优化发酵条件促海鞘桔青霉Asc2-4生长与产油
2023-12-21罗正东刘唤明雷晓凌颜珲璘
罗正东,刘唤明,雷晓凌,颜珲璘
(1.广东海洋大学 食品科技学院 广东省水产品加工与安全重点实验室 广东省海洋食品工程技术研究中心 广东省海洋生物制品工程实验室 水产品深加工广东普通高等学校重点实验室,湛江 524088;2.中国科学院深圳先进技术研究院 支撑平台处高等级生物实验室管理办,深圳 518055)
海洋共附生微生物分离于海洋生物体,包括海洋脊椎动物、海洋无脊椎动物和海洋植物,生长于恶劣且复杂多变的海洋生态环境,具有生长能力强、抗逆性强、产物结构多样等特点[1-2]。海洋产油真菌增殖快,能提供足够的胞内空间积累油脂以抵御低温,也能为浮游生物和附生动物提供脂类营养物质[3-4]。因其独特的油脂合成代谢机制,能够生产中长链脂肪酸,有潜力开发为保健品、食品、医药和燃料[5-6]。前期研究发现海鞘共附生真菌桔青霉Asc2-4具有产油菌的共同特点[7-8]。
发酵条件对产油真菌的菌丝体生长和油脂形成起到不同的效果。赵晨等[9]发现解脂耶氏酵母(Yarrowialipolytica)在低温条件下有利于合成不饱和脂肪酸,高温有利于合成饱和脂肪酸。pH能改变生物膜通透性保证足够的营养物质和氧气输入胞内促进菌体生长,也能影响脂肪酸的组成。Mironov等[10]研究高山被孢霉(Mortierellaalpina)NRRL-A-10995表明,pH值为3时能完全抑制花生四烯酸(AA)的合成,当pH值为6时,AA的百分含量和生物量皆达到最高,分别为19.9%和22.0 g/L。接种量对产油真菌的生长曲线趋势有较大影响,适宜的接种量既能使菌丝体快速增殖从而缩短发酵周期,也能让对数生长末期的菌丝球表面积增大,吸收充足的氧气及养分,从而提高菌体的油脂积累量[11-12]。溶氧量是微生物高产油脂的关键因素,可通过转速和装液量优化溶氧,Magdouli等[13]发现解脂耶氏酵母(Yarrowialipolytica)SM7在溶解氧含量30%时油脂含量能够达到44.81%。肖玉娟等[14]在红树林海水腐木中分离出一株高产油脂的破囊壶菌(Aurantiochytriumsp.)HX2019010,发酵油经GC-2010 PLUS气相色谱鉴定,测算出该菌产9.17 g/L的DHA。因此,研究发酵条件的优化对提高生物量和油脂质量浓度十分必要,通过测定发酵油的脂肪酸组分和含量可判断油脂功能性。
在前期研究的基础上,通过单因素和响应面优化发酵条件,进一步提高桔青霉Asc2-4的生长和产油能力,根据气相色谱测定的脂肪酸组分及含量评价菌油的功能性,为海鞘共附生微生物桔青霉Asc2-4开发成工业产油菌株奠定基础。
1 材料与方法
1.1 材料
1.1.1 菌株
桔青霉PenicilliumcitrinumAsc2-4为课题组前期分离和鉴定的优良产油真菌,筛选自广东省雷州市流沙虾塘的海鞘[15]。保藏于广东省微生物菌种保藏中心(保藏号:GDMCC 60059)。
1.1.2 培养基
种子培养基:马铃薯浸出粉8 g/L,葡萄糖20 g/L,海精盐15 g/L,硫酸铵10 g/L,pH(5.8)自然,饱和蒸汽121 ℃灭菌20 min。
发酵培养基:新鲜马铃薯200 g/L,葡萄糖100 g/L,蛋白胨0.5 g/L,酵母膏0.5 g/L,海精盐15 g/L,柠檬酸钠0.1 g/L,KH2PO44 g/L,MgSO4150 mg/L,FeCl32.0 μg/L,pH值5.8,饱和蒸汽121 ℃灭菌20 min。
1.1.3 主要仪器
HZQ-F160振荡培养箱;BMJ-25型霉菌培养箱;LS-B50L立式压力蒸汽灭菌锅;UGC-24C圆形水浴氮吹仪;GC-2010 PLUS气相色谱仪。
1.2 方法
1.2.1 种子液培养
接种适量桔青霉Asc2-4孢子于含有200 mL种子培养基的500 mL锥形瓶中,在28 ℃、180 r/min的摇床中培养48 h。
1.2.2 发酵液培养
将种子培养后的菌丝体接种于含有100 mL发酵培养基的250 mL锥型瓶,在摇床中发酵培养。
1.2.3 菌体生物量测定
参照文献[16]菌体生物量测定方法,略有改变。抽滤菌丝体于培养皿,在109 ℃的烘箱中烘干,恒重后称重。
1.2.4 菌体油脂提取
参考文献[17]酸热提油法,略有改进。将每1.00 g的干菌体添加6.00 mL浓度为4.00 mol/L的盐酸,在温度100 ℃的沸水浴和-20 ℃冷冻环境中分别处理10 min,重复2至3次,按2∶1(体积比)的氯仿和甲醇溶液萃取,在2 000 r/min、5 ℃条件下离心,取上层有机相氮吹即得菌油。
1.2.5 菌油脂肪酸组成测定
菌油样品甲酯化[18],使用气相色谱仪进行脂肪酸组分及含量分析,气相色谱仪配置SP-2560气相毛细管柱(100 m×0.25 mm×0.2 μm)和氢火焰离子检测器(FID)。设置进样口温度为225 ℃,检测器温度为250 ℃,分流比10∶1;设置程序升温条件:柱温100 ℃(13 min),以10 ℃/min升至180 ℃(6 min),再以1 ℃/min升至200 ℃(20 min),最后以4 ℃/min升至235 ℃ (20 min)。
1.3 单因素优化
以28 ℃、pH值5.8、接种量5%、装液量100 mL/250 mL、转速140 r/min和培养时间7 d作为初始发酵条件,分别设置温度25、27、29、31和33 ℃,pH(4.5、5.5、6.5、7.5和8.5)、接种量(1%、3%、5%、7%和9%)、250 mL三角瓶装液量(50、75、100、125和150 mL)和转速(100、140、180、220和260 r/min),每组进行3次平行实验,综合生物量和油脂含量筛选出适宜桔青霉Asc2-4的发酵条件。
1.4 响应面优化
选取单因素中3个显著影响油脂积累的因子并设计三水平响应面试验,以油脂浓度作为响应值,共设置15组试验,进行3次中心重复试验,试验设计水平及编码见表1。
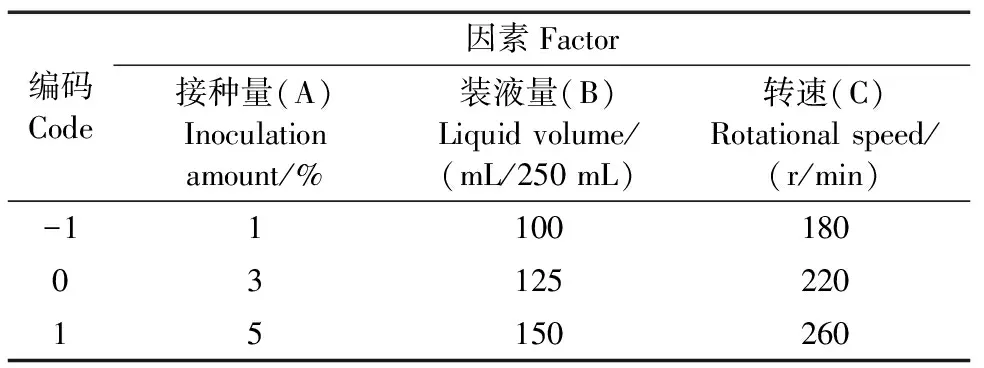
表1 响应面试验因素水平和编码
1.5 数据处理
单因素试验采用SPSS 22.0进行数据分析,使用One-way ANOVA程序对数据进行方差分析,Duncan多重分析比较验证数据,P<0.05为差异显著,响应面试验使用软件Design-Expect 10.1拟合模型和数据分析。
2 结果与分析
2.1 海鞘桔青霉Asc2-4产油发酵条件的单因素优化
2.1.1 最佳发酵温度
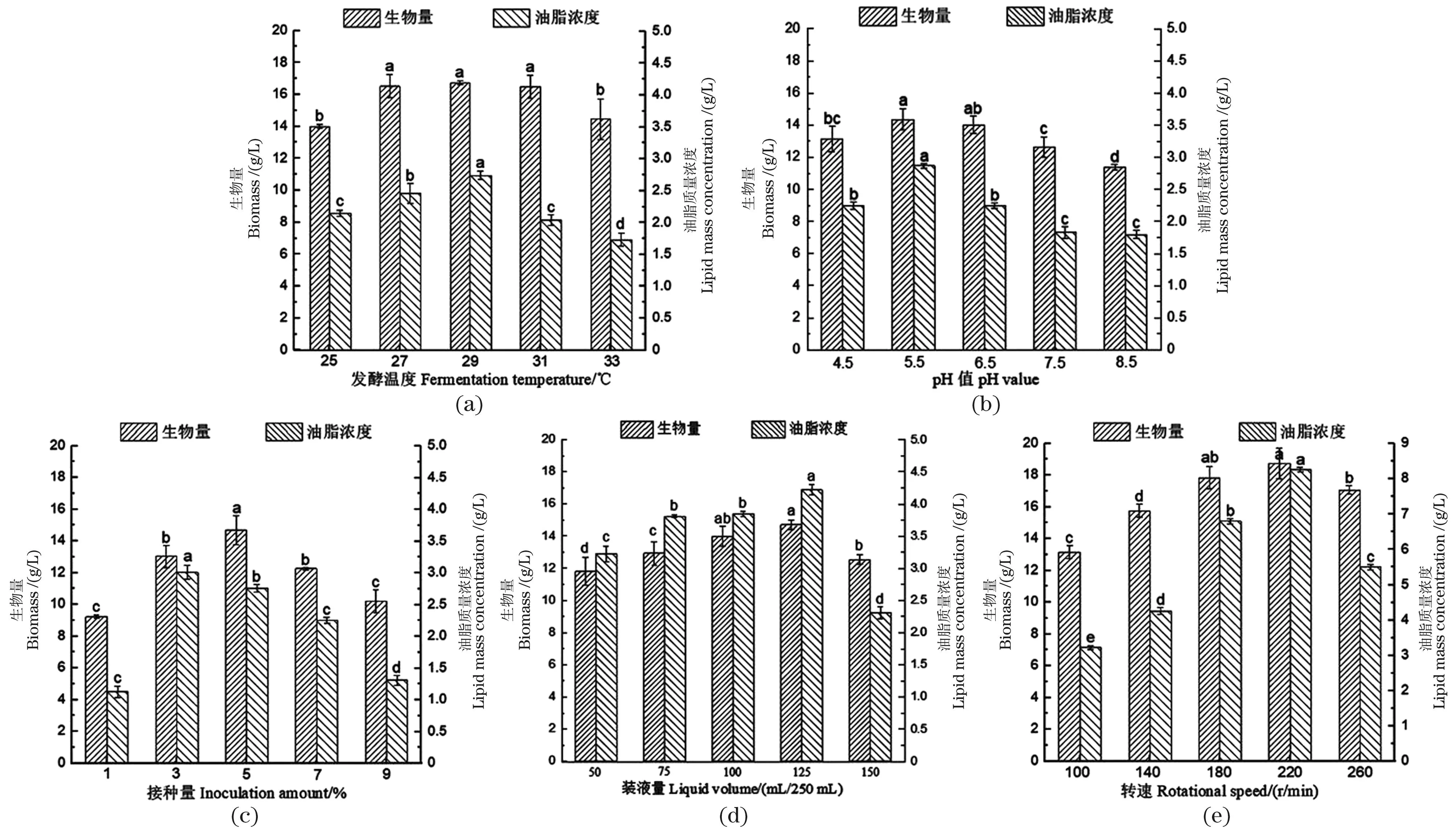
由图1(a)可知,桔青霉Asc2-4的生物量在25~29 ℃范围内随温度升高而增大,超过29 ℃则出现下降。不同温度对该菌的生物量影响程度存在差异,27、29和31 ℃的生物量较高而三者的差异不显著,但与温度25和33 ℃的生物量差异显著(P<0.05)。其中,29 ℃的生物量最高,达到16.73 g/L。29 ℃的油脂质量浓度为2.74 g/L,显著高于其他4种温度的油脂质量浓度。因此,确定29 ℃作为最佳发酵温度。
同一指标含一个相同字母则差异无统计学意义(P>0.05)。
2.1.2 最佳pH值
由图1(b)可知,桔青霉Asc2-4的生物量随pH值的增大呈现出先升高后下降的趋势,这种变化与pH影响菌体细胞通透性和参与调控生长的酶蛋白活性,导致菌体受增殖促进或抑制的情况有关[19-20]。当pH值为5.5时生物量达14.36 g/L,说明该pH值有利于菌丝体的生长。当pH值为4.5~5.5时,油脂质量浓度呈现上升,而pH值5.5~8.5时出现下降,适宜产油的pH值为5.5,油脂质量浓度为2.87 g/L。因此,确定5.5为最佳pH值。
2.1.3 最佳接种量
由图1(c)可知,桔青霉Asc2-4在5种接种量条件下的生物量和油脂质量浓度皆有不同程度提高。但是,适宜菌体增殖的最佳接种量与适宜油脂积累的有所不同,其中,接种量5%对生物量提高显著(P>0.05)达到最高的14.68 g/L。但最佳产油的接种量则为3%,油脂质量浓度为3.01 g/L,这说明生物量适宜能为积累油脂吸收更多营养成分或是溶氧量,生物量过高反而不利于微生物产油[21]。本研究为提高产油,所以将3%作为最佳接种量。
2.1.4 最佳装液量
由图1(d)可知,桔青霉Asc2-4在125 mL/250 mL时生物量和油脂质量浓度皆为最高,分别达到14.69和4.22 g/L。有报道称[6,22],装液量能为产油菌提供活性氧,刺激油脂合成途径的碳链去不饱和及碳链合成。然而,过高的装液量易造成种间竞争影响菌体生长和油脂生产,当装液量为150 mL/250 mL时,其生物量(12.53 g/L)和油脂质量浓度(2.31 g/L)相较装液量125 mL/250 mL皆出现降低。综合考虑,确定125 mL/250 mL作为最佳装液量。
2.1.5 最佳转速
由图1(e)可知,转速从100 r/min到220 r/min,该菌的油脂质量浓度从3.22 g/L大幅提升至8.26 g/L,可知,适宜产油的转速为220 r/min。结合图1(d)可知,装液量及转速不仅能大幅提高油脂质量浓度,其生物量变化趋势也相似,这与该两种发酵条件皆通过改变液态剪切力影响菌丝体的增殖有关[23-24]。转速220 r/min的生物量为最高的18.73 g/L,刺激高产油的转速也为220 r/min,其油脂质量浓度为8.26 g/L。因此,确定220 r/min为最佳转速。
综上可知,桔青霉Asc2-4发酵条件单因素优化的结果:29 ℃、pH值5.5、接种量3%、装液量125 mL/250 mL和转速220 r/min,该发酵条件下适宜该菌生长与产油。为进一步提高该菌的产油能力,选取明显提高油脂质量浓度的因子(接种量、装液量和转速),作为响应面优化的研究对象。
2.2 海鞘桔青霉Asc2-4产油发酵条件的响应面试验
2.2.1 响应面试验设计与优化

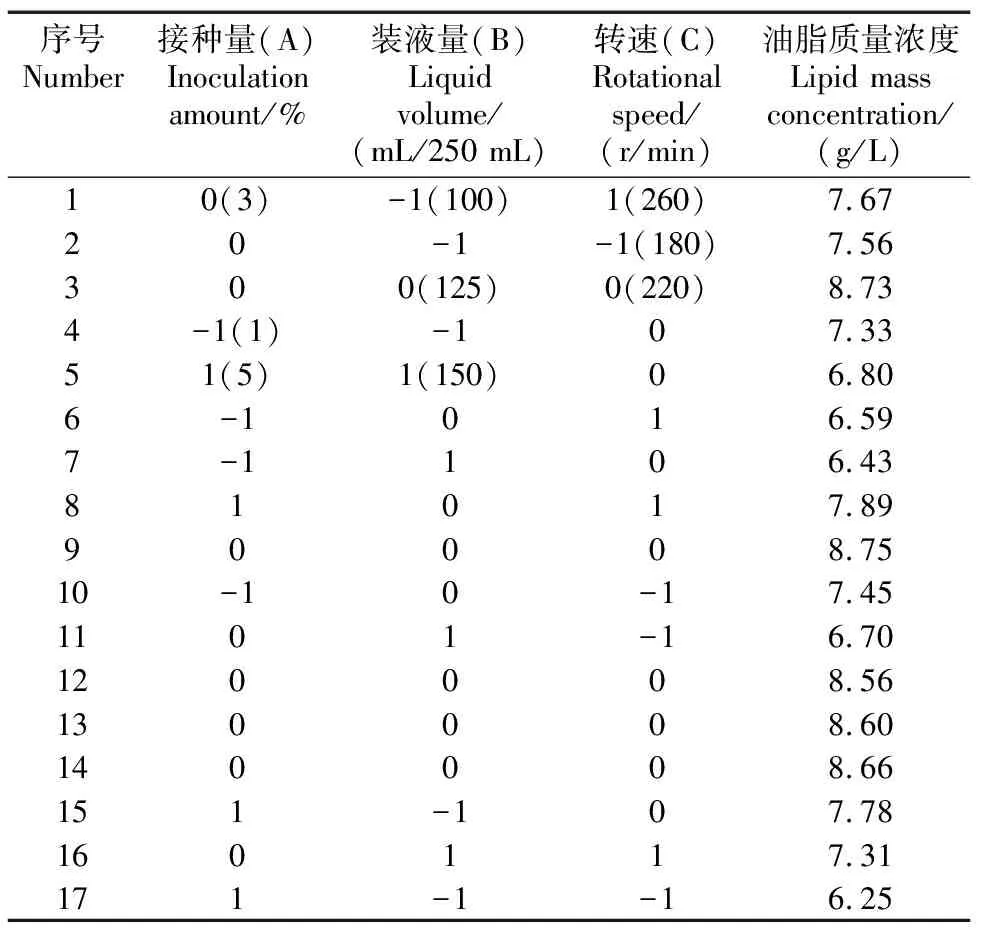
表2 发酵条件响应面试验优化
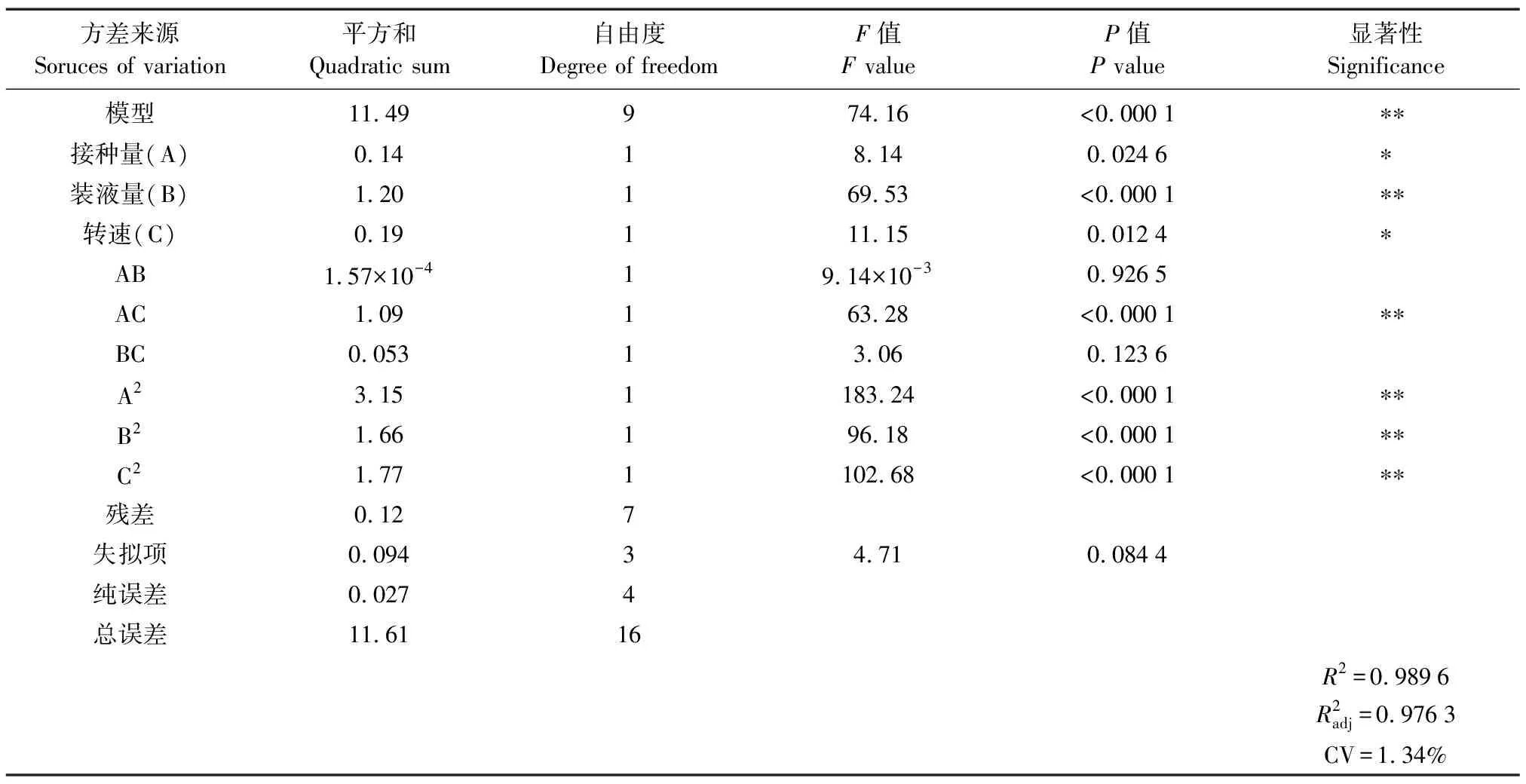
表3 回归方程方差结果
2.2.2 响应面试验的等高线与响应曲面
图2为响应面试验的等高线与响应曲面。不同交互效应的强弱可由等高线图形进行判断,圆形代表两因素间的交互显著,椭圆形则代表交互不显著,可知,接种量与转速的交互显著。结合Design-Expert软件分析数学模型得桔青霉Asc2-4在发酵条件为接种量3.26%,装液量118.42 mL/250 mL,转速226.20 r/min,油脂质量浓度最大。结合方差分析和实际情况,确定接种量3.3%、装液量120 mL/250 mL、转速220 r/min为最佳产油发酵条件。
2.2.3 验证响应面优化
表4为验证响应面优化的比较试验。经验证,得到3次试验的油脂质量浓度平均值为8.80 g/L,与理论响应值(8.78 g/L)相差0.02 g/L,表明该响应面结果真实可靠。最终结果,生物量和油脂质量浓度分别提高了65.57%和224.73%,而油脂含量从22.39%提升了109.30%,达到45.88%。
2.3 海鞘桔青霉Asc2-4菌油脂肪酸组分与含量的测定
表5为气相色谱测定的菌油脂肪酸组分与含量。该菌能够生产高级脂肪酸(C15~C24),其中C16和C18脂肪酸相对含量达96.26%。主要脂肪酸为棕榈酸(C16:0)、硬脂酸(C18:0)、油酸(C18:1)和亚油酸(C18:2)。发酵油的饱和脂肪酸共7种,占35.60%,不饱和脂肪酸6种,占64.40%;最高含量的脂肪酸为41.46%的C18:1,其余3种含量较高的脂肪酸为21.68%亚油酸(C18:2)、16.64%棕榈酸(C16:0)和15.58%硬脂酸(C18:0)。
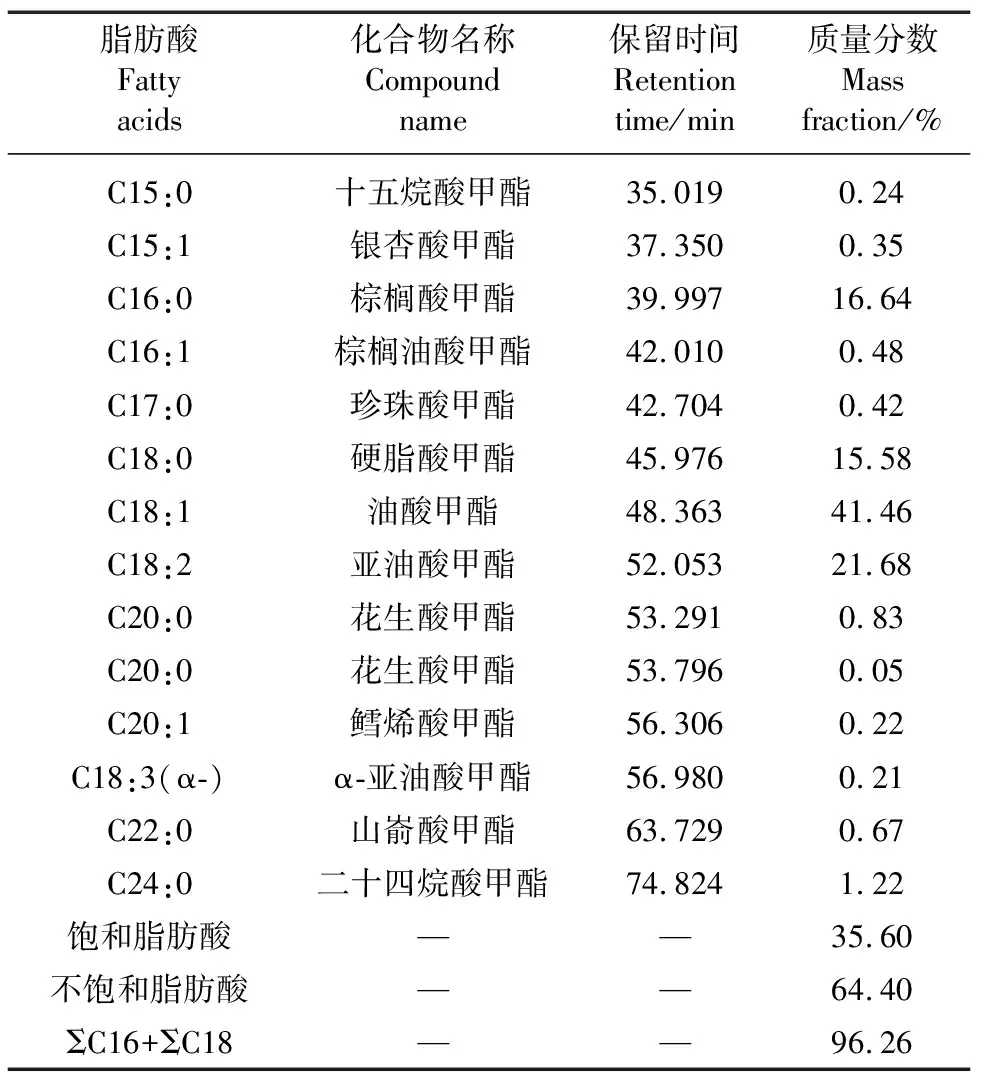
表5 发酵油的脂肪酸组分及含量
3 结论
研究表明,适宜桔青霉Asc2-4生长与产油的发酵条件为温度29 ℃、pH值5.5、接种量3.3%、装液量120 mL/250 mL和转速220 r/min,生物量和油脂质量浓度分别达到19.19和8.80 g/L,其主要脂肪酸为C16和C18脂肪酸,占总脂肪酸96.26%,各脂肪酸含量与组分分别为41.46%油酸(C18:1)、21.68%亚油酸(C18:2)、16.64%棕榈酸(C16:0)和15.58%硬脂酸(C18:0)。
优化发酵条件能提高代谢效率和充分吸收营养物质使生物量和油脂浓度得到提升[25]。本研究进行温度单因素试验发现,温度对生物量的影响大于油脂浓度,这与温度更能影响真菌增殖有关[26],而低温条件(25~27 ℃)的油脂浓度皆低于29 ℃的油脂浓度(P<0.05),则与普遍认为低温(<27 ℃)利于微生物积累油脂的理论不同[27-28]。pH通过改变细胞膜电荷影响营养物质的吸收率而提高油脂积累量,如龚佳等[29]发现圆红冬孢酵母(Rhodosporidiumtoruloides)在pH值为4~8时油脂产量变化为77.06~9.16 g/L,而本研究发现pH(4.5~8.5)时油脂质量浓度的变化范围在1.73~2.74 g/L,说明此pH值范围对桔青霉Asc2-4油脂积累影响不明显。有研究发现[30],产油真菌帚枝霉(Sarocladiumkiliense)ADH17在优化接种量(5%~10%),其油脂质量浓度最高仅为1.17 g/L,而本试验接种量3%时油脂质量浓度高达3.01 g/L,可见低接种量更适宜桔青霉Asc2-4合成油脂。本试验菌种在较高装液量(125 mL/250 mL)条件培养,生物量和油脂质量浓度分别为14.69 g/L和4.22 g/L,相比马素梅等[31]研究的粘红酵母(Rhodotorulamucilaginous)在30 mL/250 mL培养生物量和油脂质量浓度分别为3.10和4.25 g/L,可知该菌具有高密度培养的潜能。高转速易使自溶酶释放导致菌体破裂内容物流出[32]。本试验菌种在转速220 r/min的高液体剪切力下菌体增殖和积累油脂皆得到提高,该转速使生物量高达18.73 g/L,油脂质量浓度为8.26 g/L。本研究根据响应面模型设计和验证结果,可知桔青霉Asc2-4优化后的最高生物量和油脂含量比优化前分别提高65.57%和224.72%,实验值与预测的响应值吻合良好。经过发酵条件优化后的桔青霉Asc2-4油脂质量分数达到45.88%,高于16.7%~30.0%的斯达氏油脂酵母(Lipomycesstarkeyi)、12.38%~30.87%的卷枝毛霉菌(Mucorcircinelloides)、22.1%的小球藻(Chlorellavulgaris)、23.9%的珊瑚色诺卡氏菌(Nocardiacoralline)等产油微生物[33]。
微生物油脂不仅为人类提供营养膳食和疾病预防所需物质,还在化石能源紧缺的当下有望成为替代能源[34-35]。目前,研究得较为成熟的产油真菌高山被孢霉(Mortierellaalpina)被认定是长链多不饱和脂肪酸生产的重要工业化菌株[36],与其所产的长链不饱和脂肪酸ARA相比,桔青霉Asc2-4的中链脂肪酸C16与C18脂肪酸含量较高,占总脂肪的96.26%。Papapolymerou等[37]研究微生物油脂发现,C16和C18脂肪酸含量占比高,能降低碘值,使油脂具有更好的点火质量,且作为生物柴油有较好的脂肪甲酯性能。因此,桔青霉Asc2-4发酵油具有开发为生物质能源的潜力。本研究为海鞘共附生微生物桔青霉Asc2-4开发成工业产油菌株提供了理论依据。
