基于线粒体COⅠ基因的河南省境内锯齿新米虾种群遗传多样性研究
2023-10-07马志刚冯梦霞汤永涛刘如垚闫雪朦胡博聂国兴周传江
马志刚,冯梦霞,汤永涛,刘如垚,闫雪朦,胡博,聂国兴,周传江,
(1. 河南师范大学水产学院,河南省水产动物养殖工程技术研究中心,水产动物疾病控制河南省工程实验室,河南新乡 453007;2. 河南师范大学生命科学学院,河南新乡 453007)
锯齿新米虾Neocaridina denticulata是一种体型偏小的淡水虾类,对水域环境适应能力较强,但分布范围较狭窄,据记载,其在国内仅分布于太湖区、福建等特定区域,后被移养至新疆、巴楚等地的部分区域(梁象秋,2004),在新米虾属Neocaridina的修订中首次系统、详细地描述了新米虾属的分类地位及其形态特征,为相关研究提供了理论依据。随着更多物种的发现和分类学研究的深入(蔡奕雄,1996;梁象秋等,1999;梁象秋,2004),锯齿新米虾、大卫米虾Caridina davidi和异足新米虾N. heteropoda的分类命名存在一定冲突和争议,Klotz 等(2013)的研究认为,大卫米虾、锯齿新米虾大卫亚种N. denticula davidi、锯齿新米虾中华亚种N. denticula sinensis是异足新米虾的同物异名,其有效物种名为N. davidii。
锯齿新米虾作为一种形态体色多变的常见小型淡水虾,常被用作观赏虾类进行交易和养殖。为了培育更多体色增加其观赏价值,米虾属Caridina和新米虾属中许多种被人工杂交和选育(林师,2017)。此外,锯齿新米虾的环境适应性较强,有学者对其低溶解氧胁迫的耐受能力、水体污染的生态响应进行研究报道(李典宝,2015;薛毓才等,2022),为后续锯齿新米虾环境适应相关研究奠定了基础。
河南省地处中原腹地,地跨长江、淮河、黄河、海河4 大流域,境内有1 500 多条主干河流纵横交错,水系资源丰富,地形复杂。对河南省境内的虾类资源调查结果显示,相较于其他淡水虾类而言,锯齿新米虾在河南境内4大水系均有分布,且数量较大,但缺乏遗传多样性的研究报道。目前,DNA分子标记是进行群体遗传多样性研究的主要手段之一,线粒体COⅠ基因已被广泛应用于鱼类、虾类种群遗传多样性的研究(冯建彬等,2008;公月月,2017)。本研究基于线粒体COⅠ基因对河南境内19 个野生锯齿新米虾群体的149 尾样本进行遗传多样性分析,评估河南境内锯齿新米虾种质资源现状,为其种质资源保护、可持续发展等提供理论参考。
1 材料与方法
1.1 实验材料
2012—2020 年于河南省4 大水系的干流及其部分支流中采集锯齿新米虾,标本浸泡于95%乙醇中带回实验室保存。选取其中19 个地理群体149尾样本作为研究对象(表1)。
1.2 COⅠ基因的获取
取适量样本肌肉组织(约0.01 g),采取酚-氯仿法提取DNA(Sambrooket al.,1989)。用1%琼脂糖凝胶电泳和Nanodrop 微量紫外分光光度计分别检测基因组DNA的质量与浓度,-20 ℃保存备用。
扩增线粒体COⅠ基因引物为LCO1490:5’-GGTCAACAAATCATAAAGATATTGG-3’,HCO21 98:5’-TAAACTTCAGGGTGACCAAAAAATCA-3’(Folmeret al.,1994),引物由生工生物工程(上海)股份有限公司合成。PCR 反应体系为30 µL:2×Es Taq Master Mix 15 µL,正、反向引物各1.5 µL,DNA模板1 µL,无菌双蒸水11 µL。PCR 反应程序为:94 ℃ 5 min;94 ℃ 30 s,54 ℃ 45 s,72 ℃ 45 s,35 个循环;72 ℃ 7 min,4 ℃保存(冯建彬等,2008)。PCR 产物经1%琼脂糖凝胶电泳检测后送至生工生物工程(上海)股份有限公司进行测序。
1.3 数据分析
测序获得的DNA序列采用Lasergene 7.1的SeqMan(Swindell & Plasterer,1997)进行人工核查、校正及组装拼接。拼接完成的序列用BioEdit(Tippmann,2004)进行多重比对,获得一致序列备用。用DnaSP 5.0(Librado & Rozas,2009)计算DNA 序列的单倍型数、单倍型多样性(Hd)和核苷酸多样性(π)等遗传多样性参数。用MEGA 7.0(Sudhiret al.,2016)计算序列碱基组成、变异位点、简约信息位点和组内、组间遗传距离等参数。
用Arlequin 3.5(Excoffieret al.,2005)进 行AMOVA 估算遗传变异在不同水系间的分布和遗传分化系数(FST),进而评估不同水系锯齿新米虾群体的遗传分化程度;为探究其种群历史动态,进行Fu’sFs和Tajima’s D 中性检验及错配分布分析,评估群体是否发生种群扩张。根据长臂虾科Palaemonidae 的系统发育关系,以细足米虾C. nilotica gracilipes(GenBank 登录号:MK190025)为外群(李新正等,2003),Bootstrap 设置为1 000检验置信度,构建锯齿新米虾19 个群体的ML 系统发育树,以分析各群体之间系统进化关系及地理分化情况。采用Network 5.0(Bandeltet al.,2000)中的Median-Joining 法构建锯齿新米虾的单倍型网络图,以分析其单倍型演化关系。用BEAST 1.7.4(Heled & Drummond,2010)进行贝叶斯天际线点(Bayesian skyline plot,BSP)分析估算锯齿新米虾种群历史动态的变化,采用1.4%COⅠ基因进化速率(Knowlton & Weigt,1998),由Tracer 1.5(Rambaut & Drummond,2009)生成BSP图。
2 结果与分析
2.1 COⅠ基因序列基本特征
19个地理群体149条锯齿新米虾的COⅠ基因序列长632 bp,其中包括变异位点43 个(6.8%),简约信息位点32 个,单变异位点11 个,转颠换位点数的比值为2.628∶1;T、C、A 和G 的平均含量分别为33.4%、20.7%、27.2%和18.7%,表现出显著的AT偏倚性。
2.2 种群遗传多样性
锯齿新米虾样品共包含35 个单倍型,其中,24 个为独享单倍型,11 个为群体间共享,Hap6被长江、黄河、淮河10 个群体33 尾个体共享。各 水系 的Hd为0.773~0.924,平 均0.922,其中,淮河水系的最低(Hd=0.773),长江水系的最高(Hd=0.924);各水系的π 为0.002 28~0.006 37,平均为0.006 32,其中,淮河水系的最低(π=0.002 28),长江水系的最高(π=0.006 37)。整体上,长江水系群体显示出更高的遗传多样性(Hd=0.924,π=0.006 37),远高于其他3 个水系(表2)。

表2 河南省4大水系锯齿新米虾群体的遗传多样性信息Table 2 Genetic diversity of Neocaridina denticulata populations from four river systems in Henan Province
2.3 种群遗传结构
4 大水系群体内遗传距离为0.002 29~0.006 45,其中,淮河水系的最小(0.002 29),长江水系的最大(0.006 45);两两水系间的遗传距离为0.004 71~0.008 87,其中,长江水系与淮河水系的最小(0.004 71),海河水系与长江水系的最大(0.008 87)。4 大水系间的FST为0.104 68~0.586 00,4 大水系之间均存在显著的遗传差异:黄河水系和长江水系的最高(0.586 00),且存在极显著遗传差异;黄河水系与海河水系的最低(0.104 68),且存在显著差异(表3)。
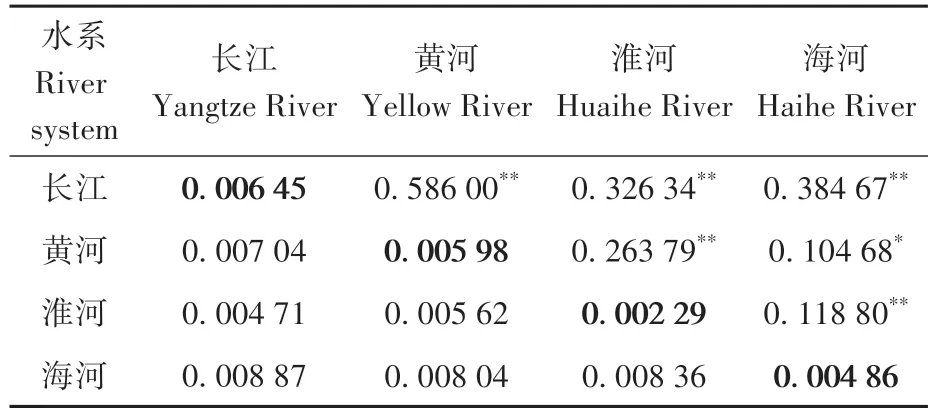
表3 河南省4大水系锯齿新米虾群体遗传结构Table 3 Genetic structure of Neocaridina denticulata populations from four river systems in Henan Province
海河水系中,E02 与C02、E05,E04 与C02 的遗传分化较低(FST<0.25),其余群体与所有群体的遗传分化程度均较高(FST>0.25),且分化程度均达到显著或极显著水平;海河水系的5个群体之间也存在一定遗传差异,除E02与E04、E05之间的遗传分化程度没有达到显著差异水平外,其他几个群体之间的遗传分化程度均达到了显著或极显著水平。FST分析与成对遗传距离分析结果一致,均表明C02、E01、E02 和E04 群体与其他各锯齿新米虾群体间存在较大遗传分化(表4)。

表4 河南省4大水系锯齿新米虾群体遗传结构Table 4 Genetic structure of Neocaridina denticulata populations from four river systems in Henan Province
AMOVA 结果显示,锯齿新米虾的遗传变异主要源自其群体内的个体间,且群体内锯齿新米虾个体间的变异占总变异的38.10%,群体间的变异则占35.71%(表5)。

表5 河南省4大水系锯齿新米虾群体AMOVA分析Table 5 Analysis of molecular variance among Neocaridina denticulata populations from four river systems in Henan Province
锯齿新米虾19个群体的ML系统发育树(图1:a)显示,各水系不同群体间系统进化关系成交叉式嵌套,无明显水系和地理分化。单倍型网络图显示,锯齿新米虾群体各水系35 个单倍型之间总体上并未呈现出明显的水系交叉,除Hap1、Hap4、Hap6存在不同水系锯齿新米虾样品之间单倍型共享的情况外,其他各个锯齿新米虾群体形成了比较明显的地理结构,不同水系的群体间多为特有单倍型;此外,35 个单倍型中,除Hap1、Hap27 和Hap29 拓展出少量的单倍型外,Hap6 的拓展单倍型最多(图1:b)。
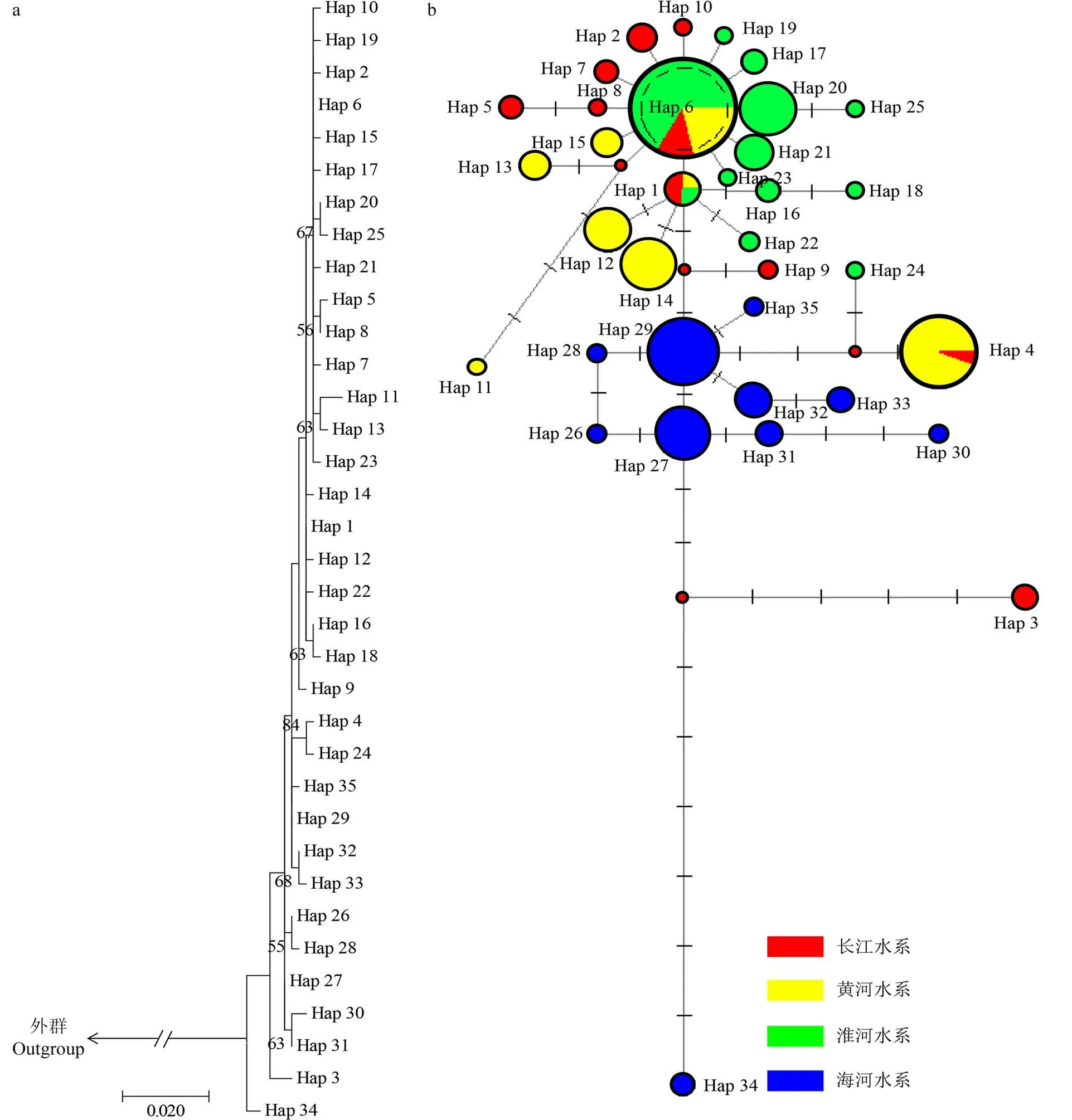
图1 基于COⅠ基因构建的河南境内4大水系锯齿新米虾的ML系统进化树(a)及单倍型网络图(b)Fig. 1 ML phylogenetic tree (a) and network of haplotypes (b) of Neocaridina denticulata from four river systems in Henan Province based on COⅠ gene sequences
2.4 种群历史动态
河南省锯齿新米虾群体的Tajima’sD检验值和Fu’sFS检验值均为负数(Tajima’sD=-1.692 13,Fu’sFS=-1.167 07),均未达到显著差异水平(表2);错配分布分析表明,群体的错配分布曲线呈现出多峰状态(图2),推测河南省锯齿新米虾种群相对稳定,近期未发生过种群扩张事件。BSP分析结果显示,锯齿新米虾群体整体保持稳定趋势,并未经历种群扩张或瓶颈效应(图3)。

图2 河南境内4大水系锯齿新米虾群体错配分布分析Fig. 2 Mismatch distribution analysis of Neocaridina denticulata populations from four river systems in Henan Province
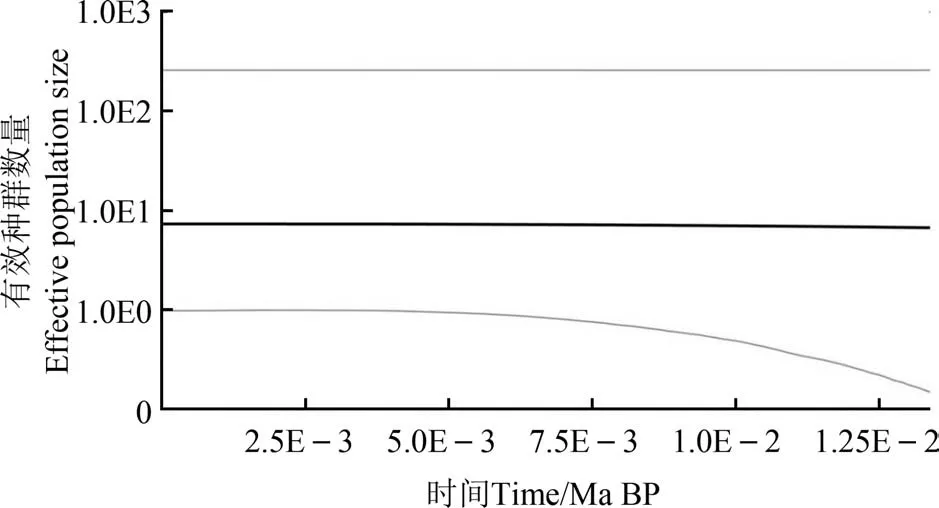
图3 河南境内4大水系锯齿新米虾群体总贝叶斯天际线点图Fig. 3 Bayesian skyline plots of Neocaridina denticulata populations from four river systems in Henan Province
3 讨论
3.1 河南境内锯齿新米虾的遗传多样性
采集的河南境内149 尾锯齿新米虾群体平均遗传多样性指数(Hd=0.922,π=0.006 32)均低于河南境内日本沼虾M. nipponense群体(Hd=0.785 74,π=0.010 59),但是略高于掌肢新米虾N. palmata palmata群体(Hd=0.557±0.025,π=0.000 98±0.000 07)(郑小壮等,2019)。总体而言,河南境内锯齿新米虾与河南境内日本沼虾群体、掌肢新米虾(郑小壮等,2019)、4 个海域口虾蛄Oratosqilla oratoria群体(董鑫等,2015)及紫贻贝Mytilus galloprovincialis群体(沈玉帮等,2011)均表现出单倍型多样性较高、核苷酸多样性较低的特点。
遗传多样性分析结果显示,长江水系锯齿新米虾群体显示出更高的遗传多样性(Hd=0.924,π=0.006 37),均远高于其他3 个水系,表明长江水系锯齿新米虾群体目前具有较高的遗传多样性,这与河南境内日本沼虾群体的遗传多样性现状(郑小壮等,2019)相一致。而淮河水系和黄河水系因采样点多为半封闭水体,群体间可能存在基因交流,导致结果偏低。
3.2 河南境内锯齿新米虾的遗传结构
淮河水系的遗传距离最小,长江水系与其他水系之间的遗传距离均较大。其次是黄河水系和海河水系,海河遗传距离也相对较大。FST值差异显著性检验显示,海河水系与其他各水系的群体均存在显著或极显著的遗传差异(P<0.05,P<0.01),达到高度遗传分化(FST>0.25),就海河水系内部来看,其5个群体之间也存在一定遗传学差异。或因海河水系涉及3 个采样点(E02、E04、E05)为水库,其他2个采样点则位于林州市太行山脉中,与其他几个地理群体存在一定地理隔离和基因交流障碍,从而导致了该水系锯齿新米虾群体与其他群体产生了差异。
单倍型Hap6 在10 个种群中共享,且以其为中心直接拓展出10个单倍型,呈明显的星形辐射状,占据除海河以外的其他3个水系,为分布最广泛的单倍型,推断Hap6 是一个存在时间较为久远的主要单倍型。
3.3 河南境内锯齿新米虾的种群历史动态
中性检测结果显示,河南境内锯齿新米虾群体的平均Fu’Fs值和Tajima’sD值均为负值但未达显著差异水平;错配分布分析显示其群体错配分布图为多峰泊松分布;BSP分析结果显示锯齿新米虾群体整体上保持稳定趋势。推测河南境内锯齿新米虾种群整体比较稳定,未经历过种群扩张及瓶颈效应。
锯齿新米虾作为一类古老的小型淡水虾类,在我国除了分布于几个已知的区域(梁象秋,2004)外,在贵州(邓一德,吴本寿,1997)、江西(李文浩等,2016)、湖北三峡(杨振冰等,2018)等多个地方记录有分布。锯齿新米虾不同地理群体间的遗传距离虽然较小,但多存在单独的遗传结构和一定的地理格局,推测其原因可能有以下几点:首先,采集样点的设置和采样方法的不同均会对分析结果产生一定的影响,如水库等静水与溪流等流水的群体可能存在因为生存环境的不同和基因交流的阻碍而产生遗传分化现象;其次,由于锯齿新米虾个体较小,且大多生活在静水或水体流动性较小的水体,迁移能力相对较弱,因此,不同水系锯齿新米虾群体之间可能因为地理、水系上的隔离而存在一定的交流障碍,从而造成群体之间的分化;最后,气候和环境等不同也会引起其局部适应性的改变,捕捞、放流等人为因素造成锯齿新米虾不同地理群体及其栖息环境之间存在一定的差异。为了适应不同的水域环境,不同群体之间可能会发生一定程度的进化或变异(易啸等,2018;郑小壮等,2019)。
本研究对河南省境内锯齿新米虾遗传多样性进行了系统研究,对进一步评价河南省淡水虾类物种多样性和淡水虾类广布种的遗传多样性现状,丰富河南省境内关于淡水虾类的相关研究,完善境内淡水虾类的物种名录和后续相关研究提供了理论依据和参考。
