基于全基因组测序的贝莱斯芽胞杆菌Ba-0321抑菌机制分析及相关功能验证
2023-10-21李小杰姚晨虓白静科陈玉国李淑君
李小杰,邱 睿,刘 畅,姚晨虓,白静科,陈玉国,李淑君
(河南省农业科学院烟草研究所/烟草行业黄淮烟区烟草病虫害绿色防控重点实验室,许昌 461000)
芽胞杆菌Bacillusspp.是一种嗜温、好氧、产芽胞的杆状细菌,能够产生耐热、耐旱、抗紫外线和有机溶剂的内生孢子、多种抗菌素和酶,具有广谱抑菌活性和极强的抗逆能力[1,2]。贝莱斯芽胞杆菌是芽胞杆菌中新发现的1 个种,普遍存在于自然界植物组织、土壤、河水以及海洋中[3-5]。有研究表明,贝莱斯芽胞杆菌具有防治多种植物病害的潜力,同时具有促生、固氮和抗逆境等多种优良功能,是重要的生防资源。郗良卿等[6]从健康甜樱桃中筛选出优质拮抗菌株——贝莱斯芽胞杆菌,能够抑制匍枝根霉Rhizopusstolonifer的生长,对樱桃软腐病也有良好的防治效果;沙月霞等[7]明确了水稻内生菌贝莱斯芽胞杆菌B.velezensisE69是一种潜在的、预防效果明显的生防菌株,具有预防稻瘟病兼防纹枯病等多种真菌病害的应用潜力;夏明聪等[8]明确了贝莱斯芽胞杆菌YB-145 是一株具有生防潜力的菌株,可用于小麦纹枯病的防控,同时能提高小麦根鲜质量、地上部鲜质量和地上部高度。
芽胞杆菌主要通过两种途径产生抑菌物质,一是通过核糖体途径合成小分子物质如肽类物质、细菌素、硫肽类物质(thiopeptide)、萜烯类物质(terpene)等[9,10],二是通过非核糖体途径合成的脂肽类化合物、聚酮类化合物和多肽类化合物等[11,12]。芽胞杆菌分泌的有益次生代谢产物种类繁多,但传统的试验分析和鉴定方法很难全面地分析其中的抑菌活性物质且耗时耗力,同时难以充分挖掘其相关基因,揭示其生防作用机制。全基因组测序为进一步认识和利用生防菌株提供了重要基础,尤其是随着测序技术的不断革新以及费用的大大降低,越来越多的芽胞杆菌基因组序列被公布。通过全基因组信息挖掘,研究者发现了大量已知或未知的抑菌物质合成基因簇。戴利铭等[13]对一株具有较强抑菌活性的内生解淀粉芽胞杆菌BS-3 进行全基因组测序分析,预测得到10 个次级代谢产物基因簇,其中包括6 类已知的具有抑菌活性的物质和4种功能未知的基因簇;高圣风等[14]从生防菌株B.velezensisZ 全基因组信息中发现编码次生代谢产物合成的基因簇13 个,其中有5 个未知功能基因簇。这些研究结果不仅有助于解析芽胞杆菌次级代谢产物的合成途径,同时能挖掘新的次级代谢产物,为新型生物药物的开发提供重要资源。
本课题组前期在健康烟株根际土壤中筛选到一株高效生防贝莱斯芽胞杆菌Ba-0321,本研究测定了菌株Ba-0321 无菌滤液对尖孢镰刀菌、疫霉菌等烟草主要病原真菌的抑制效果,同时利用第二代 illumina与第三代Nanopore 相结合的测序技术对该菌株进行全基因组测序,通过GO、COG、KEGG 和antiSMASH等数据库进行基因功能注释和次级代谢产物基因簇分析,并进行相关功能验证,以期为全面深入解析其抑菌机制、挖掘次级代谢产物基因资源和菌株的高效开发利用提供生物信息学基础。
1 材料与方法
1.1 试验材料
1.1.1 菌株 贝莱斯芽胞杆菌Ba-0321 分离自河南省许昌市烟田健康烟株根际土壤,现保藏于中国典型培养物保藏中心(保藏号CCTCCM2020440)。
植物病原真菌:烟草尖孢镰刀菌F.oxysporum、烟草疫霉菌P.nicotianae均分离自河南烟田发病烟株并保存于本实验室。
1.1.2 主要试剂和仪器 细菌基因组DNA 提取试剂盒,布鲁克(北京)科技有限公司;DL2000 DNA Marker,北京天根生化科技有限公司。PCR 仪,Applied Biosystems 公司;电泳仪,北京六一生物科技有限公司;NanoDrop 2000,赛默飞世尔科技公司。
1.1.3 所用培养基 贝莱斯芽胞杆菌Ba-0321 培养用LB 培养基:蛋白胨10.0 g,酵母提取物5.0 g,NaCl 10.0 g,蒸馏水1000 mL;尖孢镰刀菌培养用PDA 培养基:马铃薯200.0 g、葡萄糖20.0 g,蒸馏水1000 mL;疫霉菌培养用OA 培养基:燕麦片30 g,蒸馏水1000 mL。固体培养基均添加琼脂粉15 g/L。121 ℃高压灭菌20 min。
酪蛋白培养基:干酪素10.0 g,琼脂20.0 g,pH 7.2;纤维素水解培养基:蛋白胨10.0 g,KH2PO41.0 g,酵母浸粉10.0 g,NaCl 5.0 g,羧甲基纤维素钠 10.0 g,琼脂15.0 g,pH 7.2;几丁质培养基:胶体状几丁质10.0 g,KCl 0.2 g,NH4H2PO41.0 g,MgSO4·7H2O 0.2 g,琼脂 20.0 g,超纯水1000 mL。
1.2 方法
1.2.1 菌株Ba-0321 无菌滤液的抑菌能力测定 采用菌丝生长速率法,将菌株Ba-0321 接种于LB 液体培养基中,28 ℃、180 r/min 条件下恒温振荡培养3 d,5000 r/mim 离心10 min,取上清液用0.22 μmol/L 过滤器进行过滤除菌。分别按1%、5%和10%的比例将发酵滤液加入PDA 或OA 培养基后倒平板,将直径为5 mm 的尖孢镰刀菌和疫霉菌菌块分别置于上述含有不同发酵滤液的平板中间,以不加滤液处理为对照,每个处理重复3 次,25 ℃恒温箱中培养5 d,采用十字交叉法测量病原菌菌落直径并拍照,计算抑菌率,抑菌率(%)=(对照组菌落生长直径-处理组菌落生长直径)/对照组菌落生长直径×100。
1.2.2 菌株Ba-0321 全基因组测序 采用SDS 提取结合纯化柱(OMEGA)法,提取基因组DNA。利用Nanodrop、Qubit 和0.35%琼脂糖凝胶电泳进行纯度、浓度和完整性质检,然后进行文库构建。采用二代illumina 与三代Nanopore 相结合的测序技术进行测序。本次测序委托北京擎科生物科技有限公司完成。
1.2.3 基因功能注释 将预测得到的基因序列与GO(Gene Ontology)、COG(Cluster of Orthologous Groups of proteins)、KEGG(Kyoto Encyclopedia of Genes and Genomes)等数据库做 BLAST+(2.9.0+)比对,得到基因功能注释结果。
1.2.4 抑菌代谢产物分析 采用antiSMASH 程序(version 5.2.0)对菌株Ba-0321 中次级代谢产物合成基因簇进行预测。结合基因注释结果和NCBI 数据库BLAST 比对分析结果,挖掘菌株Ba-0321 可能具有抑菌活性的次级代谢产物及编码基因簇。
1.2.5 抗菌脂肽合成基因及防御机制相关基因的PCR 扩增 根据相似度高的已知芽胞杆菌基因组中Surfactin、Macrolactin、Fengycin、Difficidin、Bacillibactin、Bacillaene 和Bacilysin 脂肽抗生素合成基因(表1)及Ba-0321 菌株基因组中预测的防御机制相关基因(表2)设计引物,以Ba-0321 基因组DNA 为模板进行PCR扩增。PCR 反应体系:2×TaqPCR Mix 12.5 μL,10 μmol/L 正、反向引物各1 μL,基因组DNA 1 μL(50 ng)为模板,超纯水补足至25 μL。PCR 扩增条件为:95 ℃预变性 5 min;94 ℃变性 45 s,54 ℃退火40 s,72 ℃延伸50 s,30 个循环;72 ℃延伸8 min,4 ℃保存。PCR 反应结束后,取5 μL 扩增产物于1.5%的琼脂糖凝胶中电泳,在BIO-RAD 凝胶成像系统上观察、拍照。将获得预期大小的PCR 扩增产物送测序,测序所获得的序列在NCBI 中进行BLAST 比对分析。
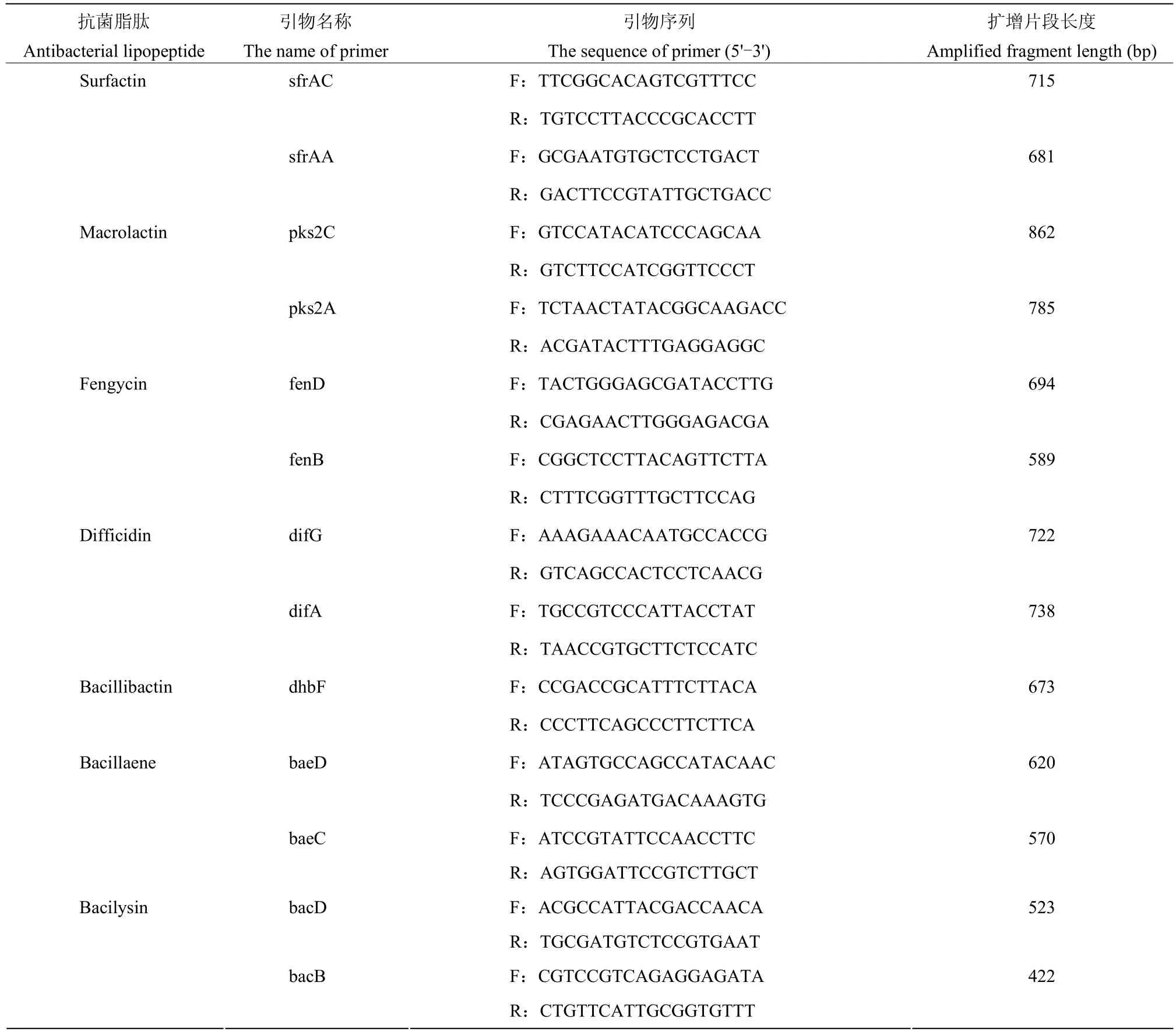
表1 抗菌脂肽合成基因的引物序列Table 1 Primer sequence of antibacterial lipopeptide synthesis gene
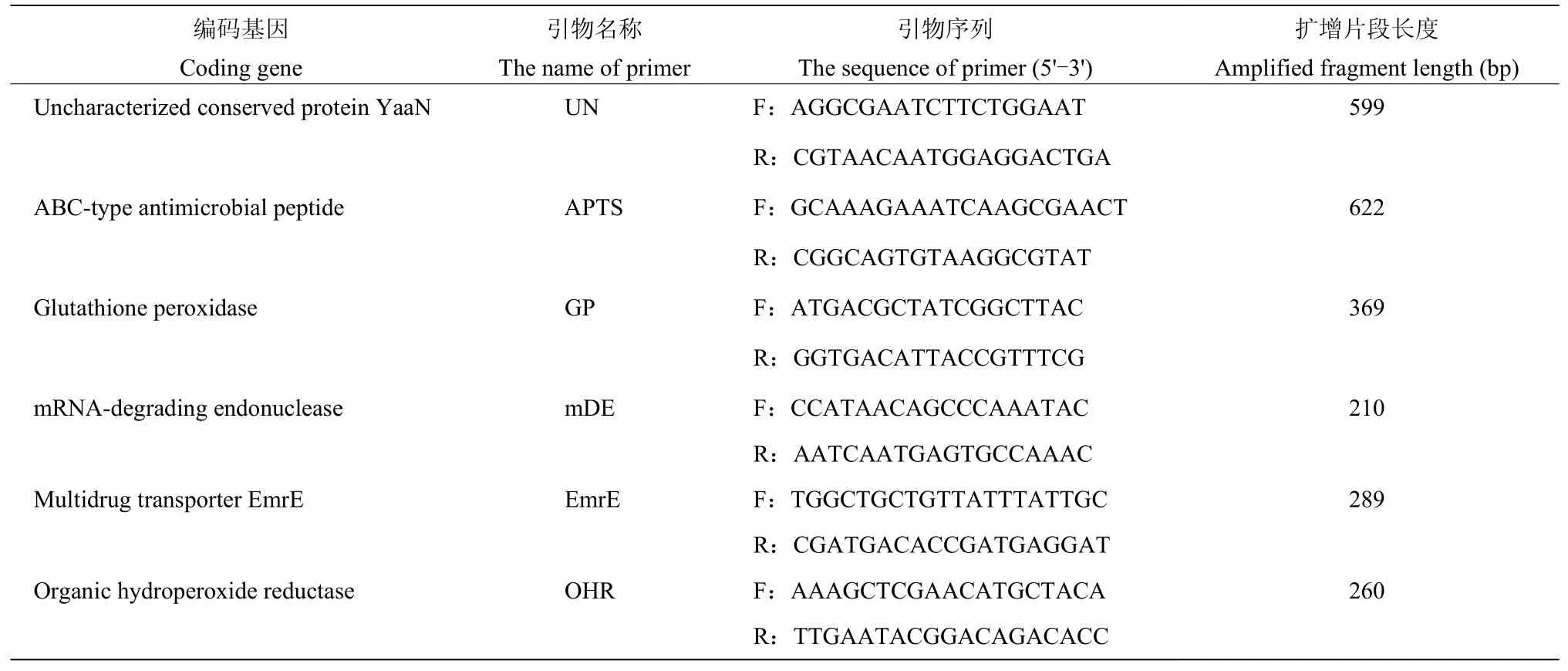
表2 防御机制相关基因的引物序列Table 2 Primer sequences of resistance related genes
1.2.6 菌株Ba-0321 产酶能力测定 将菌株Ba-0321 在LB 培养基上进行活化并划线培养,挑取单菌落接种于LB 液体培养基中28 ℃、160 r/min 振荡培养至OD600≈1.0,吸取5 μL 菌液分别点接于酪蛋白培养基、纤维素水解培养基和几丁质培养基上,28 ℃恒温培养3~5 d,观察上述培养基是否产生水解圈,以检测菌株是否具有蛋白酶、纤维素酶和几丁质酶活性。对照滴加等量LB 培养液,每个处理3 次重复。
2 结果与分析
2.1 贝莱斯芽胞杆菌Ba-0321 无菌滤液对烟草尖孢镰刀菌和疫霉菌的抑菌能力
试验结果表明,菌株Ba-0321 无菌发酵滤液对尖孢镰刀菌和疫霉菌均具有抑制效果,但不同添加量对病原菌的抑制效果不同。在相同体积的培养基中,无菌滤液所占比例越高,抑菌效果越明显。其中10%无菌滤液处理对两种病原菌菌丝生长的抑制率较高,分别可达57.38%和34.30%,极显著高于1%和5%无菌滤液处理。5%无菌滤液处理对尖孢镰刀菌菌丝生长的抑制率极显著高于1%无菌滤液的抑菌率;而5%无菌滤液处理对疫霉菌菌丝生长的抑制率与1%无菌滤液处理无显著差异。以上结果表明,菌株Ba-0321 能够产生具有抑菌活性的次生代谢产物,且抑菌活性与代谢产物的含量呈正相关(图1)。
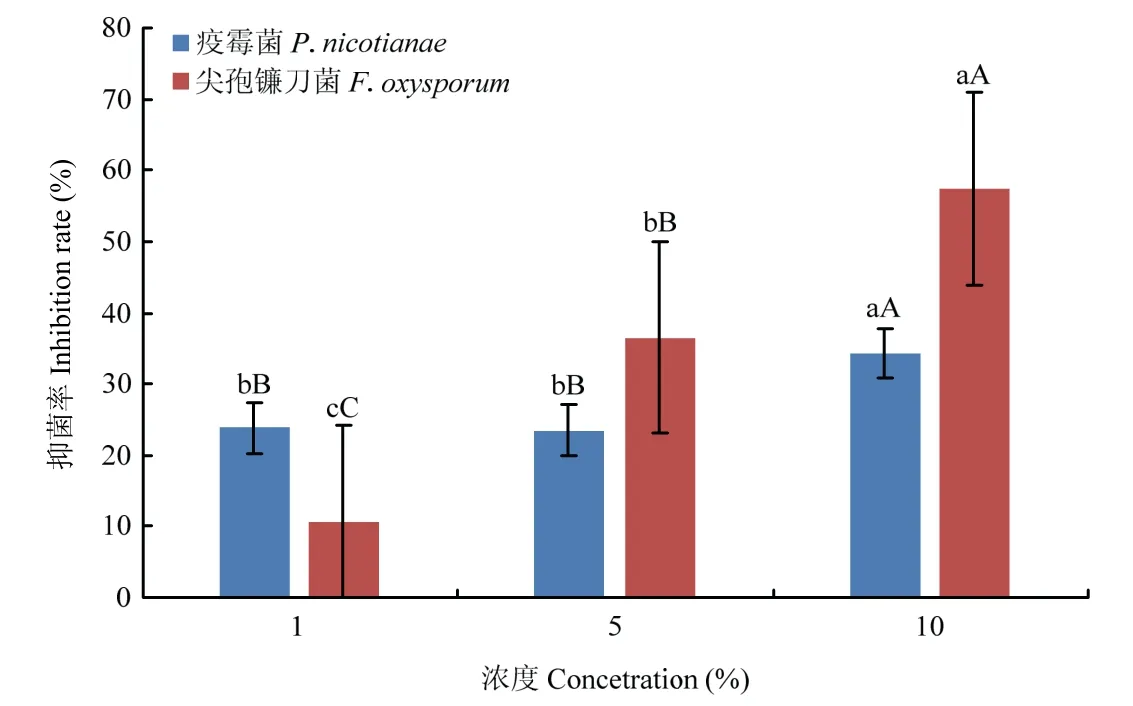
图1 菌株Ba-0321 无菌滤液不同添加量对烟草疫霉菌和尖孢镰刀菌的抑制效果Fig.1 Antifungal effects of strain Ba-0321 against P.nicotianae and F.oxysporum
2.2 贝莱斯芽胞杆菌Ba-0321 全基因组组成
菌株Ba-0321 全基因组大小为4099109 bp,平均GC 含量为46.31%;含有基因4103 个,其中编码基因3897 个,平均长度932 bp;含有tRNA 基因86 个,23S rRNA、16S rRNA 和5S rRNA 各9 个,tmRNA基因1 个,miscRNA 92 个。预测出基因岛10 个;前噬菌体2 个;重复序列202 个,覆盖率为0.45%。Ba-0321基因组测序数据提交至GenBank,登录号为CP101904,基因组圈图见图2。
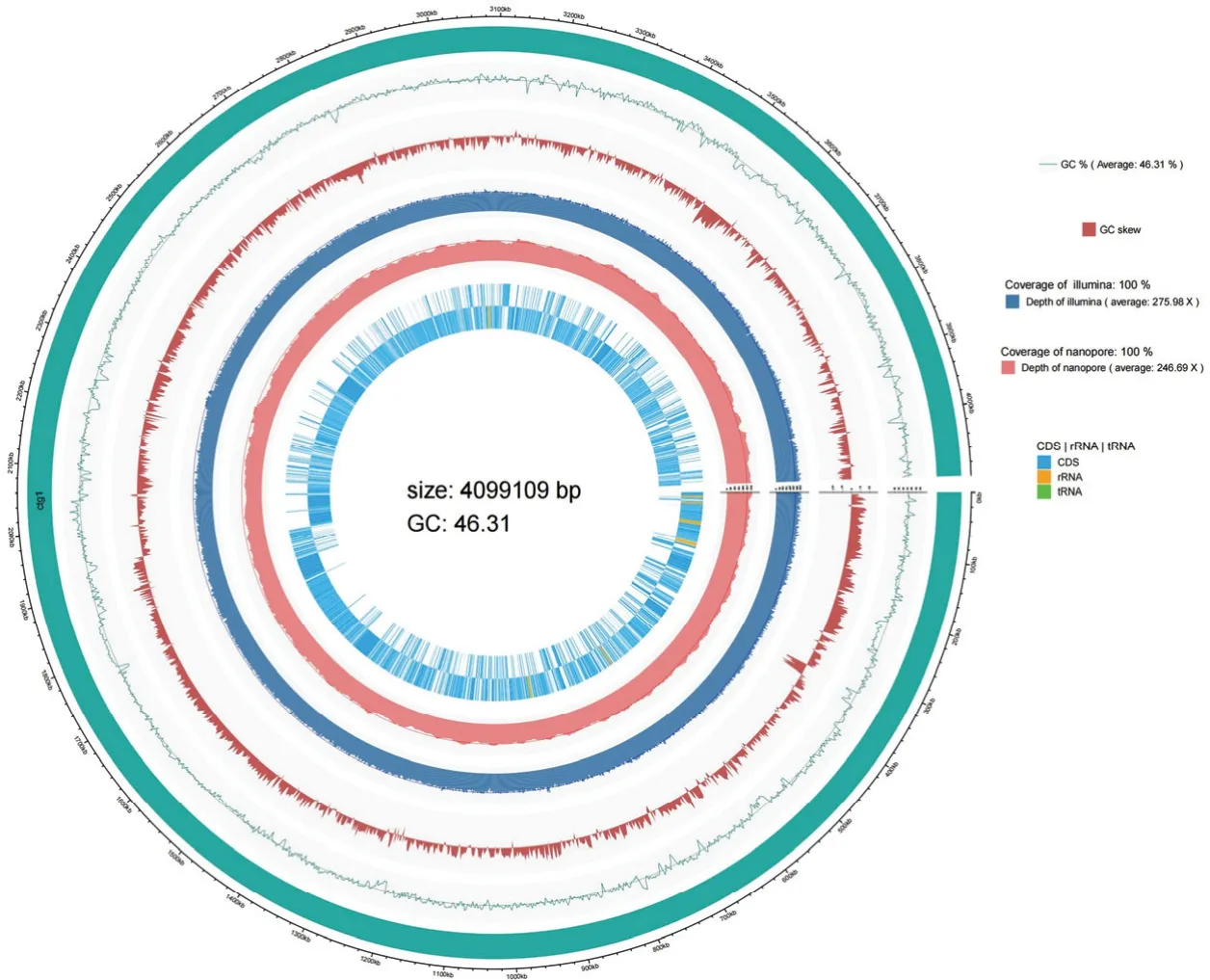
图2 贝莱斯芽胞杆菌Ba-0321 基因组圈图Fig.2 Genome circle map of B.velezensis Ba-0321
2.3 菌株Ba-0321 基因组的GO 功能注释
将菌株Ba-0321 氨基酸序列与GO 数据库进行比对,共得到2992 个注释编码基因,占基因总数的72.92%,分别按照生物学过程(Biological Process)、细胞组分(Cellular Component)和分子功能(Molecular Function)3 个方面进行注释(图3)。分类统计结果显示,在生物学过程方面,其中有56 个基因被注释到抗生素生物合成过程分类中;在分子功能方面,其中有68 个基因被注释到水解酶活性分类中。另外,菌株Ba-0321 含有纤维素酶、几丁质结合蛋白等拮抗活性相关基因和脂多糖生物合成、水杨酸生物合成、乙酰乳酸脱羧酶、乙酰乳酸合酶等诱导抗病性相关基因。以上结果表明,菌株Ba-0321 可通过产抗生素、酶类和诱导植物产生抗病性等途径起作用。
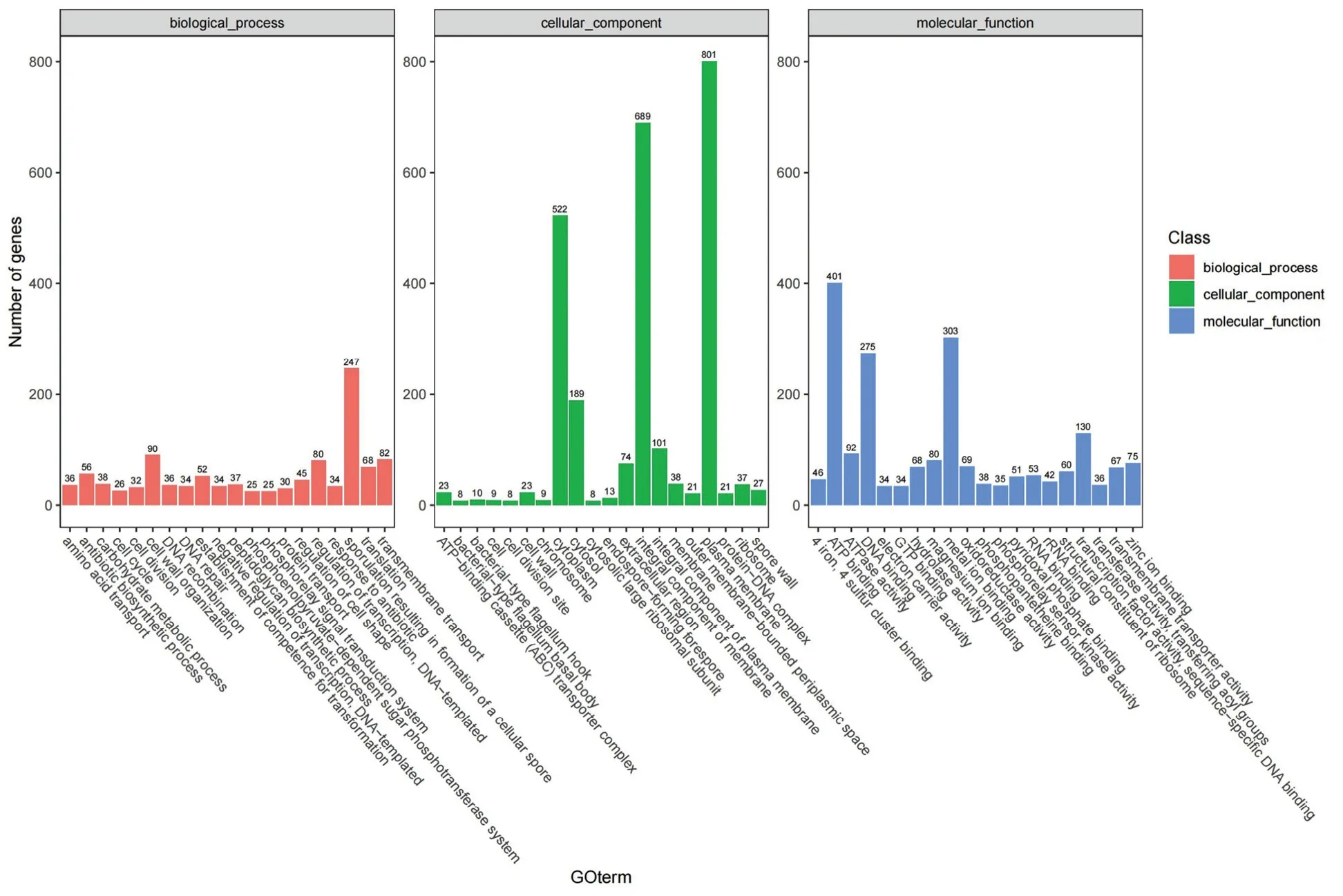
图3 GO 功能分类Fig.3 Gene ontology classification
2.4 菌株Ba-0321 基因组的COG 功能注释
对菌株Ba-0321 基因组中具有生物学功能的蛋白编码基因进行COG 注释,结果发现共有2540 个蛋白编码基因被注释(图4),占基因总数的61.91%,可分为A~Z 共24 类。其中注释到氨基酸转运及代谢(Amino Acid Transport and Metabolism)的基因最多,共306 个,占总注释基因数的12.04%;其次为翻译、核糖体结构和生物合成的注释基因数为204 个,占比为8.03%;碳水化合物转运和代谢、转录及未知功能基因分别有191 个、188 个和185 个。值得注意的是,有62 个参与防御机制(defence machainsm)的基因被注释,其中编码ABC 型多药转运系统基因最多,有21 个;其次,编码多药转运蛋白基因6 个,编码ABC 型抗菌肽转运系统基因和肽酶基因各5 个;编码焦磷酸酶水解酶基因3 个,编码参与碲酸盐抗性的非特征性保守蛋白基因2 个。推测这些基因可能参与了菌株Ba-0321 的抑菌功能。
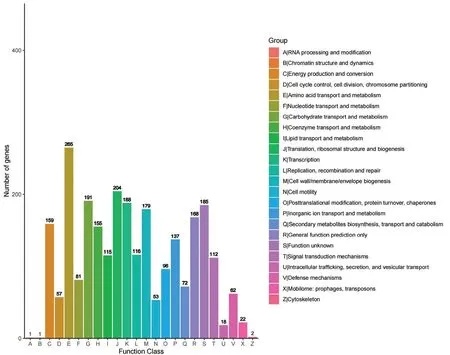
图4 COG 结果分类图Fig.4 Classification diagram of COG results
2.5 菌株Ba-0321 基因组的KEGG 功能注释
将菌株Ba-0321 与KEGG 数据库进行比对分析,对应到KEGG Pathway 的基因有1168 个,占基因总数的28.47%,参与28 条代谢通路,其中基因数超过50 的代谢通路有15 条(图5)。KEGG 富集分析结果表明,碳水化合物代谢(Carbohydrate metabolism)、Global and overview maps 和氨基酸代谢(Amino acid metabolism)是最主要的3 种代谢通路,分别有377、289 和289 个基因被注释。另外,有35 个基因被注释到其他次级代谢产物生物合成途径,有56 个基因被注释到萜类化合物和聚酮化合物代谢途径,推测这些基因与Ba-0321 抑菌活性物质的产生具有密切关系。
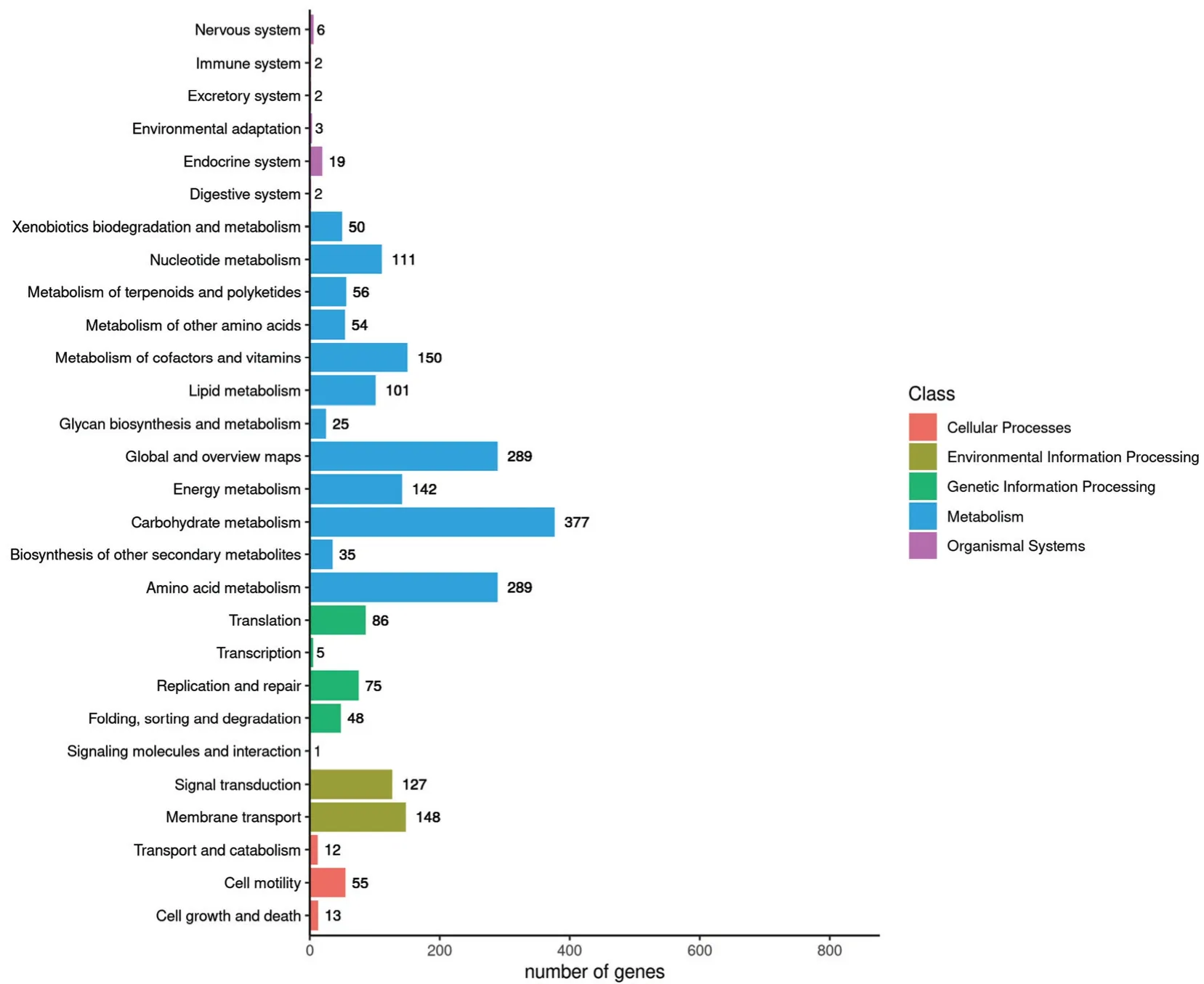
图5 KEGG pathway 结果分类图Fig.5 Classification diagram of KEGG pathway results
2.6 次级代谢产物预测分析
利用anti-SMASH 对菌株Ba-0321 基因组进行次级代谢产物分析,结果发现该菌株共编码13 个次生代谢产物合成基因簇,其中能够找到完全相同或相似度达90%以上的已知基因簇7 个,相似度在20%以下的有2 个基因簇,另外有4 个基因簇未能找到与之相似的已知基因簇(表3)。发现有7 种抗菌活性物质,分别为表面活性素(Surfactin)、大环内酯类抗生素(Macrolactin H)、多烯类(Bacillaene)、丰原素(Fengycin)、聚酮类化合物difficidin、儿茶酚型嗜铁素(Bacillibactin)和溶杆菌素(Bacilysin),除Surfactin 与已知菌株BGC0000433 来源的Surfactin 合成基因簇相似性为91%,其他6 种抗菌素合成基因簇与已知菌株来源的与之对应的基因簇相似性均达100%。Cluster 1 与已知菌株BGC0000926 来源的rhizocticin A 合成基因簇相似性较小,为22%,Cluster 3 与已知菌株BGC0000693 来源的butirosin A/B 合成基因簇相似性最小,仅为7%。比对还发现该菌基因组中存在4 种未知功能的基因簇(Cluster 4、8、9、12),其中萜类(Terpene)2 种,T3PKS(Type III PKS)类1 种、非核糖体肽合成酶(NRPS)1 种。以上结果表明,菌株Ba-0321中存在多种产生抑菌次生代谢产物的基因簇,同时也存在合成新抑菌物质的基因簇,具有较大的生防应用潜力。
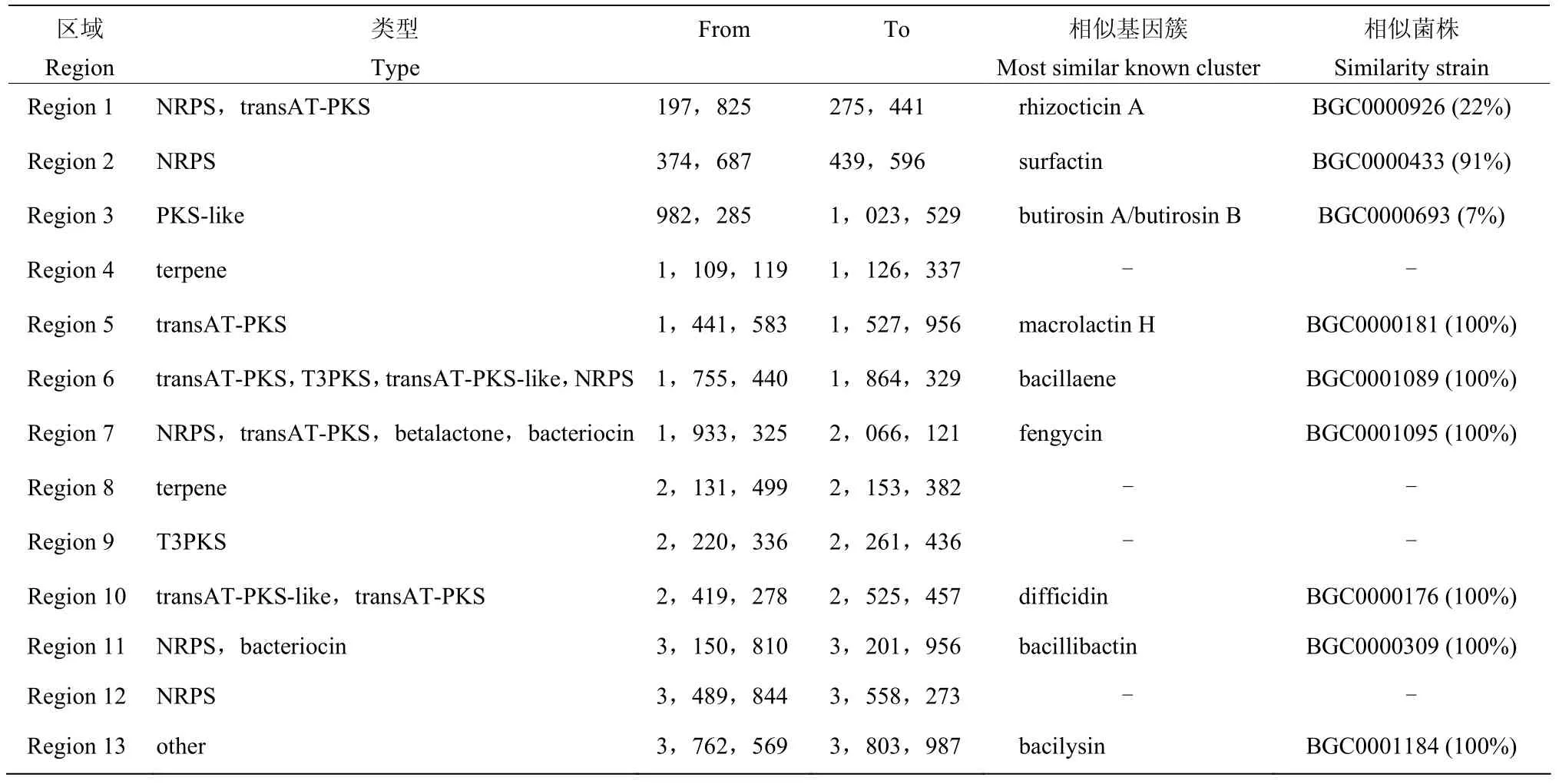
表3 菌株Ba-0321 次级代谢产物合成基因簇Table 3 Gene clusters of secondary metabolite of Ba-0321
2.7 菌株Ba-0321 基因组中抗菌脂肽合成基因和防御机制相关基因的扩增
根据菌株Ba-0321 全基因组序列中编码Surfactin、Macrolactin、Fengycin、Difficidin、Bacillibactin、Bacillaene 和Bacilysin 脂肽合成酶基因(表1)及防御机制相关基因(表2)所设计的特异引物,以Ba-0321基因组DNA 为模板进行PCR 扩增,均能扩增出明亮且单一的条带,且片段大小与预期一致(图6)。序列分析结果表明,扩增获得的PCR 产物序列与设计的目标基因片段序列同源性均为100%,与数据库中其他B.velezensis菌株基因组中相对应的基因序列同源性在91%~100%,进一步证实了菌株Ba-0321 基因组中确实存在抑菌活性物质合成基因簇及防御机制相关基因。

图6 抗菌脂肽和抗性相关基因的PCR 扩增结果Fig.6 PCR amplification results of antimicrobial lipopeptide and resistance related genes
2.8 菌株Ba-0321 产蛋白酶、纤维素酶和几丁质酶的能力
酶活性测定结果表明,菌株Ba-0321 可使酪蛋白培养基、几丁质培养基产生水解圈,显色反应中使纤维素刚果红培养基红色消失,产生透明圈,说明该菌株具有产蛋白酶、几丁质酶和纤维素酶的能力(图7)。
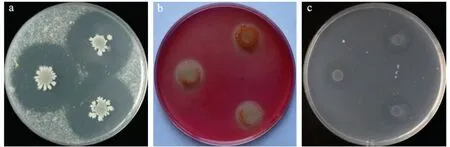
图7 蛋白酶、纤维素酶和几丁质酶活性测定Fig.7 Determination of protease, cellulase and chitinase activities
3 讨论
贝莱斯芽胞杆菌能分泌产生酶、抗菌蛋白、脂肽类抗生素、聚酮类抗生素、植物激素等多种生物活性物质,对抑制病原菌起着重要作用[9,15]。本研究通过共培养的方法明确了贝莱斯芽胞杆菌Ba-0321 无菌滤液对尖孢镰刀菌和疫霉菌菌丝生长的抑制作用,说明该菌株可分泌胞外抑菌代谢产物,同时发现其胞外代谢产物的含量与抑制率的正相关性,但未明确其抑菌物质的种类。进一步通过全基因组测序及基因功能注释,从分子生物学的角度推测出一些与其抑菌功能相关的基因,如编码肽酶、水解酶、多药转运蛋白、纤维素酶、几丁质结合蛋白、抗生素生物合成等基因,以及参与萜类化合物、聚酮化合物代谢途径的基因等,同时推测出菌株Ba-0321 基因组中还含有诱导抗病性相关基因,表明菌株Ba-0321 可能通过多种途径抑制病原菌的生长,这与前人的研究[9,15]基本一致。通过对该菌株基因组中抑菌相关基因的分子验证和抗性相关酶活力测定[16],进一步证实了该基因组测序结果的可靠性,初步明确了该菌株的抑菌机制。下一步将进行抑菌相关基因挖掘、胞外抑菌活性物质的分离提取及微生物农药的研发与利用。
本研究采用antiSMASH 工具预测菌株Ba-0321 的次级代谢产物基因簇,得到脂肽类和聚酮类抗生素合成基因簇13 个,其中有7 个相似性很高的抗生素合成基因簇,如Surfactin、Macrolactin、Bacillaene、Fengycin、difficidin、Bacillibactin 和Bacilysin,这与前人研究基本一致[17-20]。其中,Macrolactin 是一类由聚酮链环化而成的新型化合物,到目前为止,一共鉴定出26 种大环内酯类化合物Macrolactin A-Z 及其衍生物,且大部分是从海洋芽胞杆菌中获得[21,22],多项研究证明大环内酯Macrolactin 对多种植物病原菌具有显著的抑制作用[23-25]。本研究从贝莱斯芽胞杆菌Ba-0321 中预测出Macrolactin H 合成基因簇,并通过分子验证了该抗生素合成基因簇确实存在于Ba-0321 基因组中,进一步丰富并证实了该物质可存在于多种芽胞杆菌中,但该物质是否对烟草尖孢镰刀菌和疫霉菌等病原真菌具有抑制效果还需进一步研究。目前关于Rhizocticins和Butirosin 抗生素相关的研究还较少[26,27]。本研究中发现的Rhizocticin A 抗生素合成基因簇与BGC0000926来源的相似度仅为22%,Butirosin 抗生素合成基因簇与BGC0000693 来源的相似度仅为7%,说明这些基因簇可能合成了新的抑菌物质。菌株Ba-0321 基因组中还存在4 种未知的次生代谢产物合成基因簇,有待于今后进一步挖掘研究。该研究为新型微生物农药的开发提供了很好的资源。
