贵州花溪大学城破碎化林地蝴蝶多样性与嵌套分布格局
2023-10-07伍素李义杨开慧李琳曾维波蔡明飞刘宇张凯徐雨
伍素,李义,杨开慧,李琳,曾维波,蔡明飞,刘宇,张凯,徐雨
(贵州师范大学生命科学学院,贵阳 550025)
生境破碎化是指在人为活动或自然干扰作用下,大片连续分布的自然生境被其他非适宜生境分隔成许多面积较小斑块的过程(Fahrig,2003)。如水位上升导致部分陆地与大陆隔离而形成陆桥岛屿(Wanget al.,2010;Huet al.,2011),森林砍伐后形成的片断化森林斑块(Hillet al.,2011),城镇化过程中道路、房屋修建对原生生境破坏和分割后形成的斑块(Hillet al.,2011;郑进凤等,2021)。目前,已有大量研究分析了生境破碎化对生物多样性的影响,认为生境破碎化是导致生物多样性减少的重要因素之一(武晶,刘志民,2014;吴倩倩等,2017;Miller-Rushinget al.,2019)。
在众多研究中,基于岛屿生物地理学理论提出的面积效应、隔离效应是最基础的研究内容。面积效应、隔离效应从物种丰富度角度预测:随着生境被分割、缩小和隔离,其中的物种丰富度可能降低(Watling & Donnelly,2006)。在此基础上,一些学者开始从群落组成角度研究生境破碎化的影响,报道破碎化生境中的生物群落呈现嵌套格局,即小斑块中的物种是大斑块中物种集合的子集(Wanget al.,2010)。如浙江千岛湖陆桥岛屿中的大中型兽类、鸟类和两栖爬行类(张竟成等,2008;王熙等,2012;Chenet al.,2019),广西喀斯特破碎化森林斑块中的鸟类(Liet al.,2013),浙江杭州城市绿地中鸟类呈现明显的嵌套格局(Wanget al.,2013)。嵌套格局的产生可能与生境被分割、缩小和隔离过程中的物种选择性灭绝(Patterson & Atmar,1986)、生境嵌套(Blake,1991)、选择性迁入(Cook & Quinn,1995)等因素有关。但是,一些研究也指出,受竞争作用、环境过滤和生境异质性等因素的影响,破碎化生境中的一些动物群落不存在嵌套格局,甚至出现反嵌套格局(Chenet al.,2018;郑进凤等,2021;田璐嘉等,2022)。
蝴蝶是一类体型较大、易观察和辨别的昆虫类群。它们虽然有一定的飞行能力,但幼虫以植物为食,对寄主植物较为专一,对生境恶化做出反应的速度比其寄主植物快(张立微,张红玉,2016)。在生境丧失和破碎化时,蝴蝶的生存和繁殖会受到较大的影响。因此,在生物多样性监测与评价中,蝴蝶常被作为主要的指示生物类群之一(Chenet al.,2021)。当前,一些学者以海洋性群岛(英国和爱尔兰大陆附近的岛屿、中国舟山群岛)为样地,分析了蝴蝶多样性的种面积关系和嵌套分布格局,揭示随着岛屿面积减小、隔离度增加,受选择性灭绝和选择性迁入等因素的影响,蝴蝶群落分布呈现嵌套格局(Denniset al.,2012;Xuet al.,2017)。但是,有关破碎化生境中的蝴蝶多样性与群落分布的研究案例还比较缺乏(Mac-Donaldet al.,2018)。
本研究以贵州省花溪大学城破碎化自然林地为研究样地,对蝴蝶多样性进行调查,研究斑块面积、隔离度与物种丰富度的关系,并分析群落是否存在嵌套格局,以此丰富生境破碎化研究理论,并为城市规划建设和蝴蝶多样性保护提供科学依据。
1 研究方法
1.1 研究区概况
研究区位于贵州省贵安新区东南部,紧邻贵阳市花溪区(106°36′~106°40′E,26°22′~26°24′N),面积约10 km2。研究区属于典型的喀斯特地貌特征,海拔1 000~1 200 m,属亚热带湿润温和型气候,年均温15 ℃,年均降水量1 130 mm。该区域在规划修建大学城以前,为农田环绕的自然林地。2009 年大学城动工修建至投入使用,自然林地被人工设施、建筑物、道路等不透水面分割。残留林地斑块中的植被类型相对简单,主要由暖性松林、常绿落叶阔叶林、落叶阔叶灌丛和禾草灌草丛组成,受喀斯特地貌特征影响,不同斑块间的小生境尤其是植物种类存在较大差异(曹朝阳,2021)。本研究选择其中26 个斑块作为研究样地(面积0.3~290.4 hm2;表1)。
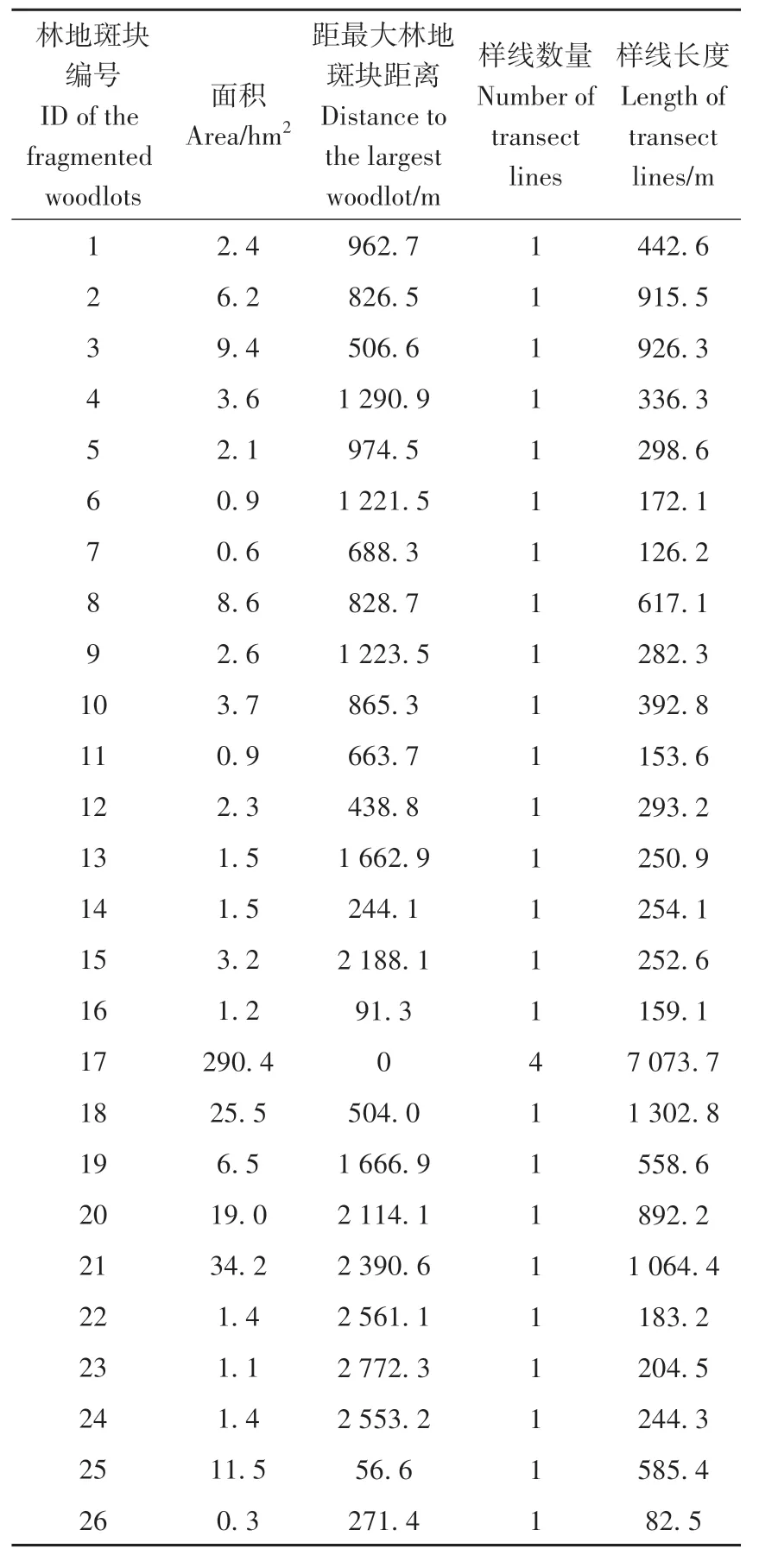
表1 贵州花溪大学城26个破碎化林地斑块的地理特征、样线数量和长度Table 1 Characteristics of 26 fragmented woodlots and number and length of transect lines recorded in the Huaxi University Town,Guizhou
1.2 斑块地理参数的测量
从Google Earth 下载研究区的卫星地图,导入ArcGIS 10.2 中勾绘出各个斑块的边界,采用“计算几何”的方式计算斑块面积,以“生成近邻表”的方式计算各斑块到最大斑块的距离,以反映斑块的生境破碎化程度。
1.3 蝴蝶采集及鉴定
参考Schoereder 等(2004)的研究,在不同斑块中设置长度与面积成正比的固定样线。于2017—2022 年的5—8 月分别对每个斑块进行了6 次蝴蝶调查与样品采集,每条样线每次调查1次。选择天气晴朗、蝴蝶活跃度较高的时间段(10∶00—17∶00)采用扫网法捕捉蝴蝶。每条样线2 名调查人员,调查行进速度为10 m/min;用捕虫网捕捉样线两侧5 m 范围内的蝴蝶,将采集到的个体置于硫酸纸三角袋中,记录采集日期、地点和采集人等相关信息(张雪梅等,2016;Xuet al.,2017)。
物种鉴定参考《中国蝴蝶分类与鉴定》(周尧,1998)、《中国昆虫生态大图鉴》(张巍巍,李元胜,2011)、《贵州蝴蝶》(陈正军,2016)和《中国蝴蝶图鉴》(武春生,徐堉峰,2017)。
对形态鉴定存疑的种类,利用DNA 条形码进行分类(Hebertet al.,2003)。提取蝴蝶腿部总DNA,扩增线粒体COⅠ基因的部分片段(658 bp)测定序列,后与BOLD 数据库(http://www.boldsystems.org/)进行比对和鉴定。
1.4 统计与分析
(1)物种丰富度:分别统计26个斑块中的蝴蝶物种数。
(2)计算Berger-Parker 优势度指数(d)(Berger& Parker,1970):d=ni/N,式中,ni为物种i的个体数量,N为全部物种的总个体数量。d≥0.05 为优势种,0.005≤d<0.05 为常见种,d<0.005 为少见种或偶见种。
(3)建立线性回归模型:按照岛屿生物地理学理论提出的种面积函数公式,对物种丰富度、斑块面积、距最大斑块距离采取10 为底数的对数转换(由于最大斑块到自身的距离为0,因此在对数转换前对各个距离值进行+1 处理)。利用R.4.2.1 构建26 个斑块中蝴蝶物种丰富度与斑块面积、距最大林地斑块距离的线性回归模型,分析是否具有面积效应和隔离效应。使用visreg 程序包绘制模型的偏残差图(Breheny & Burchett,2020)。
(4)分析群落嵌套格局:构建斑块与蝴蝶物种多度矩阵,分析群落的嵌套格局(Almeida-Neto &Ulrich,2011)。使用NODF2.0 中的WNODF 方法计算物种(WNODFr)、斑块(WNODFc)和整体的嵌套值(Almeida-Neto & Ulrich,2011)。设置行列固定(rc)、总数固定(aa)和总物种丰富度固定(ss)等3 种重采样零模型,依据软件随机产生1 000 个矩阵对嵌套度进行显著性检验。
上述统计分析中,P<0.05表示差异显著。
2 结果
共采集蝴蝶个体586 只,属于5 科51 属81 种(https://figshare.com/s/099c3ebb67588dbe57e0)。其中,蛱蝶科Nymphalidae 物种数最多(32 种),其次是灰蝶科Lycaenidae(18 种)、凤蝶科Papilionidae(13 种)、粉蝶科Pieridae(12 种)和弄蝶科Hesperiidae(6 种)。优势种有4 种,分别是东方菜粉蝶Pieris canidia、东亚豆粉蝶Colias poliographus、琉璃灰蝶Celastrinasp.和酢浆灰蝶Zizeeria maha。常见种44种,少见种或偶见种33种。
不同林地斑块中的蝴蝶物种数为2~53,平均为10.2 种±9.7 种。线性回归分析表明,蝴蝶物种丰富度与林地斑块面积呈边缘显著正相关(β=0.149,P=0.057);蝴蝶物种丰富度与距最大林地斑块距离呈显著负相关(β=-0.201,P=0.007;图1)。

图1 贵州花溪大学城26个破碎化林地中蝴蝶物种丰富度与林地斑块面积和距最大斑块距离的线性回归偏残差Fig. 1 Partial residual for linear regressions of between the species richness and the woodlot area and the distance to the largest woodlot in the 26 fragmented woodlots in the Huaxi University Town,Guizhou
3 种重采样零模型的WNODF 嵌套分析显示,与随机产生的1 000 个分布矩阵相比,rc 模型和aa模型表明没有显著嵌套格局,ss模型表明存在反嵌套格局(表2)。
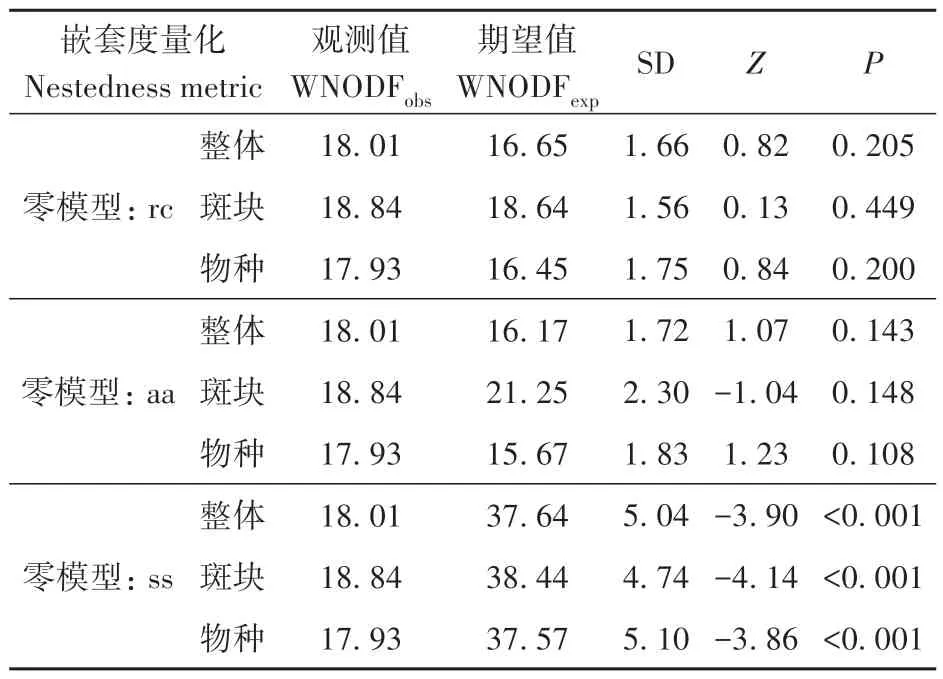
表2 贵州花溪大学城26个破碎化林地中蝴蝶物种多度矩阵的嵌套格局分析Table 2 Nestedness analyses for species-by-sites abundance matrix of butterflies recorded in 26 fragmented woodlots in the Huaxi University Town,Guizhou
3 讨论
本研究在贵州花溪大学城26个破碎化自然林地中记录到蝴蝶81 种,数据分析表明蝴蝶物种丰富度与林地斑块的面积呈边缘显著正相关,符合岛屿生物地理学理论提出的“面积效应”(McArthur & Wilson,1967)。由于土地大面积开发,贵州省花溪大学城的原生植被正不断被房屋、道路等人工建设物分割。斑块面积在一定程度上反映了可供物种利用的空间分布范围与资源量;小斑块中的生境丰富度和资源量可能较低,难以满足一些物种的生存需求,因此蝴蝶物种丰富度较低(张雪 梅等,2016;Xuet al.,2017;MacDonaldet al.,2018)。蝴蝶物种丰富度与距最大斑块距离呈显著负相关,表明存在隔离效应。虽然本研究区内各林地斑块的隔离度较低,但蝴蝶的飞行或扩散能力较弱,且人流量大和公路行驶车辆多等一定程度上增强了隔离效应。
不同林地斑块间蝴蝶群落未呈现出嵌套结构,甚至可能出现反嵌套格局。这一结果与一些海洋群岛中蝴蝶群落的研究结果不同(Denniset al.,2012;张雪梅等,2016;Xuet al.,2017),也不同于一些破碎化城市生境中其他动物群落的研究结果(González-Orejaet al.,2012;Wanget al.,2013;de la Hera,2019)。一般认为,嵌套格局的产生与选择性灭绝(Patterson & Atmar,1986)、生境嵌套(Blake,1991)和选择性迁入(Cook & Quinn,1995)等因素有关。选择性灭绝理论认为,物种分布与斑块面积密切相关,一些种群较小、最小需求面积较大的物种通常具有较高的灭绝风险,会率先从小斑块中消失。选择性迁入理论认为物种的分布与其扩散能力有关,一些扩散能力较弱的物种可能会有序从隔离度较大的斑块中消失。生境嵌套理论认为,物种的空间分布与生境结构密切相关,斑块间的生境呈嵌套分布将导致物种的嵌套分布。虽然本研究揭示蝴蝶物种丰富度随林地斑块面积减小、隔离度增加而降低,但喀斯特特殊的地质地貌造成的局部小生境和植物种类差异(曹朝阳,2021;郑进凤等,2021;朱芸等,2022),以及蝴蝶对寄主植物的较强选择性(Krämeret al.,2012),可能使物种出现在各自适应的特定生境中,从而削弱了嵌套格局,甚至可能导致反嵌套格局(Gutiérrez-Cánovaset al.,2013;郑进凤等,2021)。
综上所述,本研究以贵州省花溪大学城为例,揭示了生境破碎化影响蝴蝶物种丰富度,即斑块面积越小,蝴蝶物种丰富度越低。但是,不同林地斑块间的蝴蝶群落没有由此产生显著的嵌套格局,甚至可能呈现反嵌套格局,这可能与喀斯特生境异质性造成的斑块间物种组成差异较大等因素有关,具体机制有待进一步研究。基于本研究结果,建议在城市规划建设时应注重维持栖息地的完整性,对不同面积的林地斑块均加以保护。
致谢:感谢申力欣、张秋秋、高丽桦、张健、殷枭、梁晓珊、罗叶、李洁、朱芸在野外调查和蝴蝶形态鉴定工作中付出的辛劳,感谢汪小芳、赵娟在DNA 条形码技术中所付出的努力。
