朱鹮种群增长现状及其影响因素
2023-10-19刘尊海朱龙博雷颖虎张军风
刘尊海,朱龙博,王 晗,雷颖虎,张军风,董 荣,孙 超
(1.西北农林科技大学 动物科技学院,杨凌 712100; 2.秦岭大熊猫研究中心,西安 710000)
朱鹮(Nipponianippon)种群曾极为繁盛,但自20世纪以来,随着工业化进程加速、环境污染加剧、森林采伐过度、湿地破坏严重、人为猎杀频发以及传统农业生产方式的改变,数量急剧下降,甚至在1964年一度被认为灭绝。1981年,我国专家在陕西省洋县发现全球仅存的7只野生朱鹮,同年开展野生朱鹮的抢救性保护工作[1]。直至2000年,朱鹮受威胁等级才从“极危”调整为“濒危”。2021年是朱鹮重新被发现和保护40周年,全球种群数量已从7只发展到7 000只(图1)。

图1 40年全球野生朱鹮数量变化
朱鹮是鹮科鸟类,作为一种古老鸟类且现今为数不多的存活物种,朱鹮的生物学特性蕴含着对环境变化的冲突以及适应。针对朱鹮的研究,不仅可以指导朱鹮现阶段的保护工作,还可以与相关物种进行比对分析,以了解其进化历程以及遗传特征,是不可或缺的遗传资源。朱鹮生态环境要求苛刻,其种群的再次出现,意味着当地生态环境得到改善。随着朱鹮种群的恢复,各生态位趋于稳定,生物多样性得到恢复,生态系统的自我调节能力趋于完善,研究朱鹮在生态系统中的地位以及功能,可以加深对朱鹮及其生态系统的认知,更加科学有效地对其进行保护利用,并且对其他物种的保护利用也具有十分重要的参考价值。但是受到当下生存环境和自身诸多不利因素的影响,朱鹮保护的未来依然充满挑战。因此,本文基于上述背景及意义,结合朱鹮保护现状,对影响朱鹮种群发展的主要原因进行分析,并结合所述因素提出建议,以供朱鹮保护工作者参考。
1 朱鹮种群发展现状
目前,除了通过就地保护、易地保护以及再引入等措施拯救朱鹮以外,我国陕西、河南、浙江、四川以及河北等地分别成立了以保护为主的8个人工饲养繁育基地。近几年多次在中国、日本以及韩国开展朱鹮的重新引入计划,且在河南以及陕西省铜川一带成功形成野外种群[2-3](图2)。2019年王超等[4]在陕西省洋县及其周边地区进行调查,共统计到野生朱鹮数量2 571只,同时发现部分朱鹮出现长距离移动,扩散到周边种群。截至2020年12月,全球朱鹮数量已发展到7 655只,其中野生种群4 453只,人工种群1 988只,野化放归种群1 214只,主要分布在陕西汉中朱鹮国家级自然保护区及其周边县区,是全球唯一的野生种群。现阶段,《朱鹮保护白皮书》数据显示,朱鹮种群中幼体、亚成体及具有繁殖能力的个体占种群数量的75%以上,性别比例约为1∶1,种群年平均增长率为18.85%,种群结构合理,呈现增长趋势。同时,从1981年至今,朱鹮对栖息地的偏好发生了变化,朱鹮的分布中心从偏远的山区转移到了低地、人口较多、湿地面积较大的地区[5],自2016年以来一直停留在人类活动水平中等的地区[6],并且觅食时会接近农事活动区,在足够的预警距离里,对农事活动能产生一定适应性[7]。朱鹮向来被认为是一夫一妻制的物种,然而,在重新引入的种群中,有极少一部分雌性表现出一妻多夫制[8]。同时发现放飞的朱鹮表现出集群营巢的倾向,这与所了解的较强的单独领域性相反[9]。
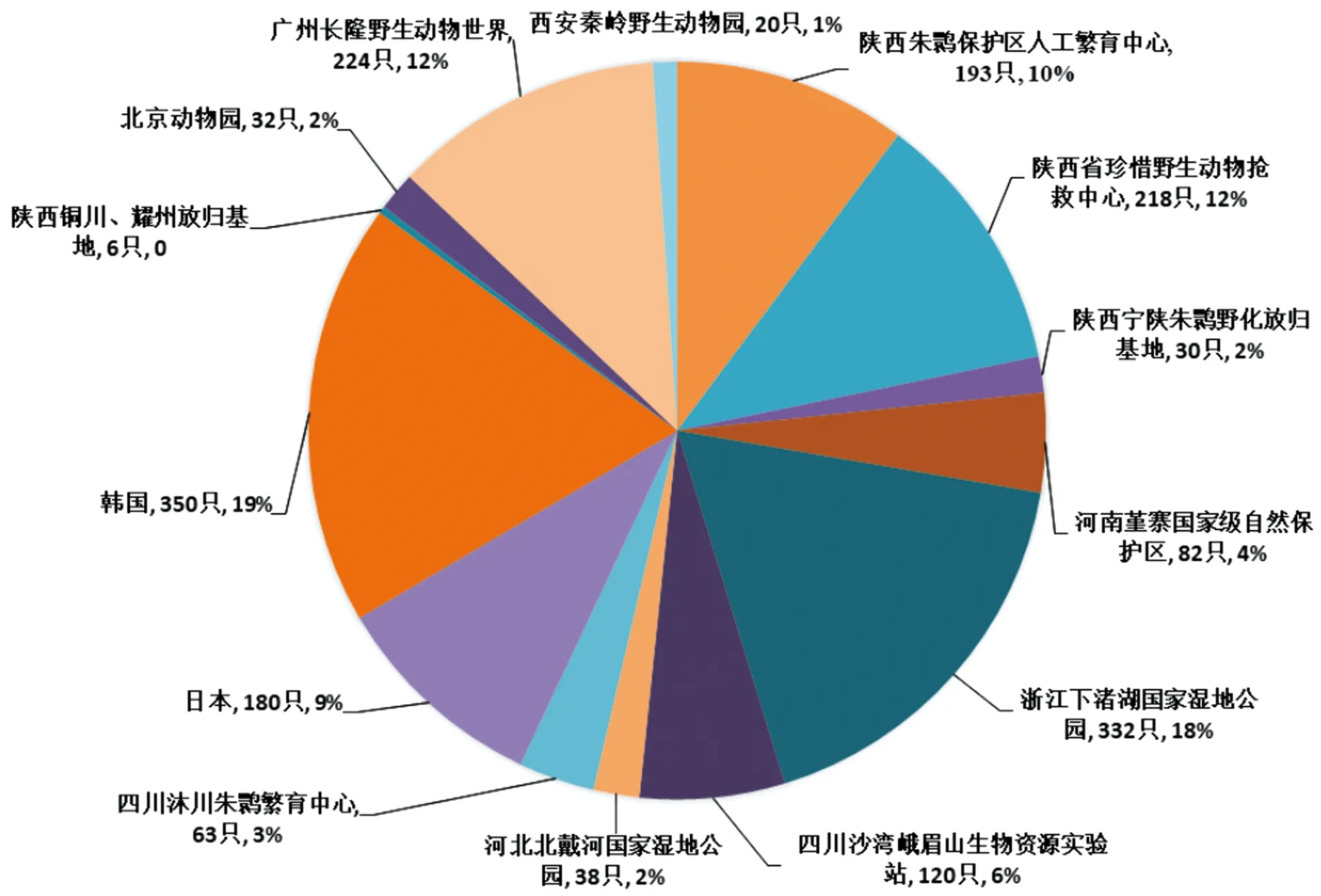
图2 人工朱鹮分布现状
2 朱鹮种群发展影响因素
野生动物的保护和恢复一直是历史难题,动物灭绝事件时有发生。且随着社会发展,生态环境与人类活动的冲突愈演愈烈,在此背景下寻找动物保护的新思路,就必须去发现影响种群繁衍及长期生存的因素。在朱鹮保护40年间,针对朱鹮的研究主要从野外保护过渡到人工繁育,再到如今开始基因组信息研究,这为揭示朱鹮的生长发育以及繁殖特点积累了大量参考材料,并为朱鹮保护和种群恢复提供了科学依据。文章从生物学因素以及环境因素方面,归纳出影响朱鹮种群发展的因素(图3)。
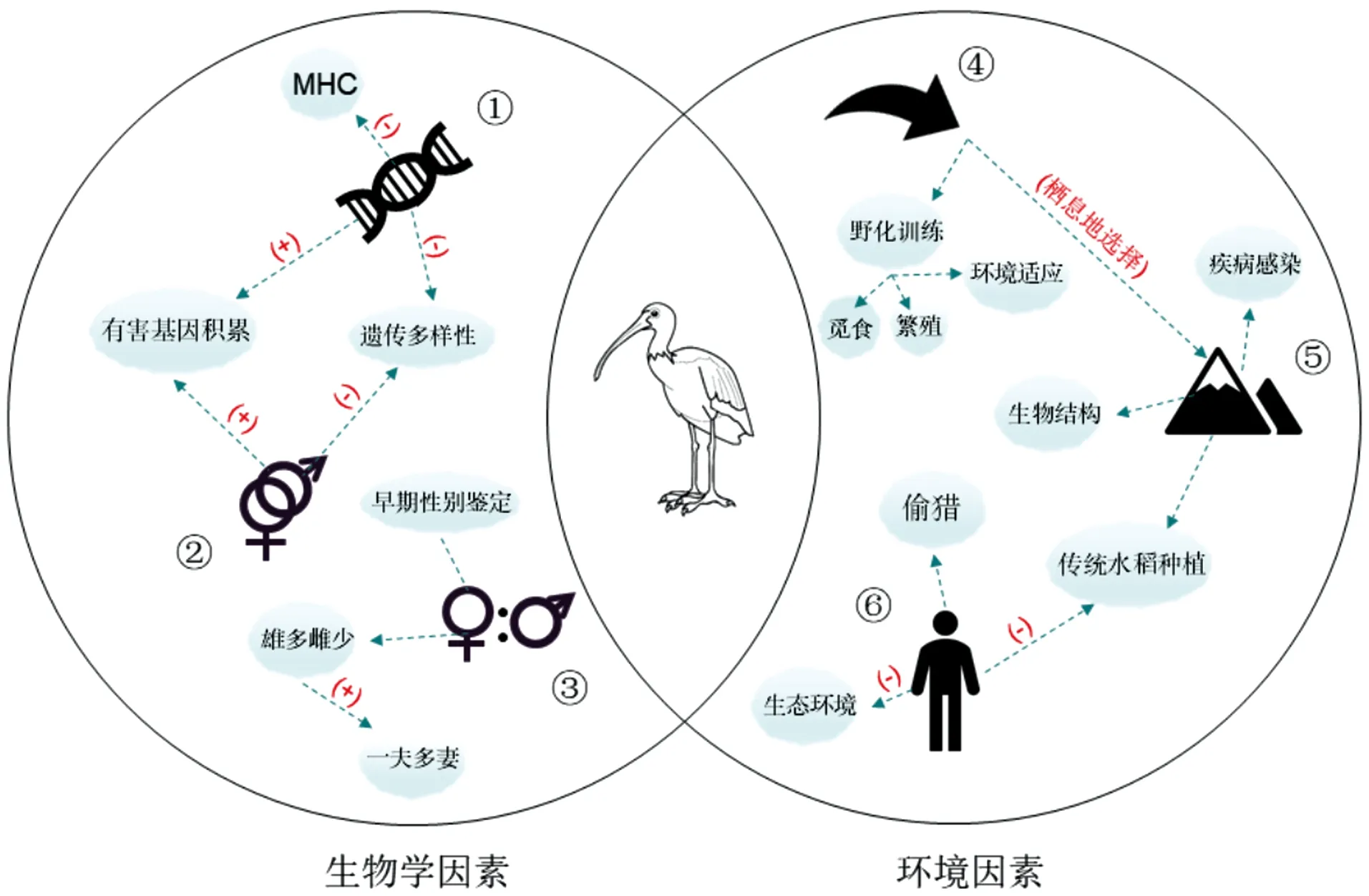
①遗传因素;②近亲繁殖;③性别比例; ④再引入;⑤栖息地;⑥人类活动。
2.1 生物学因素
2.1.1 遗传因素
有研究重建了过去10 000年中朱鹮种群发展史,结果发现,随着种群数量持续下降,当代朱鹮遗传多样性显著减少,并且携带了与近亲繁殖系数升高相关的纯合子有害突变的负载增加[10]。因此,朱鹮保护工作是在遗传多样性极低的背景下,伴随着不断升高的近亲系数下进行,近亲繁殖又反过来作用于遗传多样性以及有害突变的积累,朱鹮遗传多样性低下表现在主要组织相容性复合体(major histocompatibility complex,MHC)上。MHC是脊椎动物免疫系统的重要组成部分,通常被分为MHC Ⅰ和MHC Ⅱ两类,该基因组区域对致病性疾病的抗性和易感性至关重要[11]。现已完全分析了朱鹮MHC的基因组结构[12],并将其应用于朱鹮分子标记辅助选择中。通过对朱鹮MHC II类B基因遗传变异检测,仅获得了5个等位基因,表明该物种MHC II类基因的遗传多样性有限[13]。已知不同MHC等位基因之间共享的高度相似性将增强整个物种对某些疾病的易感性[14]。有学者针对日本最初的5只朱鹮进行MHC多态性的分析,只发现了3种MHC II类单倍型,并且目前日本朱鹮的等位基因/单倍型数量在大多数基因组位点上为3或更少,遗传多样性极低,承受更多的疾病风险[15]。在繁殖中发现雌性更喜欢MHC杂合的雄性,它们具有较暗的婚羽并能产生更多的精子。雄性更喜欢在MHC II类DAB基因座上缺乏DAB*d等位基因的雌性,雌性的平均体重更高,且可以产生更优质的卵。当亲代都具有双方偏爱的MHC基因型时,便会产生更多的受精卵和雏鸟,成活率达到最高[16]。朱鹮遗传多样性不断丧失的现状,已经严重威胁到朱鹮种群的发展,虽然现阶段不可避免,但可以人为干预其发育过程,选择优秀个体集中培养,可以根据雄性婚羽颜色以及雌性体重进行选择,形成具备良好繁殖力的种群,通过得到优秀个体抵消遗传多样性低下带来的不利影响,进而通过再引入等方式,使优秀性状得到扩散,待繁殖数代后,将不同地区的繁殖后代进行杂交,以期利用这种方法来提高朱鹮遗传多样性。
2.1.2 近亲繁殖
近亲繁殖在动物育种中通常被用来获得某些益处,如选育并保持优良种畜。但也会增加有害特征的表达,增加朱鹮病态个体,不利于种群的恢复[17]。庆保平等[18]在保护区调查时发现11只淡色型个体,推测可能是由于多年的近亲繁殖而出现的突变体。2017年对韩国朱鹮的遗传多样性进行评估,发现遗传多样性测度、观察杂合度、预期杂合度和多态信息含量值均低于2016年,认为在有限的群体中发生了近亲繁殖[19]。有学者利用基因组重测序发现死亡个体的遗传多样性较低,近亲繁殖程度更高[20]。随着种群规模的不断扩增,伴随着几乎一半的祖先遗传变异丧失和高度有害突变负荷[10]。在进行濒危动物保护过程中,近亲繁殖导致多样性降低以及有害基因的积累,朱鹮有限的空间加剧了此现象的发生,尤其现阶段虽然再引入种群在各地放飞成功,但是由于景观连通性阻断,后代个体扩散失败集中在亲代栖息地。因此,我们需要通过适当的人类干预,如监测种群结构,了解栖息地环境承载力,趋于饱和或种群内遗传系数过高时应将部分个体放飞至其他栖息地,并适当引入其他种群个体,同时加强适宜栖息地间景观连通性建设,两地间生境建设应满足朱鹮休憩、捕食等行为,以达到促进种群自然扩散和不同种群间交流的作用,既能减轻环境压力,还可以降低近亲繁殖的概率,以此维持种群的亲缘系数,提高种群总体素质。
2.1.3 性别比例
由于朱鹮雌雄同型,给维持种群性别比例造成了困难。早些年由PCR扩增雌性朱鹮W性染色体上的特异性片段,经扩增图谱确定性别[21],后来可以通过测量朱鹮的喙长、喙高综合判定成年朱鹮的性别[22],但不利于雏鸟早期性别鉴定工作开展。通过改进鉴别技术,利用两个引物组,扩增定位在鸟类Z和W染色体上的染色质域-解旋酶-DNA结合蛋白1(CHD1)基因的不同片段,除此之外,利用微卫星位点NnNF05进行性别鉴定同样有效[23]。这项研究为朱鹮早期性别鉴定提供了一种可靠的分子检测方法。通过对宁山县近10年的野外监测数据分析,发现种群规模和灭绝概率与性别比例有关[24],该结论可以在重新引入种群中得到证实,由于性别比例的失调,出现了一妻多夫现象,雌性在产蛋后会立即抛弃第一窝,转而与另一雄性结合配对,易造成一窝筑巢失败[8]。然而,由于野外种群缺乏食物资源,朱鹮在繁殖前期孵育的雏鸟普遍为雄性,因为雄性雏鸟会占用更多的资源,而雌性雏鸟集中在繁殖后期,这种行为会导致雌性雏鸟因缺乏食物而夭折,进而在种群中出现雄多雌少的现象[25],而一妻多夫导致的筑巢失败表明雏鸟阶段的双亲照顾可能是朱鹮成功繁殖所必需的。因此,为避免种群性别比例失衡,尤其在繁殖期内应确保足够的食物资源,形成正常的繁殖行为,纠正后代雄多雌少的现象,不仅如此,应在繁殖行为开始前完成性别鉴定,动态监控种群信息,根据种群结构适当引入或放飞以维持正常的性别比例。
2.2 环境因素
2.2.1 再引入
现阶段,再引入是朱鹮种群恢复的关键。再引入又称为野放,从已有种群中挑选出优良个体,经引入地考察后将其重新放归野外,以重建其历史栖息地并形成一个新的自我繁育的稳定种群。2004年,我国首次再引入在陕西洋县华阳镇进行[26],第二年再次进行并成功实现野外独立繁殖[27],再引入前的野化训练是成功的关键。通常的训练是在大的封闭网笼中进行,在网笼中还原朱鹮的野生环境[28],训练朱鹮独立觅食、繁育以及群体活动的能力。野化训练时所提供的环境条件会影响朱鹮在野外的适应能力,主要在觅食、栖息以及交流互动等方面表现差异[29]。随后,我国相继在陕西省宁陕县[30]、千湖湿地[31]、陕西铜川[3]和河南董寨[32]等地进行再引入。对秦岭一带再引入种群统计评估表明,重新引入的朱鹮种群在一定时间内可能处于调节阶段,秦岭朱鹮的再引入前景广阔,在实现自给自足方面取得了进展[33]。根据宁陕县种群的适应性,预测宁陕县重新引入的种群在未来50年内不太可能灭绝[32]。再引入在全球范围内开展,2008年日本首批人工饲养的朱鹮在佐渡岛被放归野外,此后,在佐渡岛上又进行了6次野化放飞,并在2012年朱鹮数量达到一定规模[34]。2008年以来韩国一直在进行朱鹮的人工养殖,直至2017年韩国也开展了朱鹮的野化放飞计划[35]。再引入无疑是现阶段朱鹮种群恢复的核心,但也存在朱鹮不能有效适应野外环境的现象,一方面应当考虑放飞地质量,生态环境是否适宜朱鹮生存,是否具有充足的资源以供朱鹮栖息、繁殖以及捕食,朱鹮引入后该生态系统的形态结构是否合理并且稳定;另一方面我们应当重视野化训练的质量,严格还原野外环境,摆脱朱鹮对人类的依赖,促进其自然行为的形成和表达,进而选择合适个体放飞,确保再引入个体能够形成稳定的野外种群。
2.2.2 栖息地
栖息地质量以及结构是再引入个体生存和种群建立的重要影响因素[36]。朱鹮经常在稻田、草地、未耕种的稻田和废弃的稻田周围觅食,小型无脊椎动物是主要的猎物成分[37]。朱鹮曾生活在偏远山区,食物充足,而随着杀虫剂的大量使用,导致鱼类数量锐减,直至1981年该现象得到禁止,栖息地环境逐渐改善,朱鹮才得以回到低海拔地区的繁殖地[38]。对宁陕县以及千阳县朱鹮再引入后繁殖地点适宜性评估,发现稻田和降水是选择筑巢地点的主要因素,其中稻田尤为重要[39]。在过去的20年里,冬季淹水稻田不断减少,传统水稻种植的衰落可能会继续对中国朱鹮种群的长期生存和恢复构成威胁[40]。但也发现朱鹮对冬季淹水稻田的依赖在巢址集群之间差异很大,并且这种依赖多年来有所下降[41],在低海拔地区,由于存在其他能够提供丰富食物的湿地,冬季淹水稻田面积不再是限制因素[42]。除此之外,栖息地中的生物种类也会对朱鹮产生影响,在与白鹭的混合种群中降低了警觉时间,并随着规模增加而延长了觅食时间[43]。与此同时,近些年朱鹮受栖息地变化的影响,可能被迫与同类和其他物种更接近,从而导致更高的密度和更多的交流互动,引起疾病传播[44],栖息地改变所造成的压力会影响宿主状况并降低对感染的耐受性,使其更容易感染病原体[45]。鉴于上述现象,首先应将稻田和水源作为栖息地选择的主要因素,以此为根据,在保护现有湿地的同时,恢复朱鹮历史栖息地;其次,可在栖息地中补充食物资源,例如泥鳅等高营养动物;并对可能接触的野生鸟类以及家禽的病原体进行流行病学调查,确定可能的传播途径,提前预防;为减少天敌如蛇、猫以及猛禽对朱鹮的威胁,可以在营巢树下放置敏感物或在树干涂抹润滑物以驱逐蛇和猫,对直接捕获朱鹮的猛禽,在栖息地选择时就应做好调查,合理避开其活动范围或适当进行人为干涉。
2.2.3 人类活动
朱鹮的地理分布区域呈减少趋势,但有预测在未来可能该物种将重新定居其历史分布范围的部分区域[46]。然而人为导致的高死亡风险对种群增长和范围扩大产生了显著的负面影响,近亲繁殖和功能基因的丧失,导致对人类活动的高度脆弱性[47]。农业集约化、耕地扩大和自然植被的清除导致湿地和森林的丧失和退化,从而减少了朱鹮的觅食和筑巢栖息地,使用于种植水稻和其他半水生作物的稻田已成为其不可或缺的栖息地[48]。传统的水稻种植系统是一年一季,长期休耕,为朱鹮提供了理想的食物资源,而在现代农业下,不仅农药对朱鹮构成威胁,同时农业机械和高效的灌溉排水系统促进了复种并缩短了间隔规律的田地休耕期[49],这导致朱鹮将需要承担更大的生存压力。传统农业的改变会导致栖息地严重缩小,对朱鹮构成真正的威胁[50]。虽然人类的行为曾严重危害朱鹮种群,但强有力的法律保护和更高的公众意识减少了这些问题。朱鹮的栖息地总是与人类生活区域交织在一起,朱鹮依靠淹水的稻田觅食并适应人类主导的湿地生态系统,在此背景下栖息地附近的农民被要求保持传统耕种方式,限制化学药剂的使用。如今人类开始利用各种技术手段对种群进行监测以及寻找潜在栖息地;对朱鹮实施就地保护、易地保护以及再引入,并在我国陕西汉中设立了朱鹮国家级自然保护区,对朱鹮种群的恢复有重要作用。
3 朱鹮种群发展现存问题及建议
虽然朱鹮保护现已取得显著成果,但随着朱鹮种群不断扩大,其活动范围以及栖息地逐渐分散,保护工作难以开展,已初步提出了一种基于朱鹮个体叫声的新型识别模型[51],可以极大减少人工监测的误差,能够及时发现问题个体。目前洋县周边存在大量朱鹮,影响朱鹮种群恢复的主要原因已从雏鸟死亡演变成了外伤、撞击和网线缠绕等引起的机械性损伤,并集中于洋县一带[52]。由于空间资源有限,朱鹮已经明显表现出与人类的直接冲突,并且在最近几年尤为突出,也说明目前人类保护意识尚不完善,环境设施建设不符合保护标准,如大量修建工厂、占用耕地以及破坏森林对原栖息地造成了巨大损害,人口搬迁导致耕地荒废,同时传统农业的没落进一步压缩了朱鹮生活环境。朱鹮属于晚成鸟,初生雏鸟需留在巢内接受亲鸟喂养,食物不足会严重阻碍朱鹮种群的增长,同时初生雏鸟各项机能尚未成熟,难以在极端气候以及疫病中存活,天敌侵扰等意外也会导致亲鸟弃巢发生[53]。在上述背景下,应迅速制定再引入计划,释放饱和区域朱鹮种群,缓解栖息地压力,形成更多的野生种群,积极推动朱鹮种群扩增,同时还可以改善朱鹮目前遇到的遗传多样性低下及近亲繁殖严重等问题。在人类角度应合理化解与朱鹮的空间层次冲突,朱鹮具有与人类生活交织的特性,这也造成了目前屡次遭受人类影响的现象,突出问题是在栖息地附近居民保护意识不到位,建筑设施不合理,要谨防高墙、电线、光污染以及工业化进程等对其造成的伤害。
我国已有30多年的朱鹮人工繁育历史,人工繁育技术日渐成熟,但依旧存在诸多问题尚未解决。比如雌雄个体难区分、雄鸟主动弃卵、胚胎死亡、幼雏消化道疾病、眼部感染疾病、幼雏畸形等,而蛋破损率高、死胎数多以及雏鸟发病率高是人工繁育中比较常见的问题,一定程度上阻碍了朱鹮种群发展。并且人工繁育的朱鹮并没有表现出明显优越性,野放后不能有效适应野外环境。我国近些年恶劣天气、自然灾害以及禽类疾病频发。在洋县出现了多次冰雹、洪涝、暴雪等极端天气,并且存在大规模禽流感传染案例,朱鹮生存始终面临巨大威胁。为提供针对性保护建议,发现初孵雏鸟开始繁殖的年龄越早,繁殖寿命更长,终生繁殖成功率更高,并且这种增益投射到下一代,它们具有更大的跗骨和更好的身体状况[54],因此,可以将孵化时间作为优秀个体选择标准,在此基础上,正确引导其尽早开始繁殖,以期得到优良后代。个体选择培养时,应注意性别比例的均衡,避免繁殖失败。然而,目前尚无有效的早期性别鉴定手段,因此,除了改进人工繁育的方法之外,应开展合适的分子标记或者DNA鉴定等研究,并将理论研究应用于实际生产中,以期得到一种新型快捷并且准确的早期性别鉴定方法,以填充在朱鹮繁育方面的空白,解决相关繁育问题。同时,考虑在后续的研究中将遗传标记运用在检测朱鹮多样性或者监控近亲繁殖程度,以应对朱鹮遗传多样性降低带来的适应性下降。
