单细胞分析研究二氧化氯对杀蚊细菌球形赖氨酸芽胞杆菌芽胞的影响
2022-07-09李翠梅王晓春张秀娟刘军贤王桂文
李翠梅,王晓春,张秀娟,刘军贤,王桂文*
(1.广西师范大学物理科学与技术学院,桂林 541004;2.广西科学院,南宁 530007)
蚊虫是我们生活中的常客,不但骚扰我们的工作和睡眠,而且更严重的是传播疾病。常用的化学杀蚊剂容易污染环境,破坏生态,长期使用还会出现抗药性蚊虫。而球形赖氨酸芽胞杆菌Lysinibacillussphaericus(Ls)是一种产芽胞细菌,对幼蚊有特异的毒性作用[1];在适宜的环境下,其芽胞会在死幼蚊体内再循环,是安全有效的生物防治杀蚊剂[2,3]。其主要毒力活性因子是由二元毒素(BinA和BinB蛋白)组成的蛋白晶体,伴随着芽胞的形成而形成,包裹在芽胞外壁内,对库蚊Culexspp.、按蚊Anophelesspp.有很高的毒力,对伊蚊Aedesspp.低毒或无毒[4,5]。虽然芽胞不是杀蚊的主体,但是Ls生长繁殖的重要环节。Ls制剂的施用过程可能会遇到影响制剂活性的不利条件,影响芽胞的萌发和菌体在水体环境的繁殖生长以及死幼蚊体内再循环,进而影响菌剂的生防效果和持效性。
二氧化氯(ClO2)是目前国际上公认的最新一代的杀菌剂[6,7],也是生物制剂实际使用期间可能遇到的农药。ClO2可以有效杀灭大肠杆菌、金黄色葡萄球菌、枯草芽胞杆菌、家蚕微孢子虫、甲型肝炎病毒、小球藻等绝大多数微生物。张晓煜[8]用改进的 Lowry法-二喹啉甲酸(BCA)发现:ClO2破坏了细菌屏障,造成胞内物质渗漏,但不是细菌死亡的主要原因;胞内核酸不是ClO2作用的靶位,菌体死亡与ClO2作用后引起的细胞质凝集密切相关。Ofori等[9]研究了ClO2对大肠杆菌形态、外膜通透性、细胞膜破裂和胞内酶活性的影响,发现ClO2使大肠杆菌的细胞质膜受损、胞内成分泄露。
细菌芽胞作为一种休眠体,胞内有高含量吡啶二羧酸钙(Ca2+-DPA),能耐干燥、受热、辐射、有毒的化学物质等[10]。芽胞一旦萌发,就丧失原有的抵抗力。有几种情形都可以诱发芽胞萌发,一是芽胞内膜上受体蛋白识别营养物质,触发芽胞萌发(如GerA受体识别 L-alanine,GerB/K受体识别AGFK)[11,12];二是非营养萌发剂(如十二烷胺)直接或间接打开SpoVA蛋白通道,引起DPA和芽胞内小分子的释放[13,14];三是芽胞皮层肽聚糖与皮层水解酶(CwlJ和 SleB)结合,触发芽胞萌发;或者是从一个芽胞释放出来的CaDPA可能刺激相邻芽胞的萌发[15,16]。Young和Setlow[17]发现ClO2处理没有导致枯草芽胞杆菌芽胞内核大量释放DPA,被ClO2处理的芽胞可以启动芽胞萌发的最初步骤,但可能由于某种类型的膜损伤而不能进一步发展为营养细胞,具体的机制尚不清楚。此外,前述研究都是基于群体分析得到的认识,而多细胞实时微分干涉差(Differential interference contrast,DIC)显微镜术实现了单细胞水平大量分析单个细菌芽胞的萌发动态,大大促进了对芽胞的萌发机制及理化因子对芽胞的作用机理的认识[14,15,18]。因此,本研究应用单细胞分析方法,一方面探究ClO2对Ls制剂中的芽胞结构和芽胞萌发相关蛋白的影响,另一方面分析ClO2处理造成芽胞不能进一步生长的原因,从单细胞水平补充 ClO2对细菌芽胞杀灭机制的新认识。
1 材料与方法
1.1 试验菌株和培养基
菌株:球形赖氨酸芽胞杆菌采用世界卫生组织推荐的高效杀蚊菌株 2362为研究菌株,由中国科学院武汉病毒所惠赠。
本研究采用的基本培养基如文献[18]所述,但采用的产芽胞培养基是NYSM[3]:蛋白胨10.0 g/L、牛肉浸出粉 3.0 g/L、氯化钠 5.0 g/L,酵母提取物 0.5 g/L,MnCl2·4H2O 0.009895 g/L,CaCl20.0777 g/L,MgCl2·6H2O 0.2033 g/L。经121 ℃、20 min高温灭菌备用。
1.2 芽胞制备
Ls芽胞制备与Bt芽胞制备类似[18],取2%活化后的菌液转入产芽胞培养基(NYSM),30 ℃,250 r/min,培养60~72 h,显微镜镜检芽胞率达到99%以上,离心收集芽胞,并用无菌去离子水清洗和纯化6次。将纯化芽胞放4 ℃保存备用。
1.3 ClO2处理与芽胞萌发
1.3.1 芽胞ClO2处理 将Ls芽胞稀释为浓度约为10–8个芽胞/mL,吸取芽胞悬液0.1 mL,加入2 mL的离心管中,5000 r/min离心机离心5 min,去上清,分别加入1 mL 0.05%、0.075%、0.1%、0.15%、0.3% ClO2,25 ℃处理5 min后加入0.5% Na2S2O31 mL,25 ℃处理5 min以终止ClO2作用;5000 r/min离心8 min,去上清,加入1 mL无菌去离子水洗3次。对照组用无菌去离子水代替ClO2做同样处理(即0% ClO2)。
1.3.2 芽胞存活率测定 芽胞存活率检测如文献[18]所述。芽胞经ClO2处理后做10–3~10–5梯度稀释,吸取0.1 mL在LB固体培养基上,用涂布棒均匀涂布,每个梯度3个平板,在30 ℃生化培养箱培养24~36 h,选择0~200个菌落范围内的平板计数。用自编Cell Counter小程序统计在LB培养基上形成的单菌落数,计算芽胞的存活率。
1.3.3 拉曼数据采集和处理 使用自主搭建光镊拉曼光谱系统(LTRS)[19]采集单个芽胞的拉曼光谱。将对照和ClO2处理过的芽胞悬浮在无菌水中,取0.1~0.2 mL放入样品槽内,在100倍物镜下,光镊随机捕获单个芽胞,收集芽胞拉曼光谱,信号积累时间为20 s。在相同的条件下,收集光镊中没有芽胞的光谱,并作为背景。每个处理共收集50个芽胞光谱和5个背景光谱。数据处理的方法如文献[20]所述:将光谱数据转换为文本格式,导入Matlab程序扣除背景、相邻法平滑曲线,再对光谱基线进行校正,以峰面积法计算特征峰1017 cm-1和1652 cm-1峰的峰面积作为信号强度。
1.3.4 芽胞萌发实时监测 将Ls芽胞用无菌水稀释到10-3梯度,经热激处理(80 ℃,30 min;0 ℃,15 min)后的萌发条件如下。营养萌发:50 mmol/L丙氨酸和25 mmol/L Hepes缓冲液,pH 7.4、37 ℃;非营养萌发:(1)0.8 mmol/L十二烷胺和25 mmol/L Hepes缓冲液,pH 7.4、45 ℃;(2)60 mmol/L外源CaDPA,30 ℃。
芽胞萌发监测过程如文献[18]所述。以每3 s一帧,曝光时间200 ms,连续采集120 min。Matlab自编程序,读取芽胞图像灰度值积分。将数据导入Origin 8.5,以时间为横坐标,芽胞相对亮度为纵坐标,绘制成芽胞萌发动态曲线,读取芽胞萌发参数值。将芽胞萌发经历的三个阶段(三个时间点)分别定义为 Tlag(芽胞启动快速释放 CaDPA),Trelease(CaDPA 完全释放)和 Tlys(肽聚糖皮层完全水解)。ΔTrelease是CaDPA快速释放过程(ΔTrelease=Trelease—Tlag),ΔTlys是芽胞皮层水解的过程(ΔTlys=Tlys—Trelease)。
2 结果与分析
2.1 ClO2处理后芽胞的存活率及拉曼光谱分析
0.05%、0.075%、0.1%、0.15%和0.3% ClO2处理5 min后芽胞的存活率分别约为64%、51%、42%、36%、14%。然而,即使是0.3% ClO2处理5 min,大多数芽胞仍然保留CaDPA(图1A)。
图1B是处理后单个芽胞的平均拉曼光谱,其中661 cm-1、823 cm-1、1017 cm-1、1397 cm-1、1449 cm-1和1572 cm-1是CaDPA的特征峰;1220~1290 cm-1、1630~1680 cm-1是蛋白质二级结构特征峰,其中1260~1290 cm-1、1645~1655 cm-1表征α-螺旋结构,1240~1260 cm-1和 1660~1690 cm-1表征无规则卷曲结构[22]。处理后芽胞的CaDPA和蛋白质的特征峰位置没有明显的改变,但峰强度有所减弱。图1C显示CaDPA(1017 cm-1)峰的强度随处理浓度升高而有所降低,而表征蛋白质α-螺旋结构的1652 cm-1峰和无规则卷曲的1678 cm-1峰强度与对照(即0% ClO2处理)相比没有明显变化。表明 ClO2处理后芽胞有少量CaDPA泄漏情况,但蛋白质的二级结构没有明显的改变。

图1 不同浓度ClO2处理5 min后的Ls芽胞存活率和芽胞DPA保留率(A),单个芽胞平均拉曼光谱(B),芽胞1017和1652 cm-1峰强度(C)Fig.1 Spore viability and DPA retention after 5 min treatment with different concentrations of chlorine dioxide (ClO2) at 25 oC (A),the averaged Raman spectra of treated spores retained DPA and lost DPA (B), the intensities of peaks 1017 and 1652 cm-1 of treated spores and untreated spores (C)
2.2 ClO2处理对单个Ls芽胞萌发的影响
2.2.1 营养萌发 图2A是Ls芽胞在50 mmol/L丙氨酸触发下的群体萌发曲线,对照和0.05%、0.075%、0.1%、0.15%、0.3% ClO2处理的芽胞120 min内的萌发率分别大约是97%、89%、50%、27%、17%和7%。0.05%处理对芽胞萌发影响不显著,而ClO2浓度增加到0.075%之后,萌发率降低了一半。对照芽胞萌发迅速,大多数在10 min内萌发,单个芽胞萌发动态基本相同(图2B)。而0.075% ClO2及更高浓度处理后,芽胞萌发动力学与对照明显不同,萌发速度变慢,CaDPA快速释放过程延长,皮层水解过程也相应延长,在Tlag之前,芽胞DIC图像强度缓慢下降(图2D)。
图2C是芽胞萌发参数。ClO2处理5 min后的芽胞Tlag值显著增加,是对照的2~7倍;ΔTrelease值是CaDPA释放过程,随着ClO2处理浓度增加而增大。ClO2处理5 min后,ΔTrelease值是对照的1~5倍,ΔTlys是对照的1~2倍。ClO2处理后,Tlag时芽胞的相对亮度明显低于对照,显示在Tlag之前已有部分CaDPA泄漏。根据相对亮度推算[23],在CaDPA开始快速释放前,已有约5%~31%的CaDPA已经释放到芽胞外,而对照仅有3%左右。

图2 ClO2处理5 min后Ls芽胞群体的丙氨酸萌发曲线(A),8个单个芽胞的萌发动态(B:对照;D:0.15% ClO2处理)和萌发参数对比(C)Fig.2 Germination of Ls spores treated by different concentrations of chlorine dioxide (ClO2) for 5 min (A), germination dynamics of single spores untreated (B) or treated by 0.15% ClO2 (D), and germination parameters (C)
2.2.2 非营养萌发 十二烷胺是一种阳离子表面活性剂,也可以触发芽胞的非营养剂萌发,但不涉及芽胞的萌发剂受体(GRs),也不激活皮层水解酶,可能是通过打开DPA芽胞内膜上的释放通道,导致CaDPA释放[24]。与营养萌发不同,十二烷胺萌发没有明显的芽胞皮层水解过程。ClO2处理同样影响十二烷胺触发的萌发,120 min内的萌发率由对照的95%依次降为83%、55%、36%、26%、17%左右(图3A)。与丙氨酸萌发相似,处理后的芽胞萌发缓慢,Tlag值增大,ΔTrelease延长,而在Tlag之前,芽胞DIC图像强度缓慢下降(图3 B、D)。萌发参数显示(图3C),与对照相比,处理5 min后的芽胞Tlag、ΔTrelease显著延长,分别增加2~7倍,2~3倍左右。SD值显示,对照组芽胞间Tlag、ΔTrelease差异小,而随着ClO2处理强度的增强,芽胞间Tlag、ΔTrelease值异质性增大。同样的,处理后的芽胞,已有17%~35%左右的CaDPA在Tlag前已经缓慢释放到芽胞外,而对照芽胞只有12%左右。
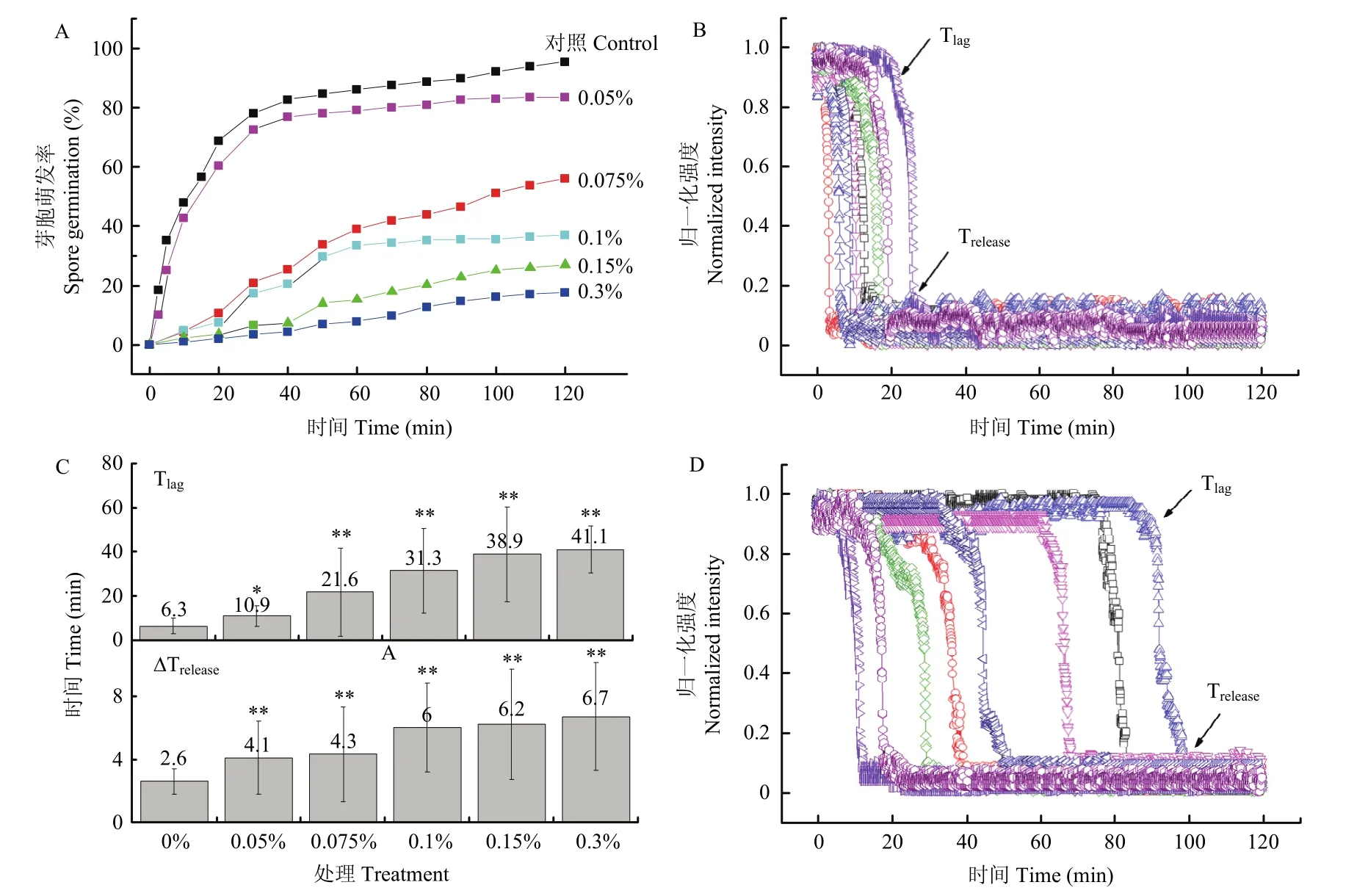
图3 ClO2处理5 min后Ls芽胞群体的十二烷胺萌发曲线(A),8个芽胞的萌发动态(B:对照;D:0.15% ClO2处理5 min),萌发参数对比(C)Fig.3 Dodecylamine germination of Ls spores treated by different concentrations of chlorine dioxide (ClO2) for 5 min (A), germination dynamics of 8 individual untreated spores (B) or spores treated with 0.15% ClO2 (D), germination parameters of single spores (C)
外源CaDPA通过激活枯草芽胞杆菌B.subtilis芽胞外膜上的皮层水解酶 CwlJ、SleB而诱导芽胞萌发[25]。外源CaDPA也可以诱导Ls芽胞的非营养萌发。在120 min内,对照组Ls芽胞萌发率达到97%左右,0.05%ClO2处理5 min后降为87%左右,而0.075% ClO2处理5 min后萌发率仅为6%;更高强度处理后的芽胞则完全失去萌发能力(图4A)。0.05% ClO2处理后的单个芽胞萌发动态与对照类似,只是萌发过程变慢,Tlag值是对照的2倍左右,而ΔTrelease和ΔTlys值基本不变(图4B)。

图4 不同ClO2浓度处理5 min后,外源CaDPA(60 mmol/L,30 ℃)触发的Ls芽胞萌发(A:群体萌发曲线;B:对照和0.05% ClO2处理的萌发参数)Fig.4 CaDPA germination of treated or untreated Ls spores (treated by ClO2 for 5 min at 30 ℃) initialed by 60 mmol/L exogenous CaDPA (A: Germination of Ls spores; B: Germination parameters for untreated and treated spores by 0.05% ClO2 for 5 min)
3 讨论
芽胞存活率显示,Ls对ClO2有一定的抗性,0.3% ClO2处理5 min后86%的芽胞在48~60 h内不能形成单菌落(图1A)。0.15% ClO2处理5 min后,有少量的芽胞内CaDPA泄漏。而拉曼数据显示,CaDPA(1017 cm-1)、蛋白质(1248 cm-1)的峰强度降低,而1652 cm-1的峰强度没有明显变化(图1C)。表明ClO2处理没有明显损伤Ls芽胞内膜。
蛋白质在芽胞萌发过程中起着重要的作用,与芽胞萌发相关的蛋白主要有芽胞内膜上的受体蛋白,皮层水解酶,DPA释放通道蛋白SpoVA以及其他辅助蛋白等[13,14]。在营养萌发中,热激温度、萌发剂浓度及萌发剂的受体蛋白(GRs)数量等3个因素影响了芽胞开始快速释放CaDPA的起始时间[26]。在本研究中,前两个因素相同,而造成ClO2处理后芽胞Tlag值增加(图2D)的可能性是另一个因素,即能感受萌发剂的受体蛋白数量减少了,原因是ClO2处理损伤了芽胞内膜上的部分受体蛋白,使其失去功能。
芽胞在接到萌发信号后,触动芽胞打开内膜上的通道,CaDPA开始快速释放,这个过程与热激处理、GRs 数量、萌发剂浓度没有关系。萌发参数(图2C)显示,与对照相比,ClO2处理的芽胞ΔTrelease值放大了1~5倍,表明ClO2可能也损害了芽胞内膜上的通道蛋白SpoVA[27]。十二烷胺萌发也印证了上述推断。十二烷胺萌发机理是直接或间接打开DPA通道,引起芽胞内小分子和DPA的释放[24],而ClO2处理后芽胞的Tlag、ΔTrelease值分别放大2~7倍和2~3倍,也表明ClO2处理影响了芽胞内膜,CaDPA释放通道及相关蛋白可能受损。
萌发过程需要的蛋白还有皮层水解酶CwlJ、SleB以及相邻的GerQ、YpeB蛋白[28]。水解酶作用于芽胞皮层,使皮层水解,核内吸水膨大,芽胞得以恢复新陈代谢,长成营养细胞。如果皮层水解酶的活力降低以及酶蛋白的数量减少,则皮层水解速度减缓,水解过程延长,甚至不水解。在外源CaDPA萌发中,0.05% ClO2处理后的芽胞萌发Tlag值是对照的2倍左右,而ΔTrelease和ΔTlys值基本不变;0.075% ClO2处理的芽胞萌发率6%,更高浓度或强度的处理,Ls芽胞完全失去萌发力(图4),这表明ClO2处理严重影响了皮层水解酶。而营养萌发中,ClO2处理也令皮层水解过程显著延长(图2)。上述两方面数据结果表明ClO2处理严重损伤Ls芽胞的皮层水解酶,这可能是ClO2处理的芽胞可以启动芽胞萌发的最初步骤,而不能进一步发展为营养细胞的原因[17]。因此,环境因子或杀菌剂对Ls芽胞的不利作用,可能影响Ls菌体在水体环境的生长繁殖能力,进而影响了菌剂的持效性。
Young等[17]在群体水平上研究了 ClO2杀死苏云金芽胞杆菌芽胞的机制,认为可能是由于某种类型的内膜损伤而使萌发的芽胞不能进一步发展为营养细胞,具体原因不清楚。本研究从单细胞水平分析了ClO2对Ls芽胞及其萌发的影响,揭示了ClO2的作用机制。试验结果表明,ClO2处理5 min后大部分芽胞不能形成新的单菌落,但营养和非营养萌发下的单芽胞萌发动态显示,ClO2处理不仅仅损伤萌发剂受体蛋白及其相关蛋白,使其失去功能,而且损伤芽胞内膜的CaDPA释放通道,受损最大的是皮层水解酶;细菌芽胞不能进一步发展为营养细胞的主要原因可能是ClO2严重损伤芽胞皮层水解酶。因此,Ls制剂在实际使用过程要避免与ClO2混用或者接触。
