SYVN1泛素化降解XRCC5对晶状体上皮细胞DNA氧化损伤的影响△
2023-10-07王从玉李鹏飞王思文鲍思洁石海红管怀进
王从玉 李鹏飞 王思文 鲍思洁 石海红 管怀进
年龄相关性白内障(ARC)是全世界可逆性视力损害的罪魁祸首[1]。迄今,ARC的致病机制尚未完全阐明。因此,寻找非手术治疗策略对ARC患者具有重要意义。研究表明,紫外线(UVB)会导致晶状体上皮细胞(LECs)内DNA发生氧化损伤、蛋白质因错误折叠而累积。如果错误折叠蛋白未能及时降解,其异常聚集将导致晶状体混浊的发生[2-3]。在哺乳动物细胞中,泛素-蛋白酶体系统降解错误折叠或不需要的蛋白,参与多种神经退行性疾病的发展过程,包括阿尔茨海默病[4]、帕金森病[5]、亨廷顿病[6]等。泛素化是由三组酶(E1、E2和E3)催化的级联反应,E3泛素连接酶被认为是泛素化修饰中最重要的组成部分[7]。SYVN1是一种滑膜蛋白,已被证实在ARC的发病中起重要作用[8],然而对DNA氧化损伤的影响尚需进一步探索。XRCC5是非同源末端连接修复途径的重要基因,负责修复DNA双链断裂(DSB)[9]。最近的研究表明,DNA损伤修复蛋白(DRPs)泛素化是调节DNA损伤反应的关键机制,如果调控不当,DNA损伤反应可能会导致人类疾病[10]。因此,本研究用人晶状体上皮细胞系SRA01/04作为研究对象,探索SYVN1是否可以通过调节DRPs中XRCC5的泛素化降解影响LECs中DNA氧化损伤修复情况。
1 材料与方法
1.1 材料
SRA01/04细胞购于中国科学院上海生命科学研究所。胎牛血清、胰蛋白酶、DMEM培养液和细胞培养板购于美国Gibco公司;聚合酶链反应(qRT-PCR)实验所需耗材购于美国Thermo Scientific公司,逆转录试剂盒购于南京诺唯赞生物科技股份有限公司,引物购于中国生工生物工程股份有限公司;SYVN1过表达质粒购于中国PPL质粒与蛋白共享库;蛋白GAPDH抗体、荧光二抗(辣根过氧化物酶标记的二抗,山羊抗兔和山羊抗鼠IgG)购于中国ABclonal公司;SYVN1、XRCC5、Ub和γH2A抗体购于英国Abcam公司。
1.2 细胞培养
取出冻存的SRA01/04细胞,放入37 ℃培养箱中快速解冻,1 000~1 200 r·min-1离心10 min,加入完全培养基(含体积分数10%胎牛血清和10 g·L-1青链霉素的DMEM培养基)后,置于37 ℃、含体积分数5% CO2的细胞培养箱中培养。
1.3 UVB构建细胞氧化损伤模型
待细胞融合度达70%~80%时,用Hanks液清洗细胞1~2次,加入适量无菌PBS,在紫外灯(波长为275~400 nm,峰值为310 nm)下 30 cm 处照射15 min后[8],继续置于培养箱中培养24 h进行后续实验。
1.4 细胞转染及分组
SRA01/04细胞随机分为对照组、UVB组、UVB+HA组和UVB+OV-SYVN1组,其中,对照组细胞正常培养,UVB组细胞行UVB照射建立细胞氧化损伤模型,UVB+HA组和UVB+OV-SYVN1组细胞行细胞转染后再行UVB照射。细胞转染方法为:6孔板中SRA01/04细胞融合度达70%时,UVB+HA组和UVB+OV-SYVN1组细胞分别加入HA过表达质粒和SYVN1过表达质粒2 μg,转染试剂opti-MEM、Lipofectamine3000和P3000体积比为250 μL:75 μL:5 μL,室温孵育15 min;随后培养箱中孵育6 h,更换为含有胎牛血清的完全培养基,于培养箱中继续培养72 h,用于后续实验。
1.5 免疫沉淀实验
根据实验要求处理细胞后,加入免疫沉淀裂解液(100 mmol·L-1NaCl,20 mmol·L-1Tris-HCl,体积分数1% TritonX-100),提取蛋白质上清。随机分为IP组和Input组。其中,IP组分为IgG组、SYVN1组和XRCC5组,IP组进行免疫沉淀实验,用IgG抗体下拉与IgG结合的蛋白,用SYVN1抗体下拉与SYVN1结合的蛋白;Input组为未经免疫沉淀实验处理的细胞总蛋白。每组取400 μg蛋白质原液,分别加入1 μL对应的抗体后在摇床上孵育过夜。第2天,应用磁珠将抗体-蛋白质复合物提取出来,进行Western blot实验,检测SYVN1、XRCC5和泛素蛋白的表达。同时,本实验的Input组取50 μL蛋白原液,检测泛素蛋白的表达。实验重复3次。
1.6 免疫荧光染色检测DNA损伤标志物
在24孔板中放入细胞爬片,将细胞均匀接种在爬片上,培养6~8 h待细胞贴壁后,对细胞进行分组处理。孵育24 h后,用PBS清洗细胞。随后加入200 μL 40 g·L-1多聚甲醛,固定1 h;加入0.3 g·L-1牛血清白蛋白和体积分数0.5% Triton X-100溶液,室温封闭2 h;之后,根据抗体γH2A说明书加入荧光一抗(1:250),于4 ℃冰箱内孵育过夜;第2天,取出复温2 h后,PBST摇床洗3次,每次5 min;避光加入荧光二抗(1:500),室温孵育2 h 后,PBST摇床洗3次,每次5 min;用DAPI染核,避光孵育8 min后,PBST摇床洗3次,每次15 min;应用荧光显微镜观察并拍照。
1.7 Western blot实验检测蛋白相对表达量
将分组处理后的细胞用冰浴预冷的PBS清洗3次,每次5 min;之后,弃掉PBS,根据细胞密度加入80~200 μL蛋白质裂解液,冰上裂解10 min;用细胞刮刮下细胞,利用移液枪转移至1.5 mL EP管中,4 ℃ 摇床裂解30 min,4 ℃、14 000 r·min-1离心30 min,提取蛋白质上清。凝胶电泳(150 V)分离蛋白,应用湿转法转膜(250 mA,2 h)将靶蛋白转移至聚偏二氟乙烯(PVDF)膜上,使用5 g·L-1脱脂奶粉在室温下封闭2 h。参照目的抗体说明书稀释一抗(γH2A、SYVN1、XRCC5和泛素稀释比例均为1:1 000,GAPDH稀释比例1:10 000),加入PVDF膜中,置于4 ℃冰箱孵育过夜。第2天,用TBST漂洗3次,每次5 min,之后加入辣根过氧化物酶标记的山羊抗兔二抗(稀释比例为1:10 000),室温孵育2 h。最后加入显影剂,置于显影机内显影。以GAPDH作为内参,计算目的蛋白表达的灰度值。
1.8 UbiBrowser软件分析
利用UbiBrowser软件(http://ubibrowser.ncpsb.org.cn/v2/)预测能与XRCC5发生相互作用的E3泛素连接酶。
1.9 统计学方法
应用SPSS 22.0统计学软件分析数据。各组细胞的蛋白相对表达量及SYNV1转染效率对比均应用单因素方差分析,两样本间比较应用两样本t检验。检验水准:α=0.05。
2 结果
2.1 SRA01/04细胞中SYVN1过表达模型验证
Western blot实验结果显示,对照组、OV-SYVN1组细胞中SYVN1蛋白的相对表达量分别为1.00±0.15、1.99±0.17。与对照组比较,OV-SYVN1组细胞中SYVN1蛋白的相对表达量明显上调,差异具有统计学意义(P<0.01)(图1)。表明SYVN1过表达模型构建成功。
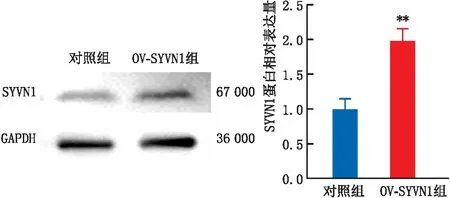
与对照组比较,**P<0.01。图1 Western blot实验检测SYVN1在SRA01/04细胞过表达模型中的表达变化
2.2 γH2A蛋白在SYVN1过表达模型中的表达
Western blot实验结果显示,对照组、UVB组、UVB+HA组和UVB+OV-SYVN1组中γH2A蛋白的相对表达量分别为1.00±0.04、1.59±0.04、1.53±0.07、1.85±0.04。与对照组比较,UVB组SRA01/04细胞中γH2A蛋白相对表达量明显升高,差异具有统计学意义(P<0.000 1);与UVB+HA组比较,UVB+OV-SYVN1组SRA01/04细胞中γH2A蛋白相对表达量明显升高,差异具有统计学意义(P<0.01)(图2)。表明过表达SYVN1会加重DNA氧化损伤。
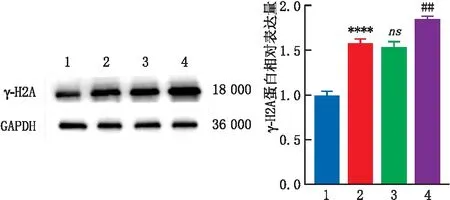
1:对照组;2:UVB组;3:UVB+HA组;4:UVB+OV-SYVN1组。与对照组比较,****P<0.000 1;与UVB+HA组比较,##P<0.01;ns表示与UVB组相比差异无统计学意义。图2 Western blot实验检测各组细胞γH2A蛋白表达
2.3 免疫荧光染色检测DNA氧化损伤程度
免疫荧光染色结果显示,与UVB+HA组比较,UVB+OV-SYVN1组SRA01/04细胞中γH2A染色明显加强(图3)。表明过表达SYVN1后,SRA01/04细胞的DNA氧化损伤程度加重。

图3 免疫荧光染色检测UVB+HA组和UVB+ OV-SYVN1组SRA01/04细胞中γH2A的表达
2.4 XRCC5存在泛素化修饰及其与E3泛素连接酶的预测
免疫沉淀实验发现,XRCC5存在泛素化修饰(图4A)。UbiBrowser软件预测发现, SYVN1可能是XRCC5的一个潜在的E3泛素连接酶(图4B)。
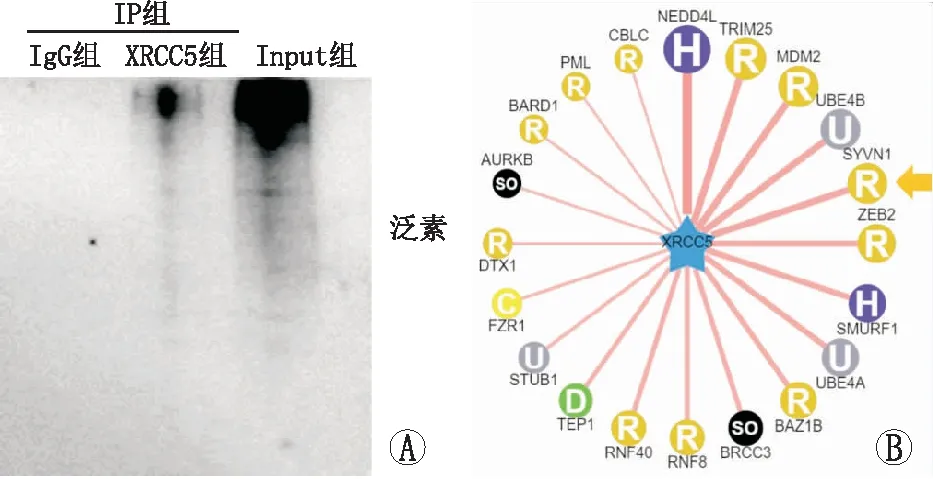
图4 免疫沉淀实验检测XRCC5存在泛素化修饰(图A)及UbiBrowser软件预测其相互作用蛋白(图B)
2.5 SYVN1与XRCC5之间存在相互作用
免疫沉淀实验结果显示,SYVN1与XRCC5存在相互作用(图5),表明SYVN1有可能参与介导XRCC5蛋白的泛素化修饰。

图5 免疫沉淀实验显示SYVN1与XRCC5存在相互作用
2.6 SYVN1促进XRCC5的蛋白质降解
Western blot实验结果显示,对照组和OV-SYVN1组中XRCC5蛋白相对表达量分别为1.00±0.12、0.33±0.05。与对照组比较,OV-SYVN1组中XRCC5蛋白相对表达量明显下降,差异具有统计学意义(P<0.001)(图6)。表明SYVN1可能通过促进泛素化修饰来加速XRCC5的蛋白质降解。
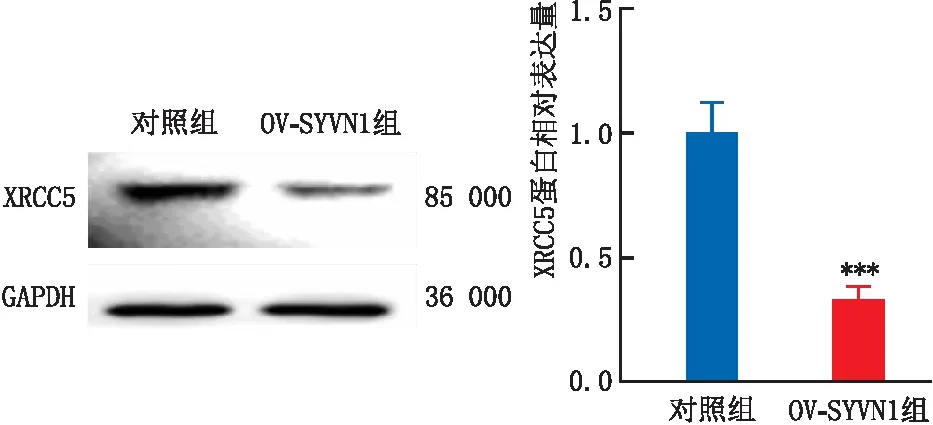
与对照组比较,***P<0.001。图6 Western blot实验检测XRCC5在OV-SYVN1氧化损伤细胞模型中的表达变化
3 讨论
ARC的发病机制仍不明确,但氧化应激诱导的DNA损伤是ARC形成和发展的关键因素[11-14]。而DNA氧化损伤后机体会启动DNA损伤反应系统,招募DRPs去修复受损的DNA。所以,探索DRPs表达改变的机制是目前研究ARC发病机制亟需解决的问题。本课题组前期研究发现,表观遗传修饰参与调控DNA损伤修复基因(DRGs)的表达,如ERCC6是核苷酸切除修复通路的关键酶,在ARC患者样本中其蛋白表达水平下降;研究还发现其启动子区存在高度甲基化,并被DNA甲基化酶DNMT3b所调控,导致ERCC6的转录程度降低,进而导致DNA修复进程不畅[15]。以上研究结果表明,DRGs的表达降低与ARC的致病过程紧密相关。所以,本课题组应用微阵列芯片检测96个DRGs在ARC中的表达变化,结果表明,仅存在10个DRGs表达明显下降,大多数DRGs的mRNA水平表达均无明显变化[16],提示可能存在其他的调控机制参与DRPs蛋白水平的表达。
泛素化修饰是蛋白质翻译后修饰之一,泛素化系统发生异常与衰老相关疾病的联系密切,包括年龄相关性白内障、癌症和神经退行性疾病[13]。此外,泛素化是由三组酶(E1、E2和E3)催化的级联反应,首先,E1泛素激活酶形成硫酯键激活泛素;随后,泛素被转移到E2泛素结合酶;最后,E3泛素连接酶催化泛素与目标蛋白的赖氨酸残基发生键合。E3泛素连接酶负责识别底物并提供底物特异性。因此,E3泛素连接酶被认为是泛素化修饰中最重要的组成部分[7]。近年来,E3泛素连接酶与ARC之间的联系引起广泛关注。Chen等[8]研究发现,E3泛素连接酶SYVN1介导MSH3泛素化降解,促进LECs凋亡,进而导致ARC进展。同样,李鹏飞等[17]研究发现,S期激酶相关蛋白2通过促进碱基切除修复蛋白8-氧代鸟嘌呤-DNA糖基化酶泛素化降解,导致LECs内部受损DNA的累积,从而加剧ARC发展。此外,泛素连接酶依据其结构域分为三类:HECT结构域、RING结构域和U-box结构域。SYVN1是RING结构域E3泛素连接酶家族成员,已经被证明在各种眼科疾病中发挥重要作用[18-19]。例如,Shruthi等[18]在糖尿病视网膜病变的大鼠中发现SYVN1可能会诱发内质网应激,在内质网应激持续作用下将引发神经元凋亡,进而导致DR进展。SYVN1对DNA氧化损伤的研究较少,已有学者证明在ARC患者的LECs中SYVN1表达升高[8]。为了探讨SYVN1在ARC患者中如何调控DNA氧化损伤,我们在氧化损伤模型中构建了SYVN1过表达模型,检测DNA损伤标志物γH2A的表达变化,由于γH2A含量与DNA受损严重程度呈正比,因此其被当作DNA损伤重要标志物之一。本研究结果表明,过表达SYVN1会加重LECs中DNA损伤,但具体机制尚不清楚。
为了进一步讨论SYVN1如何影响DNA氧化损伤修复,我们进行了生物信息学分析,预测发现可能与XRCC5蛋白发生相互作用的E3泛素连接酶,并通过免疫沉淀实验证明SYVN1与XRCC5之间存在相互作用。晶状体中存在多条DNA氧化损伤修复通路[20],DSB修复途径是DNA复制过程中的主要修复途径,经过非同源末端连接或同源重组的形式修复断裂的DNA双链,维持染色体稳定性[21]。XRCC5是DSB修复通路的主要蛋白,其表达下降将会导致DNA修复功能障碍、细胞周期停滞甚至凋亡,并诱发癌症等重大疾病[22]。有学者研究表明,蛋白质翻译后修饰可能调控DNA损伤修复基因表达。因此,及时有效地修复受损DNA对预防ARC具有重要意义。在之前的研究中,我们在ARC患者中检测到XRCC5蛋白表达水平下降,并表明XRCC5可能对LECs诱导的凋亡具有保护作用[22],提示我们在LECs中继续探索XRCC5基因表达下降的机制。在前期研究和本研究中我们均发现,XRCC5能发生泛素化修饰,但具体调控机制尚不清楚。尽管Mao等[22]研究发现,在ARC患者中Trim25表达升高,泛素化降解XRCC5导致LECs凋亡,可能与ARC的发病机制有关,但是本研究拓展了XRCC5潜在的调控机制。进一步研究发现,过表达E3泛素连接酶SYVN1能够显著下调XRCC5的表达,提示SYVN1可能泛素化降解XRCC5,参与ARC内的DNA损伤修复过程。
4 结论
SYVN1能够介导DRGs中XRCC5发生泛素化修饰,并通过降解XRCC5蛋白表达进而影响DNA氧化损伤修复过程,参与ARC的发展,这些结果提示DRPs的翻译后修饰可能与ARC的发病机制相关。本研究为ARC的发病机制提供了新见解。
