胶孢炭疽菌侵染秀丽隐杆线虫的特性研究
2023-10-19易杏盈张东华闫晓慧伍建榕
肖 月,易杏盈,张东华,刘 丽,闫晓慧,伍建榕,2
(1.西南林业大学 生物多样性保护学院 云南省高校森林灾害预警控制重点实验室,昆明 650224; 2.西南林业大学 林学院 西南地区生物多样性保育国家林业局重点实验室,昆明 650224)
天然免疫是广泛存在于各类生物体内用于对抗外来入侵物的重要防御体系,其进化地位古老且保守[1]。相关膜受体、信号转导通路及下游效应物在植物、昆虫、脊椎动物和无脊椎动物中被认为是十分保守的。但哺乳动物、线虫和植物的膜受体接受信号刺激后也呈现一定的差异,哺乳动物通常会激活转录因子NF-κB,而植物则激活转录因子WRKY,线虫中未发现以上两种转录因子的同源类似物,这表明动物和植物的天然免疫可能起源于一个共同的祖先,但很早就各自独立进化[2-3]。
油茶炭疽病是油茶种植产业中重要病害之一,引起该病的主要病原菌为胶孢炭疽菌(Colletotrichumgloeosporioides)[4]。胶孢炭疽菌还可侵染核桃、苹果、橡胶等多种经济林树种,危害植物种类较多,严重威胁各国林业生产[5]。Yang等[6]通过转录组分析揭示胶孢炭疽菌感染油茶在酪氨酸代谢、苯丙烷生物合成、黄酮类生物合成和异喹啉生物碱生物合成途径中显著富集。胶孢炭疽菌不是专性寄生菌,有学者已建立希金斯炭疽菌(Colletotrichumhigginsianum)与拟南芥(Arabidopsisthaliana)、胶孢炭疽菌与烟草(Nicotianatabacum)的互作模型[7-8],由于拟南芥等植物培养周期长、实验条件控制难度大等因素制约,致病机制以及互作关系的深入研究难度较大。目前国内外对炭疽病的研究主要集中在化学防治、生物学特性等方面,而关于炭疽病菌与寄主间互作关系及致病机制的分子水平研究较少,限制了该病防治技术的进一步发展[9]。
秀丽隐杆线虫(Caenorhabditiselegans)具有身体透明易观察、生命周期短、已知全基因序列等优点,作为模式生物在病原致病机制等领域得到广泛应用[10]。其能被人类致病菌在内的大量动物病原菌以及植物病原菌感染[11]。因而,利用植物病原菌跨越其自然寄主作用于动物,很可能揭示出天然免疫中最保守和最古老的部分,而此部分可能成为防治该植物病原菌的广谱作用靶点。
研究应用秀丽隐杆线虫评估胶孢炭疽菌对其基本生物学特性的影响,以此探寻胶孢炭疽菌诱导秀丽隐杆线虫天然免疫应答中最为保守和古老的机制。
1 材料与方法
1.1 材料
1.1.1 菌株及孢子菌悬液制备
胶孢炭疽菌从西南林业大学病理实验室获得,经PDA平板培养。孢子菌悬液以PDA液体培养基培养,接入胶孢炭疽菌后25 ℃,180 r/min培养15 d,无菌纱布过滤菌丝,收集分生孢子,M9缓冲液稀释获得相应孢子浓度,血球计数板进行孢子浓度计数[12]。
1.1.2 线虫的培养及同步化
线虫为野生型秀丽隐杆线虫(N2),采用固体线虫生长培养基(NGM),大肠杆菌OP50作为食物源,25 ℃恒温培养[13]。
M9缓冲液收集产卵期成虫置于15 mL尖底离心管中,离心清洗2~3次。收集的成虫中加入1~2 mL裂解液(0.5 mol/L氢氧化钠溶液和体积分数2.5%次氯酸溶液),裂解约5 min,2 000 r/min离心2 min,弃上清液,M9缓冲液反复清洗5~6次,收集管底虫卵,清洗后的虫卵置于无菌培养皿中,加入M9缓冲液,25 ℃恒温孵化16~24 h,获得L1期幼虫,备用[13]。
1.2 方法
1.2.1 菌-虫共培养体系的建立
炭疽菌与线虫培养的培养基差别较大,感染模型需兼顾二者均能较好生长,制定4种培养基:(1) NGM下-PDA上双层板:培养皿底部倒一层NGM培养基,待凝固后再倒一薄层PDA培养基;(2) NGM上-PDA下双层板,方法同上;(3) NGM+PDA混合平板:NGM培养基中分别加入1/50、1/30和1/20 PDA培养基;(4)NGM平板。
取相应浓度胶孢炭疽菌孢子悬浮液200 μL均匀涂布于处理板上,超净台内吹干后加入50 μL经离心浓缩的OP50菌液;以灭活的胶孢炭疽菌孢子悬浮液平板为对照组,将平板放置25 ℃培养箱培养2 d。对照板和处理板接入约50条L1期幼虫,每组设3个平行,25 ℃下恒温培养,每12 h进行线虫存活率测定[14]。
1.2.2 线虫存活率测定
分别将L1期、L4期约50条线虫接种至对照组和处理组各板中,每组设3个平行,25 ℃共培养,每12 h测定线虫存活率。
1.2.3 胶孢炭疽菌孢子浓度与线虫存活率关系的测定
分别设置106、105、104孢子/mL 等3个不同浓度的孢子菌悬液,将约50条L1期线虫分别接于处理组和对照组培养板上,每组设3个平行,每12 h测定线虫存活率。
1.2.4 胶孢炭疽菌对线虫体长大小影响的测定
约100条L1期线虫置于对照组和处理组各培养板上,分别在12、24、36、48和60 h等5个时间点,随机挑取15条线虫,适量10%福尔马林固定后,进行显微镜形态观察和体长大小的测量[13]。
1.2.5 胶孢炭疽菌对线虫繁殖能力影响的测定
线虫繁殖能力通过所产后代数目评价[13]。L1期线虫在NGM板25 ℃下培养至L4末期,挑取一只L4末期线虫放入新鲜处理板和对照板,每组处理设3个平行,48 h后统计线虫后代数目[15]。
1.2.6 胶孢炭疽菌对线虫运动能力影响的测定
将约50条L1期线虫分别接至处理组和对照组平板,25 ℃培养48 h,测定1 min内线虫头部摆动次数和身体弯曲次数,每组处理测定15条线虫[16]。
1.2.7 胶孢炭疽菌对线虫咽泵速率影响的测定
将L1期幼虫在处理板和对照板上培养,分别在48、72和96 h统计线虫每分钟咽泵活动次数,每组处理测定15条线虫[17]。
1.3 统计学分析
用 GraphPad Prism 5进行统计分析,结果用平均值±标准差表示。用t检验法分析组间差异,以P<0.05为差异有统计学意义。
2 结果与分析
2.1 菌-虫共培养体系的建立
2.1.1 NGM-PDA双层培养板
无论PDA处于上层还是下层,胶孢炭疽菌菌丝均生长迅速,PDA位于上层,36 h时菌丝密布全板[图1(a)];PDA位于下层,菌丝在48 h时覆盖全板[图1(b)],菌丝密度均过大,难以观察线虫。
2.1.2 NGM+PDA混合板
在NGM中加入不同浓度PDA(1/50、1/30、1/20)后,胶孢炭疽菌均处于快速生长状态,且PDA含量越高,菌丝生长越茂密[图1(c)~(e)],从而对线虫观察造成干扰。
2.1.3 NGM板
胶孢炭疽菌在NGM板上生长缓慢,产生的菌丝对线虫观察干扰最小[图1(f)]。因而,胶孢炭疽菌感染秀丽隐杆线虫的培养体系采用NGM培养基。
2.2 胶孢炭疽菌对秀丽隐杆线虫存活率的影响
线虫与胶孢炭疽菌孢子共培养,与对照组相比,其存活率显著下降(P<0.05)。48 h后处理组L1期和L4期线虫开始死亡,存活率下降。不同虫龄线虫接触胶孢炭疽菌孢子后半数致死时间呈现不同,L1期为108 h[图2(a)],L4期则为144 h[图2(b)],表明L1期线虫更易受胶孢炭疽菌感染。此外,在处理组部分死亡的线虫体内可显微观察到虫体周围有菌丝破坏虫体表皮的现象[图2(c)和(d)],有20%的线虫有菌丝形成。

(a)和(b)胶孢炭疽菌侵染L1期、L4期线虫的生存曲线; (c)和(d)菌丝从线虫表皮穿出。
2.3 胶孢炭疽菌孢子浓度对线虫存活率的影响
分别设置104~106孢子/mL的孢子菌悬液进行试验,结果表明,对照组中3个浓度的灭活孢子没有显著差异。与对照组比,处理组104孢子/mL的炭疽菌侵染线虫后,线虫存活率无显著差异(P>0.05);而105、106孢子/mL的炭疽菌侵染线虫后,线虫存活率显著下降(P<0.05)。从图3还可以得出,线虫在104~106孢子/mL的孢子菌悬液的半数致死时间分别为156、132和108 h,表明线虫的半数致死时间与孢子浓度有关。并且胶孢炭疽菌孢子浓度与线虫存活率呈负相关性,孢子浓度越大,线虫存活率越低。

图3 胶孢炭疽菌孢子浓度对线虫存活率的影响
2.4 胶孢炭疽菌对线虫体长大小的影响
如图4所示,线虫与胶孢炭疽菌共培养36 h后,处理组线虫的体长明显小于对照组(P<0.05)。表明胶孢炭疽菌可显著影响线虫身体发育过程,从而影响虫体大小。

* 为两组线虫显著相关(P<0.05)。
2.5 胶孢炭疽菌对线虫繁殖能力的影响
后代数目是评价线虫繁殖能力的重要指标,如图5所示,与胶孢炭疽菌共培养后,对照组统计后代幼虫约110个,而处理组则约50个,线虫后代数目显著下降(P<0.01),表明胶孢炭疽菌显著影响线虫的繁殖能力。

** 表示两组线虫极显著相关(P<0.01)。
2.6 胶孢炭疽菌对线虫运动能力的影响
如表1所示,与对照组相比,线虫头部摆动频率和身体弯曲频率均无统计学差异(P>0.05),表明胶孢炭疽菌对线虫的运动能力并未产生影响。

表1 两组线虫运动能力比较次/min)
2.7 胶孢炭疽菌对线虫咽泵速率的影响
线虫的咽泵速率与其摄食状况密切相关[18],是反映其进食能力的重要指标。如表2所示,与对照组相比,经与胶孢炭疽菌共培养后,线虫咽泵速率未发生明显改变(P>0.05),表明胶孢炭疽菌并未对线虫的摄食行为造成显著影响。
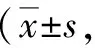
表2 两组线虫咽泵速率比较次/min)
3 讨论与结论
本研究结果表明,胶孢炭疽菌孢子对秀丽隐杆线虫存活率、繁殖能力、体长大小均产生显著影响,而对线虫运动能力和咽泵速率无明显改变。胶孢炭疽菌可导致线虫存活率显著下降,且该菌对线虫的伤害与线虫的虫龄密切相关,于L1期接触孢子的线虫,死亡速度明显快于L4期才接触孢子的线虫,原因与线虫天然免疫系统发育的完善程度相关,虫龄越小其免疫系统发育越不完善,对逆境的抵抗力较弱,而进入L4期,线虫天然免疫系统基本发育完善,因而对胶孢炭疽菌产生一定的抗性,这与Guo等[19]和卢伟伟等[20]的研究结果一致。胶孢炭疽菌孢子浓度与秀丽隐杆线虫存活率呈负相关性,孢子密度越高对线虫产生的伤害越大,这与胶孢炭疽菌菌丝生长密度及代谢产物浓度相关。此外,经显微观察发现,部分死亡虫体周围有菌丝破坏虫体表皮的现象,这与马尔尼菲青霉菌(Penicilliummarneffei)、白色念珠菌(Candidaalbicans)和一种新发现的卵菌Myzocytiopsishumicola感染线虫后的现象相似[21-23]。除存活率外,在菌-虫共培养体系中,线虫的体长大小显著缩短,表明胶孢炭疽菌对线虫生长发育产生显著影响;该菌同时也明显降低线虫的繁殖能力。由此可见,作为典型的植物病原菌,胶孢炭疽菌也可在多个方面对线虫造成伤害,呈现出一定的致病作用,而该伤害究竟是由菌丝产生的机械性损伤还是菌体生长过程中产生的代谢物的毒性导致,亦或者为二者共同作用的结果,特别是植物病原菌产生的毒素对低等无脊椎动物线虫产生危害的生物防治方法和线虫的天然免疫及相互关系有待进一步研究。
在植物中,胶孢炭疽菌感染油茶可引起落叶、落果、枝梢枯死,甚至造成整株、整片衰亡[24]。这与胶孢炭疽菌引起线虫死亡率上升及繁殖能力下降的现象具有一定的相似性。可见,胶孢炭疽菌可跨越自然植物寄主对秀丽隐杆线虫的基本生物学特性产生显著影响,且不同虫龄响应胶孢炭疽菌伤害的差异表明线虫的天然免疫在其中发挥着重要作用。因而该菌-虫共培养体系可能成为探索动物和植物天然免疫系统进化关系以及寻找天然免疫中最古老和保守部分的理想工具,由此可进一步探索防治此类病害的广谱性作用靶点。
