终在西南桦的各项生长指标上得以体现。Aldea 等[24]对苏格兰松Pinussylvestris-挪威云杉Piceaabies混交林的研究结果表明,苏格兰松在混交林中占据着上层空间,与其纯林相比,减少了同一冠层内的竞争,因而其生长更快。西南桦树高和胸径随着红锥比例的增加规律不明显,且不同比例混交处理间差异不显著(P≥0.05),其主要与各混交处理间西南桦冠层尚未郁闭有关。对于已郁闭的混交林而言,其规律性则增强。如,Forrester 等[25]对不同比例蓝桉Eucalyptusglobulus-黑木相思Acaciamangium混交林的研究表明,11 年生时蓝桉(处于上层)的树高和胸径生长量随着黑木相思比例的增大呈现升高趋势。
对于红锥而言,混交促进了其树干径向生长和树冠发育,这与欧建德等[26]对杉木Cunninghamia lanceolata-乳源木莲Manglietiayuyuanensis混交林的研究结果部分一致;混交显著促进了乳源木莲胸径、树高和单株材积的生长。在本研究中,各树干径向生长和树冠发育指标绝大多数仅在1∶1混交处理与纯林间的差异达到显著水平。造成此现象的原因可能是:其一、适当遮阴有利于红锥的早期生长,相比于1∶3 和1∶5 混交处理,1∶1混交处理中所有红锥更靠近西南桦,尽管与西南桦的竞争强度更大,但是西南桦提供的遮阴环境更适合其生长;其二、由于西南桦与红锥存在空间互补,1∶1 混交处理中红锥生长空间大于1∶3和1∶5 混交处理,从而降低了林分内红锥种内竞争强度。总而言之,红锥生长受种内竞争的影响要明显高于种间竞争。Forrester 等[25]的研究表明,11 年生时黑木相思(处于下层)的树高和胸径生长量随着蓝桉比例的增大而减小,与本研究明显不同,与林分已郁闭,种间竞争加剧有关。
各混交处理的西南桦和红锥枝下高、树冠率大多显著低于相应纯林,其中1∶1 处理中西南桦枝下高显著高于1∶3和1∶5混交处理,这是由于1∶1处理中西南桦的种内竞争显著大于其余混交处理,且该处理中红锥生长状况优于1∶3 和1∶5 混交处理,从而促进了西南桦的自然整枝。而Liu 等[27]的研究表明,西南桦分别与思茅松Pinuskesiyavar.langbianensis、格木Erythrophleumfordii混交,西南桦的枝下高小于纯林,格木的枝下高则显著大于纯林,思茅松在混交林和纯林间差异不显著,各混交模式间的差异可能与混交树种的生长习性及对光环境的适应性有关。Forrester 等[25]研究发现,蓝桉和黑木相思的高径比分别随着黑木相思和蓝桉比例的增大而下降,与竞争强度有关。在本研究中,混交处理的高径比大多显著小于纯林,3 个比例混交处理间西南桦高径比基本稳定,红锥的高径比则随着西南桦比例的增大而降低。这主要是由西南桦与红锥的生长特性决定。两者混交形成了空间互补,有效降低了林分内的竞争强度,而纯林因树冠处于同一冠层,种内竞争强度大,造成西南桦和红锥纯林内树高生长优于径向生长,以获得更多的生长空间,造成两者纯林高径比显著高于混交林。
西南桦在其纯林内所承载的竞争强度显著高于混交林,而不同比例混交处理间竞争强度随着红锥比例的增加呈现下降趋势,这与汪清等[15]对不同比例马尾松-木荷混交林内马尾松的竞争强度变化趋势相似。然而,本研究中红锥则在混交比例为1∶1 时承载的竞争强度显著高于纯林和其他混交比例处理。造成此差异的原因,一方面与混交树种生长特性有关,因为西南桦生长速度比红锥快,且西南桦为强阳性树种,而红锥偏阴性,两者在空间利用和习性上形成互补;另一方面,由于红锥的竞争范围为4 m,因此混交比例1∶1处理内红锥单株的竞争木中西南桦比例高于其他混交处理,而Hegyi 提出的竞争指数与对象木的胸径大小密切相关,并且西南桦胸径显著大于红锥。因此,红锥在混交比例为1∶1 时在林分中所承载的竞争强度最高。此亦可从竞争指数与对象木生长指标的负相关性得到验证。
由于本研究仅对6 年生西南桦和红锥的混交林进行评价,还需要对其混交效应进行持续跟踪研究,揭示其生长和竞争的动态变化,更好地为其种间关系调控提供科学依据。
5 结 论
综上所述,本研究分析了广西凭祥6 年生西南桦和红锥纯林及其不同比例混交林的生长表现和竞争状况,整体而言,西南桦与红锥不同比例混交可有效促进两者生长的协同提升和竞争强度的降低,而且其促进作用因混交比例而异;西南桦和红锥的种内和种间竞争强度与其自身的生长大多呈显著负相关。综合考虑西南桦和红锥的生长表现、自然整枝状况以及林分空间利用状况,建议我国南方地区西南桦与红锥采用1∶1 的比例营建混交林。
