龙葵腺毛发育的细胞学研究
2023-12-05马璐璐陈佳琪金晓霞岳中辉
马璐璐,刘 昭,陈佳琪,李 伟,金晓霞,岳中辉
(哈尔滨师范大学 生命科学与技术学院,黑龙江省植物学省级重点实验室,哈尔滨 150025)
腺毛(glandular trichomes)是植物在长期进化过程中为了应对生物和非生物环境胁迫而演化出来的特殊适应性结构,广泛分布于陆地植物(包括裸子植物、被子植物、苔藓植物)地上部分的表面,是植物重要的分类学性状之一[1]。腺毛可合成、分泌、储存多种天然化合物[2-3],既是“细胞化学工厂”[如黄花蒿(Artemisiaannua)腺毛合成青蒿素[4]],又是植物防御器[如烟草(Nicotianatabacum)腺毛分泌物可抵御蚜虫[5]]。此外,植物可通过腺毛储存隔离毒物,如蜈蚣凤尾蕨(Pterisvittata)将重金属砷积累于腺毛中以降低胁迫[6]。
龙葵(SolanumnigrumL.)为茄科茄属一年至多年生草本植物,分布广泛,中国有3个种及1个变种[7]。全草入药,具清热消肿、止咳平喘、抗菌、抗氧化、抗肿瘤等作用[8-9]。龙葵茎叶表面腺毛丰富,张海洋等[10]对其腺毛形态进行了描述,但关于龙葵腺毛发育、成分及功能却鲜有报道。龙葵为镉超富集植物,可参与土壤修复[11-12],研究表明龙葵可将镉富集于叶片与茎的细胞壁及液泡中[13-14],然其腺毛是否参与镉胁迫的解除目前尚不清楚。本研究通过光学显微镜、扫描电镜、透射电镜技术对龙葵腺毛的类型、起源、生长、成熟、分泌、衰老等发育过程的细胞学特性进行了观察,并结合组织化学分析,揭示龙葵腺毛成分及分布,为龙葵进一步开发与利用提供参考。
1 材料和方法
1.1 材 料
试验材料为龙葵叶片和茎。龙葵种子由黑龙江省普通高等学校植物生物学重点实验室提供。2022年,温室培养[温度25 ℃/18 ℃,相对湿度(70±5)%]。待幼苗长至四叶一心时期取叶片边缘、叶片中部、叶脉及茎进行试验。
1.2 试验方法
1.2.1 龙葵腺毛的光学显微观察
用蒸馏水清洗龙葵叶片和茎表面,分别在叶片不同位置(叶片边缘、叶片中部、叶脉)切取边长为1 cm的小方块,切取茎表面宽度1 mm,长度1 cm大小,制备临时水装片,光学显微镜(Nikon Eclipse E600w)下观察腺毛的类型、分布及发育过程并拍照。
1.2.2 龙葵腺毛的扫描电镜观察
使用双面刀片切取新鲜叶片不同位置(长度5 mm,宽度3 mm)和茎(长度5 mm,宽度3 mm),粘贴在样品台上,利用扫描电镜(日立SU8010)的低温冷冻制样及传输技术(Cryo-SEM)实现腺毛发育各阶段,尤其是分泌活动的快速有效观察。避免传统固定、化学干燥等处理对试验结果造成的影响。
1.2.3 龙葵腺毛的透射电镜观察
使用双面刀片切取新鲜叶片不同位置(宽度1 mm,长度3 mm)和茎(宽度1 mm,长度1 cm),采用常规超薄切片技术,戊二醛(3%)、锇酸(1%)双固定,磷酸缓冲液冲洗,Epon812包埋,超薄切片机(Leica-UC6)切片(70 nm),柠檬酸铅与醋酸双氧铀染色,透射电子显微镜(Hitachi HT7800)观察腺毛超微结构及其分泌特征并拍照。
1.2.4 龙葵腺毛的组织化学染色观察
如1.2.1节所述切取试验材料并置于载玻片上,分别滴加组织化学染色剂进行染色,染色后用蒸馏水冲洗去浮色,制成临时装片,在光学显微镜(Nikon Eclipse E600w)下观察,通过不同颜色判定腺毛中成分类型及分布情况,通过颜色深浅判断成分含量高低(图版Ⅴ,a-g,i-j)。组织化学染色剂显色反应:Nadi试剂通过半缩醛羟基可与萜类物质中的羰基反应,形成蓝色沉淀[15],Wagner试剂使生物碱呈现棕黄色,苏丹黑显示总脂质,尼罗蓝可使中性脂质呈现红色,酸性脂质呈现蓝色,Lugol试剂用以检测蛋白质呈黄色,甲苯胺蓝O用以检测酚类呈蓝色[16],PAS反应显示中性多糖呈红色[17],钌红显示酸性多糖呈淡红色[18]。其中,类黄酮成分可自发荧光,在荧光显微镜下(UV激发)[19]观察并拍照。
2 结果与分析
龙葵茎与叶表面密生腺毛(图版Ⅰ,a-c),可分为两大类:单细胞头腺毛,即腺毛头部为单个细胞,主要分布于茎表面及叶的上下表面(图版 Ⅰ,b、d);多细胞头腺毛,即腺毛头部由多个细胞组成(图版 Ⅰ, e),主要分布于茎的单细胞头之间及叶脉、叶边缘。
2.1 龙葵腺毛的发育
龙葵腺毛由表皮细胞发育而来:表皮细胞局部突起为腺毛原始细胞(图版 Ⅱ,a1),经平周分裂,形成1个基细胞与1个顶细胞(图版 Ⅱ,a2);顶细胞继续平周分裂,形成1个柄细胞与1个原始头细胞(图版 Ⅱ,a3)。
2.1.1 单细胞头腺毛
当原始头细胞膨大,直接发育为成熟头细胞,则与纵向延长的柄细胞共同构成具1个柄细胞的单细胞头腺毛(图版Ⅱ, a4)。
当原始头细胞不膨大,而平周分裂形成2个子细胞:1个子细胞位于顶端,获得细胞质较多,可膨大发育为头细胞;另1个子细胞获得细胞质较少,发育为新的柄细胞,与原柄细胞共同构成具2个柄细胞的单细胞头腺毛(图版Ⅱ,a5-a6)。
当位于顶端的子细胞不膨大(图版Ⅱ,a5),而是充当新的原始头细胞,继续平周分裂产生新的头细胞与柄细胞(图版Ⅱ,a7),新的头细胞膨大,柄细胞伸长,可与原有的2个柄细胞共同构成具3个柄细胞的单细胞头腺毛(图版Ⅱ,a8);同理,若新的头细胞继续分裂,则形成具4个柄细胞的单细胞头腺毛(图版Ⅱ,a9-a10)。
可见,单细胞头腺毛的发育方式为顶端生长,即新柄细胞均来自于头细胞的分裂,而非旧柄细胞的分裂。基细胞形成后则不再分裂,直接分化成腺毛基部。
随着单细胞头腺毛的成熟,头细胞直径不断增加,呈淡黄绿色(图版Ⅱ,a11);柄细胞逐渐伸长,无色。分泌物由头细胞产生,逐渐积累并充满皮下空间(图版Ⅱ,a12),可通过顶端孔隙释放出来(图版Ⅱ,a13-a15)。分泌物量大且粘度高,通常在头与柄细胞表面形成局部的厚层(图版Ⅱ,a16)。分泌完成后,腺毛头细胞皱缩、凹陷,柄细胞随之退化,整个腺毛趋向衰老死亡(图版Ⅱ,a17)。
2.1.2 多细胞头腺毛
腺毛原始细胞经平周分裂,形成1个基细胞与1个顶细胞,基细胞不再分裂,直接分化为腺毛基部;顶细胞直径逐渐增加,形成原始头细胞(图版 Ⅱ,b1、b2)。根据原始头细胞分裂次数与分裂方向不同,可将多细胞头腺毛分为1层、2层与3层多细胞头腺毛。
1层多细胞头腺毛的发育:原始头细胞平周分裂1次,形成2个子细胞,位于顶端的细胞膨大为头细胞,另1个为颈细胞(图版Ⅱ,b3)。接着,头细胞垂周分裂2次,产生4个均等的子细胞排列为1层(图版Ⅱ, b4、b5)。头细胞积累分泌物于皮下空间,并分泌(图版Ⅱ,b6),分泌后皱缩(图版Ⅱ,b7)。另外,发现2种特殊类型的1层多细胞头腺毛:一种具2个柄细胞,且腺毛柄极长(图版Ⅱ,e1);另一种具2个颈细胞(图版Ⅱ,e2)。
2层多细胞头腺毛的发育:原始头细胞平周分裂1次,产生2个子细胞(图版Ⅱ,c1),位于顶端(上层)的子细胞再进行2次垂周分裂,形成同层4个细胞;下层的子细胞进行1次垂周分裂,形成2个细胞,因此,腺毛头部分两层,整体呈倒三角形并贴近表皮(图版Ⅱ,c2)。分泌物由贴近表皮一侧的头细胞产生,于皮下空间积累,并通过分泌孔释放,腺毛随后衰老(图版Ⅱ,c3、c4)。另外,发现1种特殊类型的2层多细胞头腺毛,即头细胞基部具有1个颈细胞(图版Ⅱ,e3)。
3层多细胞头腺毛的发育:原始头细胞平周分裂1次,形成2个子细胞,位于顶端的子细胞继续进行1次平周分裂,而另1个子细胞不分裂,形成3层头细胞(图版 Ⅱ,d1),每层细胞再分别进行1~2次垂周分裂,形成椭球形头部(图版 Ⅱ,d2)。腺毛成熟后,分泌物由最顶层且贴近表皮一侧的头细胞产生,由顶端分泌孔隙释放,随后腺毛衰老(图版 Ⅱ,d3、d4)。
2.2 龙葵腺毛超微结构特征
2.2.1 扫描电镜观察
龙葵发育成熟的腺毛具有分泌能力。单细胞头腺毛的分泌物在头细胞皮下空间积累,致使头细胞表面形成数个大小不等的透镜状突起(图版Ⅲ,a1-a3);随着分泌物积累增多,突起不断增大,头细胞表面现包块样结构(图版Ⅲ,a4);随后,包块表面逐渐呈棱状,并破口,分泌物被释放,角质层部分凹陷(图版Ⅲ,a5-a7);分泌物量大且粘滞性强,可在头细胞与柄细胞表面形成局部的厚层,甚至包裹头细胞(图版Ⅲ,a8、a9),与光学显微镜观察结果相吻合。
与单细胞头腺毛类似,在多细胞头腺毛成熟期,分泌物在皮下空间不断积累,头细胞表面及头细胞间出现多个透镜状突起(图版Ⅲ,a10)。3层多细胞头腺毛的分泌物由顶端靠近表皮细胞处释放,分泌物附着于表皮细胞(图版Ⅲ,a11、a12),随后腺毛皱缩衰老(图版Ⅲ,a13),与光学显微镜观察结果吻合。
2.2.2 透射电镜观察
发育成熟的3层多细胞头腺毛(图版Ⅳ,a1),头细胞的细胞壁较厚,且电子密度高,外被薄层角质层;头细胞间存在胞间连丝(图版Ⅳ,a3),头细胞中细胞质丰富(图版Ⅳ,a2),存在液泡、高尔基体、内质网等细胞器(图版Ⅳ,a3、a5),其中滑面内质网数目众多,高尔基体膜囊数多,且分泌活动活跃,形成大量含有嗜锇物质的囊泡,运输至细胞壁附近(图版Ⅳ,a6),与细胞质膜融合(图版Ⅳ,a7),进而将嗜锇物质转移至细胞壁,经细胞壁积累(图版Ⅳ,a8),呈弥散状扩散至细胞壁与角质层之间的皮下间隙(图版Ⅳ,a9)。随着嗜锇物质不断增多,皮下间隙增大(图版Ⅳ,a9),直至分泌物释放。
2.3 龙葵腺毛组织化学分析
以龙葵腺毛水装片为对照(图版Ⅴ,a1-a4),其组织化学染色结果显示,腺毛中存在萜类、生物碱、脂质(中性脂质与酸性脂质)、蛋白质和酚类(类黄酮)、多糖(中性多糖与酸性多糖)。具体为:4种腺毛中头细胞与柄细胞中均呈现蓝色(图版Ⅴ,b1-b4)和棕黄色(图版Ⅴ,c1-c4),说明存在大量萜类物质和生物碱;头细胞与柄细胞均呈现黑色(图版Ⅴ,d1-d4),说明其存在总脂质,头细胞中蓝色较柄细胞中略深(图版Ⅴ,e1-e4),说明头细胞中酸性脂质含量多于柄细胞;头细胞中分别呈现黄色(图版Ⅴ,f1-f4)、蓝色(图版Ⅴ,g1-g4)、绿色荧光(图版Ⅴ,h1-h4)、红色(图版Ⅴ,i1-i4),说明其中含大量蛋白质、酚类、类黄酮及中性多糖(纤维素),而柄细胞中较少,值得一提的是,3层多细胞头腺毛的皮下空间呈现红色(图版Ⅴ,i4),说明皮下空间含有中性多糖。酸性多糖主要集中于多细胞头腺毛的头部,呈淡红色,其含量较少,单细胞头腺毛呈现极少淡红色,说明其中含量极低(图版Ⅴ,j1-j4)。
3 讨 论
3.1 腺毛类型与发育方式
腺毛是茄科植物重要的分类学特征之一,番茄[20]有4种类型腺毛;烟草[21]有6种类型腺毛;马铃薯、茄子、胡椒、矮牵牛均具腺毛且形态类型多样[22-24]。本研究发现龙葵具2大类腺毛,即单细胞头腺毛(柄细胞1~4个)和多细胞头腺毛(1层、2层与3层多细胞头腺毛),与张海洋等[10]报道类似,但与其单细胞头腺毛的柄细胞数目2~5不一致。另外,张海洋等[10]认为在单细胞头腺毛的发育中,新的柄细胞由原柄细胞平周分裂而来,而本研究认为腺毛的生长方式属于顶端生长,新的柄细胞始终由顶端头细胞平周分裂而来。值得注意的是,笔者还发现3种特殊类型的多细胞头腺毛:具2个柄细胞的1层多细胞头腺毛、具2个颈细胞的1层多细胞头腺毛、具1个颈细胞的2层多细胞头腺毛,在茄科植物中尚未见报道。
3.2 腺毛分泌方式
植物腺毛分泌方式多样:(1)腺毛可能通过渗透释放分泌物,如蓝花鼠尾草(SalviafarinaceaBenth.)[25]、广藿香[Pogostemoncablin(Blanco) Benth.][26];(2)腺毛可通过头细胞顶端细胞壁或角质层微孔或小孔释放分泌物,如菊科山金车(ArnicachamissonisLess.)[27]、刺槐(RobiniaviscosaVent. var.hartwigii)[15]、葡萄属(VitisL.)[28]与牛至木属(Lippia)[29];(3)腺毛可将分泌物积累于头细胞角质层下的皮下空间,分泌物增多导致角质层突起、破裂,进而释放分泌物,如树莓(RubusidaeusL.)[30]、夏至草(MarrubiumvulgareL.)[31]、罗勒(OcimumbasilicumL.)[32]、迷迭香(RosmarinusofficinalisL.)[33]、决明属(Chamaecristadentata)[34]、多榔菊属(DoronicumL.)[35]。本研究通过光镜水装片及组织化学结果直观显示了龙葵腺毛分泌物储存于头细胞的皮下空间,呈透明带样结构,并释放。扫描电镜冷冻传输图像进一步显示腺毛分泌过程其头细胞表面变化:分泌物积累导致头细胞表面形成大小不等透镜样突起—突起呈包块状—包块出现棱状破口—分泌物释放,甚至包裹头细胞,与光镜观察结果吻合。结合透射电镜研究结果显示腺毛头细胞分泌活动:腺毛头细胞中内质网与高尔基体极为丰富,合成代谢及分泌活动活跃,产生大量包裹嗜锇物质的囊泡,囊泡被运输至细胞壁附近,并与细胞膜融合,进而将嗜锇物质转移至细胞壁并积累,随后储存在角质层下皮下空间,直至角质层破裂。综上,笔者认为龙葵腺毛分泌方式属上述第(3)类,与树莓等[25]相似。
3.3 腺毛功能
植物腺毛可合成、储存和分泌多种化合物,在植物与其他生物及周围环境的相互作用中至关重要。番茄(SolanumlycopersicumL.)腺毛分泌的粘性物质(多糖、生物碱、萜类以及黄酮类)可困住真菌小蚊蚋,降低小型节肢食草动物的灵活性[36];烟草(NicotianatabacumL.)短柄腺毛中的叶状蛋白可抑制叶表面寄生虫孢子的萌发,从而降低叶片被侵染的机率[37]。本研究通过组织化学与荧光显微法得出,龙葵腺毛中含有大量多糖、生物碱、萜类、酚类、脂类和蛋白质,具分泌能力,分泌物量大且粘度高,推测其在防御植食性动物与病原微生物方面有重要作用。然而其腺毛中的成分是否为其主要药用成分尚待进一步研究。
植物腺毛具有减毒功能。蜈蚣凤尾蕨腺毛可积累重金属砷,保护植物免受砷毒害[6]。龙葵作为一种重金属镉的超富集植物,可将镉富集于叶片与茎的细胞壁及液泡中[13-14],笔者推测腺毛可通过其富集及分泌活动缓解镉胁迫。

图版 Ⅰ 龙葵表皮毛的形态与分布a.龙葵植株幼苗;b.茎, 箭头示单细胞头腺毛;c.叶片;d.叶脉表皮, 箭头示单细胞头腺毛;e.叶边缘, 箭头示多细胞头腺毛。Plate Ⅰ Morphology and distribution of epidermal trichomes of S. nigrumFig.a. Solanum nigrum L. plant seedings; Fig.b. Stems, arrows show unicellular head glandular trichomes; Fig.c. Leafblade; Fig.d. Vein epidermis, arrow shows unicellular head glandular trichomes; Fig.e. Leaf margin, arrow shows multicellular head glandular trichomes.

图版Ⅱ 龙葵腺毛的发育a1.表皮细胞突起为腺毛原始细胞;a2.箭头示基细胞与顶细胞;a3.基细胞、柄细胞与原始头细胞, 箭头示原始头细胞;a4.原始头细胞膨大, 形成具1个柄细胞的单细胞头腺毛;a5.原始头细胞分裂形成2个子细胞;a6.顶端子细胞膨大, 形成具2个柄细胞的单细胞头腺毛;a7.顶端子细胞分裂, 产生新的头细胞与柄细胞;a8.具3个柄细胞的单细胞头腺毛;a9.头细胞继续分裂;a10.具4个柄细胞的单细胞腺毛;a11.成熟的头细胞;a12.分泌物在皮下空间积累, 箭头示皮下空间;a13-a16.分泌物释放, 箭头示分泌物;a17.腺毛衰老。b1-b2.腺毛原始细胞平周分裂产生基细胞与顶细胞;b3.箭头示颈细胞;b4、b5、b5’.头细胞垂周分裂2次, 形成1层多细胞头腺毛;b6、b7、b7’.头细胞积累分泌物于皮下空间并分泌, 箭头示分泌物;c1.原始头细胞平周分裂1次;c2.2层多细胞头腺毛;c3-c4.分泌物积累于皮下空间并释放, 箭头示分泌物;d1.3层多细胞头腺毛初期;d2.每层头细胞进行1~2次垂周分裂;d3-d4.分泌物积累与释放, 箭头示分泌物;e1.具2个柄细胞的1层多细胞头腺毛, 箭头示2个柄细胞;e2.具2个颈细胞的1层多细胞头腺毛, 箭头示2个颈细胞;e3.具1个颈细胞的2层多细胞头腺毛, 箭头示颈细胞。Plate Ⅱ Development of glandular trichomes of S. nigrumFig.a1. Epidermal cell protuberances are glandular trichomes primordial cells; Fig.a2. Arrow shows basal cells and apical cells; Fig.a3. Basal cells, stalk cells and primitive head cells, primitive head cell is showed by arrows; Fig.a4. The primitive head cells enlarge to form glandular trichome of single cell head with a stalk cell; Fig.a5. Primitive head cell divides to form two daughter cells; Fig.a6. The apical daughter cells enlarged to form glandular trichome of single cell head with two stalk cells; Fig.a7. The apical daughter cells divide to produce new head cells and stalk cells; Fig.a8. Single-celled head glandular trichomes with three stalk cells; Fig.a9. Head cells continue to divide; Fig.a10. Single-celled head glandular trichomes with four stalk cells; Fig.a11. Mature head cells; Fig.a12. Secretions accumulate in subcutaneous space, and arrows show subcutaneous space; Fig.a13-a16. The secretion is released, and the arrow shows the secretion; Fig.a17. Senescence of glandular trichomes. Fig.b1-b2. The primitive cells of glandular trichome divide peripheral to produce basal cells and apical cells; Fig.b3. Arrow shows cervical cells; Fig.b4, b5 and b5’. Cephalic cells divide peritoneal twice to form a layer of multicellular head glandular trichomes; Fig.b6, b7 and b7’. Head cells accumulate secretions in subcutaneous space and secrete them, and arrows show secretions; Fig.c1. The primitive cephalic cells divid peripheral once; Fig.c2. Two-layer multicellular head glandular trichomes; Fig.c3-c4. Secretions accumulate in subcutaneous space and is released, and arrows show secretion; Fig.d1. Early stage of three-layer multicellular head glandular trichomes; Fig.d2. Each layer of head cells underwent 1-2 vertical peridivision; Fig.d3-d4. Accumulation and release of secretions, secretion is showed by arrows; Fig.e1. A layer of multicellular head glandular trichome with 2 stalk cells, the arrows show two stalk cells; Fig.e2. A layer of multicellular head glandular trichomes with two cervical cells, arrows showing two cervical cells; Fig.e3. Two layers of multicellular head glandular trichomes with one neck cell, and the cervical cells are indicated by arrows.
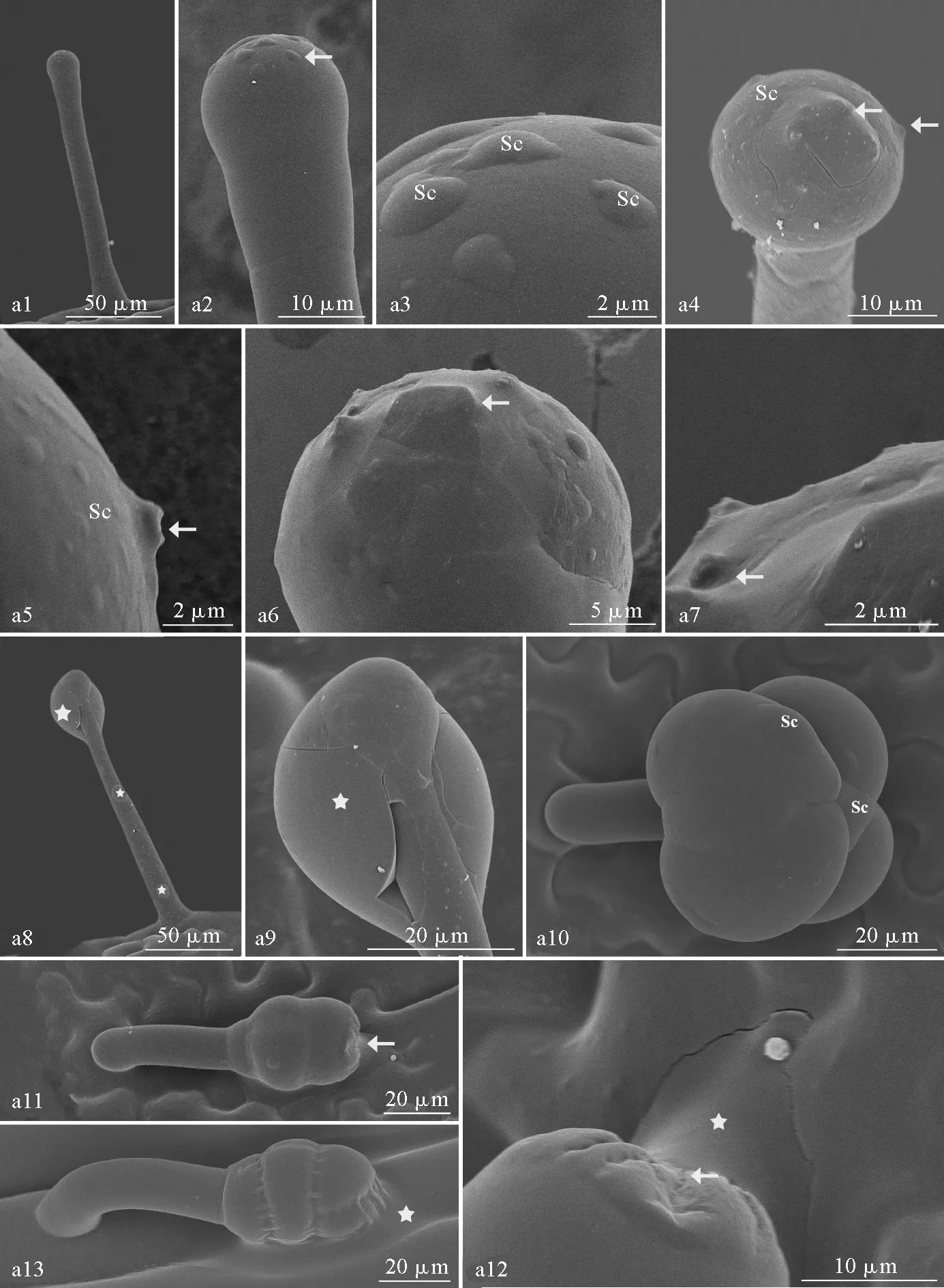
图版Ⅲ 龙葵腺毛超微结构(a)a1.单细胞头腺毛;a2-a3.头细胞表面透镜状突起(Sc);a4.头细胞表面包块样结构(箭头);a5.头细胞表面棱状突起、破口(箭头);a6.分泌物释放, 箭头示其他包块; a7.分泌后的破口(箭头); a8-a9.分泌物包裹头细胞(星号);a10.1层多细胞头腺毛, 头细胞表面透镜状突起(Sc);a11-a13.3层多细胞头腺毛释放分泌物(星号)。Sc.皮下间隙。Plate Ⅲ Ultrastructure of glandular trichomes of S. nigrum (a)Fig.a1. Glandular trichomes of single cell head; Fig.a2-a3. Lenticular process on the surface of cephalic cells(Sc); Fig.a4. Crouton-like structure on head cell surface (arrow); Fig.a5. Ridges and lacerations on the surface of head cells (arrow); Fig.a6. Secretion release, arrows show other packet blocks; Fig.a7. Post-secretory laceration (arrow); Fig.a8-a9. Secretions encased head cells (*); Fig.a10. A layer of multicellular head glandular trichomes, lenticular process on the surface of cephalic cells(Sc); Fig.a11-a13. Three-layer multicellular glandular hair discharge (*). Sc. Subcutaneous space.

图版Ⅴ 龙葵腺毛组织化学定位a1-a4.对照组;b1-b4.Nadi试剂检测萜类;c1-c4.Wagner试剂检测生物碱;d1-d4.苏丹黑检测总脂质;e1-e4.尼罗蓝检测酸性脂质与中性脂质;f1-f4.Lugol溶液检测蛋白质;g1-g4.甲苯胺蓝O检测酚类;h1-h4.类黄酮自发荧光;i1-i4.PAS检测中性多糖;j1-j4.钌红反应检测酸性多糖。Plate Ⅴ Histochemical localization of glandular trichome of S. nigrumFig.a1-a4. Control group; Fig.b1-b4. The terpenes were detected by Nadi reagent; Fig.c1-c4. Wagner reagent for the detection of alkaloids; Fig.d1-d4. Sudan Black assays total lipids; Fig.e1-e4. Nile blue detects acidic and neutral lipids; Fig.f1-f4. Lugol solution detects proteins; Fig.g1-g4. Toluidine blue O detection of phenols; Fig.h1-h4. Flavonoid autofluorescence; Fig.i1-i4. PAS reagent was used to detect neutral polysaccharides; Fig.j1-j4. Detection of acidic polysaccharides by ruthenium red reaction.
