乳及乳制品中的酶及对其关键品质的影响
2023-12-14郭蓉剧柠卞永霞王媛媛李彩虹牛书第于一凡李璞钰
郭蓉,剧柠,卞永霞,王媛媛,李彩虹,牛书第,于一凡,李璞钰
(1.宁夏大学食品科学与工程学院,银川 750021, 2.宁夏农产品质量标准与检测技术研究所,银川 750002, 3.宁夏畜牧工作站,银川 750004)
0 引 言
乳及乳制品中的酶种类繁多,一般根据来源不同,将其分为内源酶和外源酶。内源酶来源于动物的血浆、体细胞(如白细胞)和分泌细胞(如乳腺分泌细胞)的顶膜或细胞质[1],外源酶一部分是挤奶、运输、加工过程中混入乳中的微生物所产生的胞外酶;另一部分则是乳制品加工过程中添加微生物产生或直接添加功能性酶制剂,如凝乳酶、谷氨酰胺转氨酶等[2-3]。这些酶共同发挥作用,影响着乳及乳制品的品质。为了保障乳及乳制品在货架期内的品质及质量安全,研究乳及乳制品中的酶及其对关键品质的影响具有非常重要的意义。
1 乳及乳制品中的酶
1.1 内源酶
乳中存在的内源酶种类繁多,其中有脂肪酶、蛋白酶、黄嘌呤氧化酶(Xanthine oxidase,XO,EC 1.1.3.22)、过氧化氢酶(Catalase,CAT,EC 1.11.1.6)、乳过氧化物酶(Lactoperoxidase,LPO,EC 1.11.1.7)、和碱性磷酸酶(Alkaline phosphatase,ALP,EC 3.1.3.1),这些酶对乳及乳制品的品质影响较大。除此之外,乳中还存在有其他内源酶,比如L-乳酸脱氢酶(Lactate dehydrogenase,LHD,EC 1.1.1.27)可以催化乳酸向丙酮酸的可逆氧化,超氧化物歧化酶(Superoxide dismutase,SOD,EC 1.15.1.1)催化超氧化物阴离子的歧化,还有巯基氧化酶(Sulphydryl oxidase,EC 1.8.3.2)和γ-谷酰胺转肽酶(γ-glutamyl transpeptidase,GGT,EC 2.3.2.2)等酶[1]。
乳中的内源性脂肪酶是脂蛋白脂肪酶(Lipoprotein lipase,LPL,EC 3.1.1.34),它能诱导脂肪水解产生游离脂肪酸(Free fatty acids,FFAs)。该酶在乳腺分泌细胞中合成,分子量为100 ku,最适温度为37 ℃,最适pH 为9.2。LPL 在牛乳中含量约为2 mg/L,其中80 %以上与酪蛋白胶束连接,部分以可溶形式存在,少量与乳脂肪球膜(Milk fat globule membrane,MFGM)有关[4]。新鲜分泌的牛乳中,乳脂肪以小球形式被MFGM 包裹,脂解程度并不剧烈;一旦MFGM被破坏,牛乳会发生剧烈脂解[5]。LPL 不稳定,巴氏杀菌后残留活性小于10%。
纤溶酶(Plasmin,PL,EC 3.4.21.7)是乳中的主要的内源性蛋白酶,属于纤溶酶系统。乳中纤溶酶系统通过血液转运至乳腺细胞并分泌进入乳中,由PL,纤溶酶原(plasminogen, PG),纤溶酶原激活剂(plasminogen activators, PA),纤溶酶原激活剂抑制剂(plasminogen activator inhibitors,PAI) 和纤溶酶抑制剂(plasmin inhibitors, PI)组成[6],如图1 所示。纤溶酶由无活性的纤溶酶原在纤溶酶原激活剂的作用下产生,是一种碱性丝氨酸蛋白酶,分子量大小为88 ku,最适温度为37 ℃,最适pH 范围是7.5 ~8.0[4]。PL 在牛乳中含量在0.1~0.7 mg/L 之间,主要与酪蛋白胶束结合,作用于β-酪蛋白(β-Casein,β-CN)、αS2-酪蛋白(αS2-Casein,αS2-CN)和αS1-酪蛋白(αS1-Casein,αS1-CN),但无法水解κ-酪蛋白(κ-Casein,κ-CN)和乳清蛋白[7]。PL 对热稳定,受巴氏灭菌条件的影响很小,超高温处理(137 ℃,4 s)后仍残存30%~40%活性[8]。
黄嘌呤氧化酶通过嗜乳脂蛋白附着在MFGM 内表面,是乳脂肪球膜蛋白的主要成分(约占20%),与乳脂肪小球的分泌过程相关[4]。XO 拥有2 个完全对称且具有独立催化活性的亚基[9],是一种二聚体金属黄素蛋白,分子量约为300 ku,最适pH 为8.3。该酶可催化次黄嘌呤氧化为黄嘌呤,将黄嘌呤氧化为尿酸,同时将O2还原为H2O2[10]。原料乳中XO 经巴氏灭菌和均质化后活性降至30%以下[4]。以下为XO 催化的反应:
次黄嘌呤+水+氧气→黄嘌呤+过氧化氢
黄嘌呤+水+氧气→尿酸+过氧化氢
乳过氧化物酶以可溶形式存在于哺乳动物的乳汁中,与硫氰酸盐、H2O2共同构成的乳过氧化物酶系统(Lactoperoxidase system,LPS)具有良好的抗菌能力。LPO 与H2O2共同作用催化氧化SCN-,生成具有抗菌活性的OSCN-并生成水,激活乳本身的过氧化物酶形成天然的抗菌体系,起到抗菌作用[11]。该酶分子量在72.5 ~88 ku 之间,最适pH 为9.0。LPO 在牛乳中的含量丰富(30 mg /L),该酶相对耐热,巴氏灭菌后仍有70% 活性残留[4]。
过氧化氢酶是一类广泛存在于动物、植物和微生物体内的含四聚体血红素的末端氧化酶,可通过两步氧化还原反应将H2O2分解为水和氧气[12]。CAT 存在于牛的各处组织与细胞中,分子量为225 ku,通过能与膜结合的体细胞(主要是白细胞)进入牛乳[4]。奶牛罹患乳腺炎时,乳腺细胞壁对体细胞(特别是白细胞)渗透性提高,此时分泌的原料乳中CAT 活性与体细胞数成正比,因此,CAT 活性可作为检测奶牛罹患乳腺炎的一个指标[4]。
碱性磷酸酶是含有唾液酸的一类糖蛋白,最适pH 范围是9.0~10.5。牛乳中的ALP 主要分布在乳脂相中(30%~50%),通过与磷脂酰肌醇的键合与MFGM的微粒体膜结合[13]。ALP 的热稳定性高于乳中的大部分致病微生物,灭活需要的条件更加苛刻,ALP 活性常用作为检查牛乳巴氏杀菌是否彻底的指标[4,13]。但牛乳中微生物释放的ALP 的耐热性比原料乳中强,可能会造成检测假阳性现象或干扰测试结果[14]。
近年来,随着组学技术的发展,乳中一些新的内源酶被发现。SPADA 等[15]利用shotgun 蛋白质组学分析发现驴乳中pH 4.6 不溶部分含有大量的L-氨基酸氧化酶,能催化氨基酸氧化脱氨生成酮酸、氨和H2O2,是驴乳中较为丰富的酶之一。此外,该酶与溶菌酶、乳铁蛋白和免疫球蛋白结合,与驴乳的抗菌潜力密切相关。NGUYEN 等[16]利用无标记相对定量液相色谱串联质谱技术分析水牛和奶牛的乳脂肪球膜蛋白,发现相同质量浓度的乳脂肪球膜蛋白中,水牛乳中的XO浓度是奶牛乳的6 倍;通过蛋白质印迹法和蛋白质在MFGM 表面的分布证实了XO 的表达会影响乳的风味和保质期。
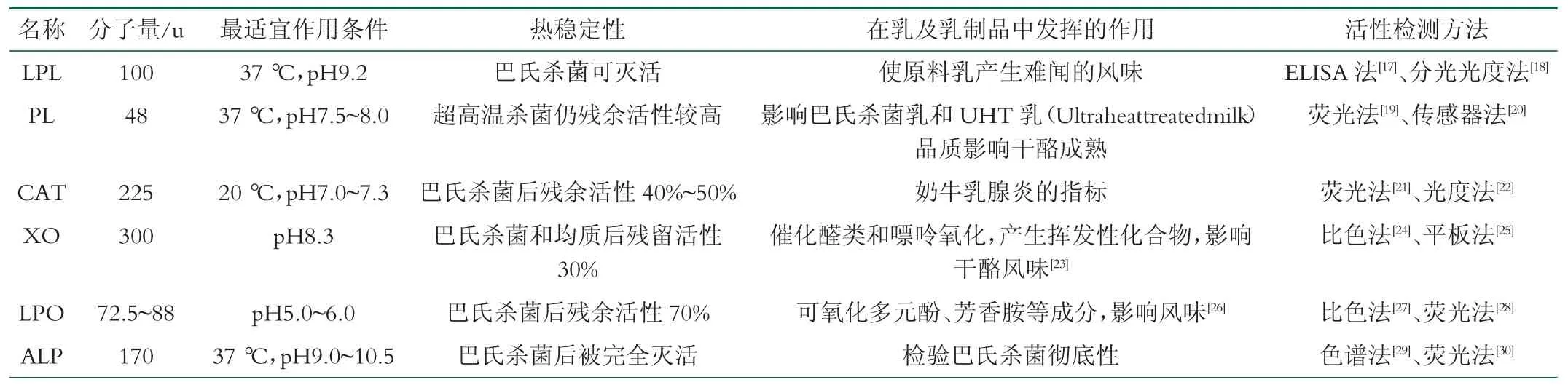
表1 乳及乳制品中主要内源酶的特性
1.2 乳中微生物产生的酶
原料乳在挤出后,易被乳房表面、挤奶设备、空气、水、饲料、土壤等多种污染源污染[31]。低温贮藏(4~6 ℃)可以减缓但无法完全抑制乳中微生物,特别是假单胞菌(Pseudomonas)、不动杆菌(Acinetobacter)、芽孢杆菌(Bacillus)和沙雷氏菌(Serratia)等嗜冷微生物的生长[32-33]。它们产生的脂肪酶十分耐热,150 ℃时D 值(在一定的处理环境中和在一定的热力致死温度条件下某细菌数群中每杀死90%原有残存活菌数时所需要的时间)可以达到290 s[34],这些酶能够直接水解乳脂肪小球,即使水解程度很小(1%~2%),也能使乳制品在货架期内发生水解酸败,出现乳脂分离、脂肪上浮等不利变化[35-36]。嗜冷微生物产生耐热蛋白酶(140 ℃条件下,D 值达到300 s)[37]会导致UHT 乳出现质量缺陷,包括黏度增加、形成沉淀、老化凝胶化、脂肪分离和产生苦味等[38]。有研究表明不同微生物来源的蛋白酶水解酪蛋白的顺序有所差异:来源于假单胞菌的蛋白酶AprX、蜡样芽孢杆菌的蛋白酶HhoA 水解酪蛋白顺序相同,依次为κ-CN、β-CN、αs-CN[34,39];而产自液化沙雷氏菌的蛋白酶Ser 2 水解酪蛋白顺序为β-CN、αS1-CN、κ-CN、αS2-CN[40]。
除嗜冷微生物产生耐热脂肪酶和耐热蛋白酶之外,在传统乳制品发酵过程中存在的乳酸菌和链球菌等微生物也能够产生酶。常见的有蛋白酶、脂肪酶和β-半乳糖苷酶,这些酶使乳制品产生独特的风味[41-42]。其中β-半乳糖苷酶(β-galactosidase,EC 3.1.2.23)能够催化乳糖水解生成葡萄糖和半乳糖,也被称为乳糖酶。曹永强等[43]基于一株从奶豆腐中分离得到的嗜热链球菌,探究了嗜热链球菌产乳糖酶的主要影响因素,并确定了乳糖酶的酶学特性(酶活性与温度成正比,最适温度为30~40 ℃,最适pH 值为6;锰离子对酶的激活作用最强,铜离子对酶具有较强的抑制作用)。乳糖酶可用于降低乳糖含量,生产无乳糖原料乳和低聚半乳糖[44-45]。
1.3 酶制剂
人为添加至乳制品中的酶制剂来源广泛,植物、动物和微生物均能生产[34,36]。脂肪酶作用于脂肪产生FFAs,被广泛应用于改善干酪风味、缩短干酪成熟时间[46]。在全球酶制剂市场,蛋白酶是市场份额最大的一类[2]。凝乳酶(Chymosin,EC 3.4.23.4)作为一种重要的蛋白酶制剂,最早从犊牛皱胃中提取得到,能使牛乳中κ-CN 的Phe105 与Met106 之间的肽键断裂,能改善干酪组织结构并提高干酪风味和营养价值及凝固性发酵乳的功能特性、组织状态、凝胶强度和物理稳定性[47-48]。目前国际上使用最广泛的微生物凝乳剂产自米黑根毛霉[49]。我国凝乳酶是近年来工业化生产的酶制剂来源于微小毛霉[50]。乳糖酶同时也是一类常见的酶制剂,用在发酵乳生产中可以提高其黏度,形成独特风味,提高发酵效率,延长保质期,用在干酪加工生产中可以缩短凝乳时间[47]。谷氨酰胺转氨酶(Transglutaminase,EC 2.3.2.13)是一类催化蛋白质(或多肽)之间的酰基发生转移,导致蛋白质(或多肽)之间发生共价交联聚合从而增强蛋白质凝胶网络结构,达到改变蛋白质的理化性质的酶[51],该酶常被用于干酪、酸乳等发酵乳制品生产中,发挥提高干酪产量或改善凝乳硬度的作用[52]。过氧化氢酶作为新型的酶制剂常用于去除乳及乳制品在紫外线照射过程中产生的H2O2异味[53-54]。CAT 在哺乳动物的肝脏与红细胞中含量较高,因此研究者起初从动物肝脏及血液中提取该酶[55-56]。随着微生物技术的发展,目前利用发酵法进行工业化生产从而获得CAT 已成为主流[57]。
2 酶对乳及乳制品关键品质的影响
2.1 原料乳
原料乳无法直接食用,在投入生产前会冷却至4~6 ℃保藏,但在酶的作用下仍易发生脂肪上浮、乳成分变化等品质劣变。在内源酶以及微生物产生的外源酶的作用下,4 ℃下冷藏3 d 后的原料乳成分发生了显著变化(P<0.05),其中脂肪含量增加0.01 g/100 g,蛋白质含量下降0.03 g/100 g,乳糖含量增加0.13 g/100 g[58]。GLANTZ 等[59]采用傅里叶变换红外光谱(Fourier transform infrared spectrometry,FT-IR)技术,经关联性分析结果显示牧场中的脂肪酶活性与Ca2+和谷氨酸-β-CN 结合物呈正相关,与十八碳三烯酸、细菌总数和β-乳球蛋白A 呈负相关,蛋白酶活性与总蛋白、αS2-CN、αS1-CN 片段(50-180)和β-CN 片段(108-209)呈正相关,与碳十六酸和αS1-CN 呈负相关;乳品加工厂中的脂肪酶活性与原料乳的pH、乳糖含量呈负相关,与Ca2+、十八碳烯酸、细菌总数呈正相关;蛋白酶活性与原料乳中SCC、β-乳球蛋白B 和β-CN片段(108-209)呈正相关;YUAN 等[60-61]利用随机扩增多态DNA 分析和16S rRNA 测序技术,在中国11 个城市共16 个牧场的4 ℃原料奶中分离得到76 株嗜冷菌,共涵盖24 个菌属,研究这些嗜冷菌在不同贮藏温度(7、28 ℃)下的腐败能力发现不动杆菌属和假单胞菌属可产生高活性脂肪酶;耶尔森菌属、假单胞菌属、沙雷菌属和金杆菌属可产生蛋白酶并表现出较高的蛋白水解活性;部分嗜冷菌菌株可产生β-D-半乳糖苷酶和磷脂酶。将嗜冷菌(假单胞菌属和沙雷氏菌属)接种于乳中并在不同温度下储存20 d,经顶空固相微萃取(Head space solid phase micro extraction,HSSPME)和气相色谱-质谱联用(Gas chromatography mass spectrometry,GC-MS),研究者在变质乳中检测出47 种挥发性有机化合物,其中包括丁酸、己酸和辛酸等[62],丁酸和己酸具有强烈的令人不快的味道,如腐臭味、苦味、不洁味、涩味,辛酸则具有肥皂味[63]。本课题组对原料乳4 ℃冷藏期间微生物变化及功能注释进行宏基因组学分析,同时对各冷藏阶段脂类代谢物的差异及变化进行分析[64-65]。结果表明,不动杆菌属及假单胞菌属在冷藏中后期丰度较高,KEGG 通路分析显示该阶段新增游离脂肪酸表现活跃且主要参与了脂肪酸生物合成、不饱和脂肪酸生物合成及亚油酸代谢。不动杆菌(Acinetobacter)和假单胞菌属(Pseudomonas)皆可以产生蛋白酶和脂肪酶,脂肪酶作用使乳黏度逐渐增强,蛋白酶溶解乳中部分酪蛋白,导致胶束稳定性破坏、苦肽形成及氨基酸分解,最终乳组织出现乳清蛋白沉淀、凝块等现象并产生苦味物质[66-67]。
2.2 巴氏杀菌乳
巴氏杀菌乳(Pasteurized milk)的特点是比较完好地保留了营养物质和风味,但低强度热处理(63 ℃,30 min;72 ℃,15 s)并不能杀死/灭活原料乳中所有的微生物和酶,稍有不慎就会腐败变质。FERREIRA等[68]利用FT-IR 技术发现荧光假单胞菌所产生脂肪酶诱导巴氏杀菌乳脂解后,脂肪和蛋白质含量发生了显著变化。基于高通量测序和相关性分析手段,DING等[69]发现巴氏杀菌乳中的优势菌种,即假单胞菌(Pseudomonas)、棒状杆菌(Corynebacterium)、链球菌(Streptococcaceae)和芽孢杆菌(Bacillus)等,与挥发性有机化合物(Volatile organic compounds,VOCs)的产生具有显著相关性。ZIYAINA 等[70]研究发现随着贮藏时间的延长,巴氏杀菌乳中细菌数量的增加、酶活性的增强,当蛋白酶和脂肪酶达到一定活性时,开始形成VOCs,不断增加的微生物水平及其释放的酶可直接影响巴氏杀菌乳风味。该结果为使用风味敏感的化学传感器监测巴氏杀菌乳质量提供了基础。此外,最近的研究结果表明外源添加脂肪酶对巴氏杀菌后脱脂重组乳的风味、结构性质和感官特性方面起着重要影响。ZHANG 等[71]研究发现脂肪酶可以催化巴氏杀菌全脂乳和奶油产生大量的VOCs,尤其是短链脂肪酸和脂肪酸乙酯。感官评分表明,添加脂肪酶后的重组乳口感更醇厚,这可能是由于脂肪酶催化脂肪酸、甘油酯和醇之间的(反式)酯化反应,产生乳化增稠效果。CHI等[72]采用HS-SPME 和电子鼻技术区分不同脱脂乳制品中的挥发性风味成分,与巴氏杀菌脱脂乳和UHT脱脂乳相比,重组乳中的VOCs 含量显著增加,并且检测到26 种全新的VOCs(包含有酸类、酯类、酮类等),表明脂肪酶催化反应导致了重组乳的风味增强。而从非挥发性的角度来看,ZHANG 等[73]研究发现脂肪酶催化后,重组乳的乳脂滴有相互聚集的趋势,使整个乳体系的粒径分布和微观结构变得多样化,并影响了乳体系稳定性的变化趋势。
2.3 UHT 乳
UHT 乳经超高温瞬时灭菌后,UHT 乳中的少量耐热酶(PL 以及嗜冷微生物产生的蛋白酶如AprX)会有20%~40%的残存活性,不断水解乳蛋白导致UHT 乳在货架期内出现质量缺陷[74-75]。研究表明,添加了PG 和低水平PL 的UHT 乳比未添加同类物质的牛乳形成凝胶更快[76],且PL 能够促进分子间形成β-折叠,导致蛋白沉积[77]。AprX 残留活性会导致UHT乳在货架期内蛋白质水解程度提高,非蛋白氮和非酪蛋白氮含量显著增加[78-79]。STOECKEL 等[80]研究发现,利用添加了不同假单胞菌菌株的原料乳所生产的UHT 乳中,UHT 乳发生质量缺陷的顺序为:出现苦味、形成颗粒、脂肪乳化、产生沉淀、开始凝胶化,缺陷的发生与蛋白酶活性线性相关,但凝胶化只发生在酶活较高时期。进一步,研究者曾比较探讨了PL 与AprX 对UHT 乳品质影响的差异。PL(0.8 mg/L)可导致脱脂UHT 乳中60%以上的β-CN 水解,水解度达到2.1%时形成软凝胶;AprX(10 mg/L)造成UHT乳中80%的κ-CN 水解,20%的αS1-CN 水解,15%的β-CN 水解,水解度超过1.3%时形成强凝胶[81]。高水平的PL 或AprX 均可导致UHT 乳中脂肪不稳定,但形成机理有所不同,前者通过诱导脂肪小球合并导致脂肪上浮,后者引起脂肪小球絮凝而导致发生沉积[82]。
2.4 干酪
干酪中的酶与干酪品质的形成息息相关,干酪风味一般通过加入凝乳酶或者谷氨酰胺转氨酶对干酪凝乳后进行蛋白水解和脂解来获得的。KENDIRCI等[83]发现添加脂肪酶的白奶酪FFAs 浓度和VOCs 丰度与未添加的白奶酪相比有显著提升,并形成了8 种新的挥发性物质。RANI 等[84]发现瑞士干酪经龙舌兰芽孢杆菌PR13 所产脂肪酶(200 U)处理后,风味更受大众欢迎,且成熟期缩短了1 个月。HERNAWDEZ等[85]研究了胃前脂肪酶对Idiazabal 干酪成分和感官特性的影响。结果表明,随脂肪酶添加量增加,干酪中FFAs 的含量先增加后降低;脂肪酶对Idiazabal 干酪的气味强度、丁酸味或凝乳味等特性有着积极的影响。除上述脂肪酶对干酪的影响之外,干酪中同时存在的蛋白酶,也在不同程度上影响干酪的风味、质构和凝乳效率。萨仁高娃等[86]研究发现添加PL 可增加干酪中可溶性氮、游离氨基酸和FFAs 含量,且对干酪质构有积极影响:随贮藏时间延长,干酪硬度降低,咀嚼性变差,而黏着性增强;酪蛋白结构愈发致密,空穴小且少,干酪成熟所需时间变短。除PL 外,外源添加酶制剂对干酪品质的影响同样有不少研究。徐辉艳[87-88]发现在制作干酪过程中,添加木瓜蛋白酶可使蛋白质降解更彻底,促进游离氨基酸形成,形成大小不一的孔洞,使致密结构变得松散、柔软且有弹性,随着木瓜蛋白酶添加量的增加,干酪的硬度和咀嚼性等质构参数呈下降趋势。此外,谷氨酰胺转氨酶用于干酪生产,通过强化蛋白网络提高了干酪的硬度、蛋白含量及产量[89-90]。
2.5 酸奶
酸奶是一种具有特殊感官特性的发酵乳制品,由于营养健康价值和良好风味深受消费者青睐。酸奶风味的主要贡献者是脂肪酶,它可以水解脂肪产生脂肪酸,因此添加脂肪酶改善酸奶风味受到了不少研究者的关注。HUANG 等[91]采用扫描电镜、高效液相色谱(High performance liquid chromatography,HPLC)和GC-MS 技术对混合乳酸菌发酵酸奶进行研究,发现脂肪酶的添加显著提高了产品中与甜味和苦味相关氨基酸的含量,并造成乳球表面粗糙,网状结构更加紧密。白雪[92]、杨爽等[93]利用电子鼻结合GC-MS 及HS-SPME 技术,对外源脂肪酶处理酸奶前后的VOCs 进行鉴定与比较。结果发现,脂肪酶处理显著增加了酸奶中醛酮类、酯类、杂环和芳香族类化合物的相对含量,风味得到改善。上述研究为开发酸奶风味补充剂提供了新的见解,具有良好的应用潜力。此外,谷氨酰胺转氨酶处理也能有效改善酸奶的感官及食用品质,使胶粒分布更加均匀,网络结构更加致密,感官评分更高[94-95]。
2.6 其他发酵乳制品
除牛奶外,其他畜种的原料乳或乳粉经杀菌、发酵后形成的pH 降低的乳制品统称为其他发酵乳制品。此类乳制品中涉及多种微生物共栖生长,向其中添加凝乳酶等酶制剂能够起到改善感官特性、缩短发酵时间和延长保质期的作用[96]。制备酸驴乳时添加凝乳酶和谷氨酰胺转氨酶,能提高产品凝乳硬度、减少凝乳时间[97]。添加谷氨酰胺转氨酶生产凝固型发酵酸羊乳可显著提升产品的黏性、硬度、持水性等参数,减少凝乳时间,提高感官特性,电镜结果表明该酸羊乳形成了更为致密的三维蛋白网状结构[52,97]。此外,IBRAHIM等[98]研究表明发酵时添加乳糖酶相较于其他处理(不添加乳糖酶或者发酵前添加乳糖酶),生产得到的发酵驼乳中胞外多糖含量最高,表观黏度增加,感官特性评分显著提升。
3 结论与展望
酶作为一种生物活性物质,在乳及乳制品的生产和贮存中起着重要作用。内源酶和污染微生物产生的外源酶通常会对乳品质造成消极作用,改变乳的成分性质进而造成质量和安全问题;而人工添加的酶制剂常常起到积极影响,在改善了乳制品品质的同时,起到降低生产成本、延长保质期的作用。
得益于近代生物技术和分析方法的进步,研究思路逐渐成熟,研究成果日趋完善。然而乳及乳制品的贮藏与加工是一个复杂的工程,研究面临的难题依然很多,如何从本质上解释由酶诱导的发生在货架期内的乳制品品质劣变;如何平衡内、外源酶在乳制品中的水解程度;酶制剂如何打破技术壁垒、提高应用技术和扩大应用范围等问题仍然是乳品科学和乳品产业发展的难题。因此,研究酶对乳及乳制品品质变化的本质、探究酶引起乳及乳制品品质变化的机制,控制酶影响因素及作用水平,推动酶制剂在乳及乳制品中的运用,将有助于乳及乳制品科学的发展,促进我国乳及乳制品贮藏与加工产业的持续发展。
