食品安全菌生产蔗糖异构酶的研究进展
2023-10-19陈慧玲沈浩天李宪臻郭小宇
陈慧玲,沈浩天,姜 晨,李宪臻,郭小宇
(大连工业大学 生物工程学院 微生物资源与催化实验室,大连 116033)
蔗糖作为一种易于吸收的营养素,能够为有机体快速提供能量,但过量摄入蔗糖容易引起肥胖及糖尿病等健康问题。因此,蔗糖的健康替代品——异麦芽酮糖引起了研究者的关注。与蔗糖相比,异麦芽酮糖具有耐酸性高、吸湿性低、非致龋齿性和安全性高,在血液中分解缓慢,不刺激胰岛素分泌,避免脂肪过度积累等优点[1]。异麦芽酮糖在自然界中含量低,且难以使用化学方法合成,主要利用微生物转化法及酶转化法生产。但微生物转化法存在分离提取较困难、生产强度弱等缺点,工业上一般采用酶转化法生产异麦芽酮糖[2]。酶转化法的关键是蔗糖异构酶(SIase),该酶亦称异麦芽糖合酶、蔗糖α-葡萄糖基转移酶和海藻糖合酶。蔗糖异构酶主要通过微生物发酵获得,如非食品安全级菌株大肠杆菌(Escherichiacoli)、食品安全级菌株枯草芽孢杆菌(Bacillussubtilis)、解脂耶氏酵母(Yarrowialipolytica)、乳酸乳球菌(Lactococcuslactis)等。与非食品安全级菌株相比,食品安全级菌株不存在热原和内毒素等安全问题,其表达的酶制剂、抗体等可直接用于食品药物,所以利用食品安全级菌株进行蔗糖异构酶的异源表达备受关注[3]。本文总结了近十年的文献报道和最新进展,系统阐述了蔗糖异构酶的来源、结构、催化机制和在食品安全菌株中的异源表达进展。
1 蔗糖异构酶概述
1.1 蔗糖异构酶的来源微生物及性质
蔗糖异构酶来源菌株的筛选及分离工作始于20世纪50年代,Ralf等[4]从甜菜厂排放的废水中第一次筛选并分离出可以产生蔗糖异构酶的红色精朊杆菌(Protaminobacterrubrum)。经过60多年的研究,已经分离获得多种能够合成蔗糖异构酶的微生物[4],如分散泛菌(Pantoeadispersa)、红色精朊杆菌(Protaminobacterrubrum)、肠杆菌(Enterobactersp.)、大黄欧文氏菌(Erwiniarhapontici)、嗜中酸假单胞菌(Pseudomonasmesoacidophila)、普城沙雷氏杆菌(Serratiaplymuthica)、克雷伯氏杆菌(Klebsiellapneumoniae)和放射性土壤杆菌(Agrocbacteriumradiobacter)。将这些生产蔗糖异构酶的微生物按照其生产的蔗糖异构酶的催化主产物进行分类,主要分为两类微生物[5]:(1)以催化主产物为异麦芽酮糖的异麦芽酮糖产生菌,主要以P.dispersaUQ68J、Klebsiellasp.LX3为代表,该类生产菌的异麦芽酮糖的转化效率达到60%~90%;(2)以催化主产物为海藻酮糖的海藻酮糖产生菌,主要以P.mesoacidophilaMX-45、A.radiobacterNX-232为代表,该类生产菌的海藻酮糖的转化效率达到80%~95%。
不同微生物来源的蔗糖异构酶都是单亚基分子,蛋白分子质量大部分集中在62~70 ku,其等电点大多小于7.0,属于酸性蛋白质,S.plymuthicaATCC15928的蔗糖异构酶等电点偏高,为9.0。蔗糖异构酶反应的最适温度在30~40 ℃,最适pH在5.0~6.0,如表1所示。蔗糖异构酶与蔗糖反应的温度和pH对产物组成有一定的影响,P.dispersaUQ68J蔗糖异构酶与底物的反应温度从20 ℃升到40 ℃时,异麦芽酮糖与其他产物比例从21∶1降低至8∶1;当pH 3.0时,K.planticolaUQ14S蔗糖异构酶无异构活性,但随着pH值的增加,异麦芽酮糖成为主要产物。此外,不同来源的蔗糖异构酶的底物亲和力存在较大差异,E.rhaponticiNX-5的Km达到257 mmol/L,而P.dispersaUQ68J的Km仅为40 mmol/L。

表1 不同来源的蔗糖异构酶的相关性质[2]
1.2 蔗糖异构酶的结构
为探索不同蔗糖异构酶转化蔗糖的主产物不同的原因,对多种蔗糖异构酶的晶体结构进行X射线衍射解析。经蔗糖异构酶三级结构的分析,发现各种转化蔗糖形成不同主产物的蔗糖异构酶三级结构具有高度相似性,都是由N-结构域、C-结构域和亚结构域组成的单亚基分子,如图1所示[2,6-9]。其中,N-结构域是一个(β/α)超二级结构,是蔗糖异构酶最重要的部分。在N-结构域中,Asp241、Glu295、Asp369、His145和His368(编号参照于1M53)这5个高度保守的氨基酸构成蔗糖异构酶的催化口袋,参与底物的催化[6-9],Glu295可以作为酸催化剂使蔗糖α1-β1糖苷键的氧质子化导致底物水解,Asp241亲核攻击异头碳的氢原子形成葡萄糖基-酶中间体,Asp369与葡萄糖基的O2和O3形成氢键,His145和His368分别与O6、O2形成氢键[7]。在活性位点附近存在一个保守的序列,该序列与酶的特异性有关,在蔗糖异构化过程中影响产物的比例。主产物为异麦芽酮糖的蔗糖异构酶其保守的序列为RLDRD序列,而主产物为海藻酮糖的蔗糖异构酶的序列为RYDRA,与RLDRD相比存在两个碱基的差异[8]。
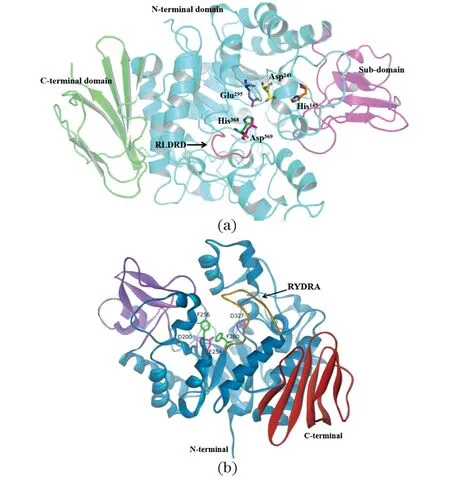
(a)主产物为异麦芽酮糖的蔗糖异构酶三维结构;(b)主产物为海藻酮糖的蔗糖异构酶三维结构。
1.3 蔗糖异构酶的反应机理
进一步探索蔗糖异构酶转化蔗糖的主产物不同的原因,研究了蔗糖异构酶生成异麦芽酮糖或海藻酮糖的机理。Véronèse等[10]从沙雷菌ATCC 15928中纯化了蔗糖异构酶并提出了分子内重排的设想。Ravaud等[7]以来源于P.mesoacidophilaMX-45的蔗糖异构酶作为研究对象提出两步双置换机制。Véronèse等[11]基于动力学分析提出蔗糖异构酶作用原理为乒乓机制。对蔗糖异构酶活性位点反应机制预测为(1)蔗糖异构酶与蔗糖结合形成复合物;(2)酶活性位点的Glu的羧基离子作为酸催化剂,使蔗糖α1-βα1糖苷键的氧质子化;活性位点的Asp攻击蔗糖的C1氢原子,使C1氢原子脱质子,形成葡萄糖基-酶中间体[7];(3)果糖基异构形成异麦芽酮糖或海藻酮糖[7,11];(4)蔗糖水解生成果糖和葡萄糖。催化过程如图2所示。蔗糖异构酶催化机理的研究为后续蔗糖异构酶分子改造提供理论基础。
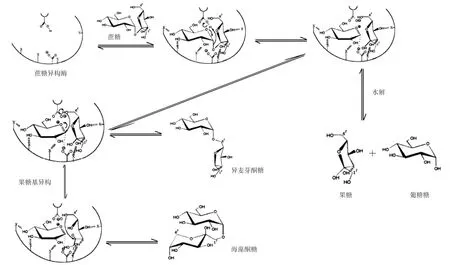
图2 蔗糖异构酶催化蔗糖的过程[11]
2 利用食品安全菌异源表达蔗糖异构酶的进展
目前已有大量关于利用野生菌发酵生产蔗糖异构酶的报道,但存在周期长、发酵液成分复杂、分离纯化复杂等缺点,无法实现工业化生产[1]。因此,利用工程菌异源表达蔗糖异构酶转化蔗糖生产异麦芽酮糖是工业生产异麦芽酮糖的主流。
2.1 食品安全菌株概述
重组微生物在工业领域主要被用作表达宿主来生产蛋白质。用于食品和医疗领域的蛋白质因其特殊性,在安全方面具有更高的要求,如不能包含或产生毒素、热原和过敏源物质等[12],因而需要采用食品级微生物进行表达使其符合食品安全或医药安全标准。被一般公认安全(Generally Recognized As Safe,GRAS)的微生物主要有真菌中的酵母菌、细菌中的枯草芽孢杆菌和乳酸菌等。
2.1.1 酵母表达系统
酵母是基因工程常用的底盘细胞,由于遗传操作简单、培养成本低等优点,被广泛用于各种小分子和大分子化合物的生产[13]。目前常用的酵母主要为毕赤酵母(Pichiapastoris)、解脂耶氏酵母(Yarrowialipolytica)、酿酒酵母(Saccharomycescerevisiae)等。
毕赤酵母广泛应用于食品药品生产。毕赤酵母具有强启动子AOX(醇氧化酶基因),可用甲醇严格调控异源表达的水平;发酵培养中不含蛋白,分离纯化简单;外源基因能以高拷贝数整合到毕赤酵母基因组中,结构稳定不易丢失[14]等优点。目前,已有上千种蛋白在毕赤酵母系统中得到成功表达,如黄原胶内切酶[15]。
解脂耶氏酵母是非常规半子囊酵母,被认为是非致病性的,因其对pH、高盐浓度和有机化合物有高耐受性;可以利用廉价碳源生长;有高效表达外源蛋白的能力,表达的外源蛋白不会被自身的蛋白酶降解;蛋白表达糖基化程度低等优点[16],多种蛋白已成功在该系统中异源表达,如葡萄糖氧化酶[17]。
酿酒酵母由于遗传背景清楚、遗传操作简单;可以进行蛋白翻译后修饰;发酵工艺简单,成本低廉;能将外源蛋白分泌到培养基中;不携带病毒,在蛋白生产方面极具优势,并被认为是用于生产食品和保健产品的安全菌株[13]。因此,酿酒酵母是目前工业领域应用较多的表达宿主,如细胞色素还原酶[18]等在酿酒酵母中实现高水平异源表达。
2.1.2 芽孢杆菌表达系统
枯草芽孢杆菌(B.subtilis)发酵历史悠久,不产生毒素和致热致敏原,故被归为食品级微生物范畴[12]。枯草芽孢杆菌无密码子偏好性,其蛋白分泌能力强且不易形成包涵体;本身拥有高效的信号肽和分子伴侣,可以将蛋白质分泌到培养基中,方便目的蛋白的分离和纯化,可降低成本[19]。由于这些优良特性,枯草芽孢杆菌表达系统已被构建用于各种蛋白的生产中,如淀粉酶[20]等。
桥石芽孢杆菌(Brevibacilluschoshinensis)作为食品与医药行业领域的商业食品安全菌株,发酵过程中无需纯氧的补充,发酵成本低;蛋白表达量高、单位体积细胞产率高[21]。目前,桥石芽孢杆菌表达系统已经成功用于多种外源蛋白的表达,如磷脂酶D[22]等。
2.1.3 乳酸乳球菌表达系统
乳酸乳球菌(Lactococcuslactis)作为全球公认安全的微生物,具有分泌蛋白能力强、表达产物纯化简单、非致病性等特点,是异源蛋白表达和分泌的理想宿主[23]。随着基因工程的发展,乳酸乳球菌表达系统被进一步探索,通过多个诱导型和组成型启动子成功构建乳酸乳球菌表达系统高效表达外源蛋白,如多铜氧化酶[24]。
2.2 利用食品安全菌异源表达蔗糖异构酶的研究进展
由于食品级表达系统存在巨大应用潜力,其已成为分子生物学研究热点[12]。故蔗糖异构酶在食品安全级宿主中的异源表达,如毕赤酵母(Pichiasp.)、解脂耶氏酵母(Y.lipolytica)、枯草芽孢杆菌(B.subtilis)、桥石芽孢杆菌(B.choshinensis)和乳酸乳球菌(L.lactis),取得突破性进展。
2.2.1 酵母表达系统生产蔗糖异构酶
Cao等[25]将来自Klebsiellasp.LX3的蔗糖异构酶编码基因PalI构建在毕赤酵母中,重组酶的酶活力为36.6 U/mL。罗科[26]将来源于红色精朊杆菌的蔗糖异构酶进行突变获得一个突变体PRSiQRY,将其构建到pPICZA载体,表达载体pPicZa-PRSi-QRY转入毕赤酵母X33获得重组菌株,蔗糖异构酶的胞外酶活力达到1 219 U/mL。宋蕾等[27]对Klebsiellasp.LX3的蔗糖异构酶编码基因PalI进行密码子优化,将优化后的PalI成功在解脂耶氏酵母中异源表达,酶活力达到916 U/mg。Zhang等[28]将P.dispersaUQ68J中编码蔗糖异构酶的基因构建在解脂耶氏酵母中重组表达,酶活力最高为7.43 U/mL,异麦芽酮糖转化效率高达97.8%。
2.2.2 芽孢杆菌表达系统生产蔗糖异构酶
Wu等[29]使用穿梭质粒pHA01,将来自E.rhaponticiNX-5的蔗糖异构酶首次在枯草芽孢杆菌WB800中过表达,全细胞酶活力达到5.2 U/mL。王兵兵[30]将来源于Klebsiellasp.LX3的蔗糖异构酶编码基因PalI构建到pHT43质粒上并于B.subtilisWB800中表达,并通过定点诱变将459位Ser和452位Arg置换为Ala,导致50 ℃下蔗糖异构酶的半衰期增加11倍,显著提高了热稳定性,并略微改善了异麦芽酮糖的产物比。刘军彤[21]将来源于P.dispersaUQ68J的蔗糖异构酶基因成功在桥石芽孢杆菌中异源表达,胞外蔗糖异构酶酶活力达到138 U/mL。在此基础上邹亮[31]通过启动子优化获得最优启动子Papr,其介导的重组菌BCpNapr-SI的胞外上清液酶活力最高,为85.1 U/mL。
2.2.3 乳酸乳球菌表达系统生产蔗糖异构酶
Park等[32]首次报道了来自Enterobactersp.FMB-1的蔗糖异构酶在乳酸乳球菌MG1363中的异源表达,通过使用自诱导启动子(P170)和优化的信号肽(SP310mut2),重组蔗糖异构酶成功地在乳酸乳球菌胞外分泌表达,异麦芽酮糖转化率达到72%。乳酸乳球菌表达系统已成为生产蔗糖异构酶的新兴系统,但其遗传背景尚不清楚,需要进一步探索。
2.3 利用食品安全菌株异源表达蔗糖异构酶存在的问题
目前,各种来源的蔗糖异构酶已成功在多种食品级安全菌株中实现异源表达,但工业大规模生产上仍存在一些影响因素:(1)游离的蔗糖异构酶稳定性差、易被降解和污染、重复使用率低、发酵培养基中成分复杂、背景蛋白多、分离纯化繁琐复杂,导致工业生产成本增加,并且胞内的蔗糖异构酶需要离心破碎,步骤多,损耗大,因而后续可以研究蔗糖异构酶的固定化;(2)现有的食品级重组菌株蔗糖异构酶的表达水平依旧达不到工业大规模生产的要求,导致大批量工业化生产价格过高,因而构建高效的食品级基因工程菌仍是蔗糖异构酶大规模生产的关键;(3)蔗糖异构酶与蔗糖反应生成异麦芽酮糖或海藻酮糖的同时还伴随葡萄糖和果糖等副产物的生成,为了进一步提高蔗糖异构酶的转化效率,需要分析关键区域对蔗糖异构酶催化性能的影响,提高底物转化的特异性,通过分子生物学方法改造蔗糖异构酶编码基因中的相关区域,提高其转化效率。
3 提高食品安全菌中蔗糖异构酶表达的优化策略
由于利用食品级安全菌株工业大规模生产蔗糖异构酶存在限制因素,因而提出多种优化策略提高食品安全菌中蔗糖异构酶的表达水平,如将游离酶固定化、优化启动子和信号肽、优化培养基和发酵条件。但由于食品级安全菌转化效率低、筛选方法繁琐、无法进行简易的高通量筛选等因素,直接在食品级安全菌株中进行的蔗糖异构酶蛋白质工程还未涉及。
3.1 蔗糖异构酶在食品安全菌的固定化
近年来,蛋白质展示技术取得巨大进展。Lee等[33]使用锚定蛋白的糖基磷脂酰肌醇(GPI)锚定附着信号序列,将来自肠杆菌FMB-1的蔗糖异构酶成功展示在酿酒酵母EBY100的细胞表面,与游离酶相比,酿酒酵母EBY100细胞表面的蔗糖异构酶热稳定性显著增强;Li等[34]将P.dispersa的蔗糖异构酶编码基因,以细胞壁蛋白Pir1为锚蛋白,成功地在解脂耶氏酵母细胞表面表达,其异麦芽糖转化率为93%,且重复使用12次后转化效率仍能达到80%;吴琦等[35]将锚定蛋白Pir1与Klebsiellasp.LX3的蔗糖异构酶编码基因融合,在解脂耶氏酵母细胞表面展示,酶活力达到4 694.6 U/g细胞干重。固定在细胞表面的蔗糖异构酶有较好的pH稳定性和热稳定性,其二糖产物与单糖副产物的比例可达到91∶9;Zhang等[36]使用枯草芽孢杆菌的孢子外膜蛋白CotX作为锚定蛋白将来自E.rhaponticiNX-5的蔗糖异构酶显示在B.subtilis168孢子上,被锚定的蔗糖异构酶具有较高的生物活性和稳定性,对从农业残留物中、未经处理的甜菜糖蜜和大豆粉经济生产异麦芽酮糖,转化率达到92%;Zheng等[37]利用糖基磷脂酰肌醇(GPI)-细胞壁蛋白(CWP)锚定信号序列构建P.dispersa蔗糖异构酶表面展示载体,并成功地在解脂耶氏酵母细胞表面展示,其酶活力达到2 910.3 U/g细胞干重。与游离的蔗糖异构酶相比,该酶有较好的热稳定性,且以低成本的甘蔗糖蜜为底物时,经过9批加工后,异麦芽糖的转化率仍保持在85%。
3.2 启动子和信号肽优化
信号肽或启动子优化是提高酶表达或分泌的常用方法。Guo等[38]采用半合理方法筛选出13个候选信号肽,比较其对KsLX3-SIase分泌的影响,结果表明,信号肽WapA最有效地引导KsLX3-SIase分泌到培养基中,摇瓶培养显示其酶活力为23.0 U/mL;邹亮[31]对启动子进行优化,考察了组成型启动子Papr、Pnpr、Pamy和诱导型启动子Pxyl对蔗糖异构酶在B.choshinensis的转录水平,发现启动子Papr调控的蔗糖异构酶的胞外酶活力最高,达到85.1 U/mL;Zhang等[39]又利用强组成型TEF启动子,通过在解脂耶氏酵母中过度表达来自P.dispersionUQ68J的蔗糖异构酶,获得了高水平的分泌蔗糖异构酶,其酶活力达到49.3 U/mL。
3.3 培养基和发酵条件优化
除启动子和信号肽优化外,发酵培养基组成和发酵条件的优化也是提高酶活性或表达水平的常用策略。培养基优化的重点是碳源、氮源和无机盐的组成和含量。刘军彤[21]将改造过的蔗糖异构酶克隆至宿主B.brevis中,并对该培养基氮源进行优化,当氮源为8.5 g/L的多聚蛋白胨和8.5 g/L的牛肉浸膏时,酶活力最高达138 U/mL,是优化前TM培养基的2.7倍;张贺朋等[40]对培养基中的蔗糖含量进行优化,当蔗糖含量为12%时,发酵液中蔗糖异构酶转化蔗糖的二糖产物与单糖副产物的比值达到92∶1;宋蕾等[27]对重组解脂耶氏酵母培养基的碳源、氮源、无机盐进行正交实验分析,确定最优条件为80 g/L果糖,2%含氮量的蛋白胨,0.5 mol/L NaCl,其酶活力达到251.96 U/mL,是优化前的2.9倍。发酵条件如溶氧量、发酵温度等也是影响蛋白表达以及催化产物组成的一大因素。刘军彤[21]对发酵过程中的搅拌速度进行优化,当转速为400 r/min时,蔗糖异构酶的酶活力提高到190 U/mL;邹亮[31]探究发酵温度对重组菌B.choshinensis/pNCMO2-SI在3 L发酵罐中的菌体生长浓度与酶活水平的影响时发现,30 ℃是最优温度,酶活力最高达到275 U/mL。
4 展望
目前,异麦芽酮糖作为一种安全的蔗糖替代品的市场需求迅速增长,蔗糖生物转化生产异麦芽糖已取得重大进展。然而,在工业生产中仍然存在一些问题,如食品安全级菌株的分泌水平低、热稳定性弱、蔗糖异构酶的应用性能差。为了解决上述问题,可以对表达宿主菌株和蔗糖异构酶编码基因分别进行改造。在表达宿主菌株改造方面:(1)敲除宿主细胞相关蛋白基因,防止外源蛋白被降解;(2)改造宿主细胞的细胞膜和细胞壁,提高外源蛋白转运效率;(3)对宿主细胞进行碳氮代谢调控优化、基因组精简等新型策略;(4)提高基因转录水平和稳定性。在蔗糖异构酶编码基因改造方面:(1)提高外源蛋白折叠效率;(2)利用蛋白质工程,对蔗糖异构酶进行定向进化或对编码基因如活性位点附近氨基酸残基、底物结合相关氨基酸残基进行定点突变,提高蔗糖异构酶的稳定性、酶活力、特异性等;(3)蔗糖异构酶的产物特异性不仅是保守序列RLDRD单独决定的,还存在多个调控区域的协同作用[7],需要进一步探索和修饰蔗糖异构酶的关键位点,提高蔗糖异构酶在工业生产中的应用性能。
