四川发现云开脊蛇
2023-10-07张佳琦李科王朝英吴亚勇舒国成时磊
张佳琦 ,李科,王朝英,吴亚勇,舒国成 ,时磊
(1. 新疆农业大学,乌鲁木齐 830052;2. 宜宾学院,四川宜宾 644007)
脊 蛇属AchalinusPeters,1869,是闪 皮蛇 科Xenodermidae 物种数量最为丰富的类群,目前已描述物种22 种,其中我国记录有16 种(Peters,1869;赵尔宓,2006;Liet al.,2021;Yanget al.,2022)。近些年,随着分子技术的发展,脊蛇属物种多样性得到了更深入的研究,但多数物种的研究仅基于个别采样地或少数个体,大多数物种的具体分布区及种下关系并不明晰(Wanget al.,2019;Houet al.,2021;Liet al.,2021;Yanget al.,2022),如近年新发表 的云 开脊 蛇A. yunkaiensisWang,Li & Wang,2019、越北脊蛇A. emilyaeZiegler,Nguyen,Pham,Nguyen,Pham,Van Schingen,Nguyen & Le,2019(Van Phamet al.,2019)和杨氏脊蛇A. yangdatongiHou,Wang,Guo,Chen,Yuan & Che,2021的新分布区陆续被报道(喻敏等,2020;Houet al.,2021)。我国是脊蛇属物种的主要分布区,其中四川省已记录脊蛇属物种3 种:黑脊蛇A. spinalisPeters,1869、美姑脊蛇A. meiguensisHu & Zhao,1966和攀枝花脊蛇A. panzhihuaensisHou,Wang,Guo,Chen,Yuan & Che,2021(赵尔宓,2006;Houet al.,2021)。
2022年6月,在四川省泸州市画稿溪国家级自然保护区开展动物学野外实习期间采集到2 号蛇类标本,通过形态学比较和分子遗传关系分析,确定为云开脊蛇,系四川省蛇类分布新记录种。
1 材料与方法
1.1 实验材料
共采集到1号雌性亚成体标本(YBU 22050)和1号雄性幼体标本(YBU 22148)。在拍摄完照片后经安乐死取得肝脏等组织,组织置于95%乙醇溶液,后转存于-80 ℃超低温冰箱;标本先经10%甲醛溶液处理,待标本定型后保存于80%乙醇溶液。所有组织及标本均保存于宜宾学院(YBU)的动物多样性与生态保育宜宾市重点实验室。
1.2 形态鉴定
形态测量及描述依据赵尔宓等(1998)、赵尔宓(2006)、Wang等(2019)、Li K等(2020,2021)和喻敏等(2020)。包括长度测量、鳞片计数及色斑描述。使用软尺测量尾长(tail length,TaL)、头体长(snout-vent length,SVL)和总长(total length,TL)(精度1 mm);利用数显游标卡尺测量鳞片长度:前额鳞沟(length of suture between the prefrontals,LSBP)和鼻间鳞沟(length of suture between internasals,LSBI)、颊鳞的长度(length of loreal,LeL)、颊鳞的高度(high of loreal,HiL)、前1枚鼻鳞的长度(length of anterior section of nasal,LaSN)和后1 枚鼻鳞的长度(length of posterior section of nasal,LpSN)(精度0.01 mm)。借助体式显微镜[舜宇ESDPTOP SZN71,舜宇光学科技(集团)有限公司]计数鳞片,若是对称性鳞片,则对左右两侧分别计数。计数形态特征包括背鳞(dorsal scale rows,DSR)、上颌齿(maxillary teeth counts,MT)、鼻鳞(nasals,NS)、颊鳞(loreals,LR)、上唇鳞(supralabials,SPL)、下唇鳞(infralabials,IFL)、颞鳞(temporals,TMP)、前颞鳞(anterior temporals,aTMP)、中颞鳞(middle temporals,mTMP)、后颞鳞(posterior temporals,pTMP)、腹鳞(ventral scales,V)、颔片(chin-shields,CS)、眶上鳞(supraocular,SpO)、眶前鳞(preoculars,PrO)、眶后鳞(postoculars,PtO)、颏鳞(mentals,PM)和尾下鳞(subcaudals,SC)。
1.3 分子鉴定
使用TIANGEN 试剂盒(天根生化科技有限公司,北京)提取肝组织基因组总DNA,线粒体COⅠ基因扩增引物Chmf4/Chmr4(Cheet al.,2012)。用Lasergene 7.1(DNASTAR)的SeqMan 对所获得的序列进行拼接。从GenBank 下载已发表的脊蛇属物种的COⅠ序列,选取老挝拟须唇蛇Parafimbrios laoTeynié,David,Lottier,Le,Vidal & Nguyen,2015(Teyniéet al.,2015)、越南拟须唇蛇P. vietnamensisZiegler,Ngo,Pham,Nguyen,Le & Nguyen,2018(Ziegleret al.,2018)、克 氏须唇 蛇Fimbrios klossiSmith,1921(Schizaset al.,2015)和爪哇闪皮蛇Xenodermus javanicusReinhardt,1836(Rovatsoset al.,2016)为外群(附表1)。利用MEGA 7.0 的Clustal W 算法对获得的所有序列进行比对(Kumaret al.,2016),人工校正。利用PartitionFinder 2.1.1选择数据集最适模型(Lanfearet al.,2017),最终在MrBayes 3.2.4(Ronquistet al.,2012)和Raxml-GUI 1.3(Silvestro & Michalak,2012)中分别构建贝叶斯树和最大似然树。

表1 基于COⅠ基因的不同产地云开脊蛇个体间的遗传距离Table 1 Genetic distance between snakes from different regions based on COⅠ gene
附表1 样本序列信息Appendix 1 Information for COI gene sequences used in this study
利用MEGA 7.0(Kumaret al.,2016)中的未校正距离模型估算物种间的遗传距离。
2 结果
2.1 形态鉴定与描述
采自画稿溪国家级自然保护区的1 号雌性亚成体(图1)和1 号雄性幼体,其特征表现为眶前鳞和眶后鳞常缺失、尾下鳞不成对等特征,初步鉴定为脊蛇属物种。其具体形态如下:(1)通身背鳞23 行,除最外行外均起棱明显且具有自前向后增强趋,最外行背鳞显著扩大;(2)上颌齿数24枚,靠近吻端略小,后端几乎等大且排列不均匀;(3)鼻间鳞2枚,前额鳞2枚;(4)额鳞1枚,未入眶;(5)颌片2对,与腹鳞相切,其间无小鳞,后颌片略长于前颌片;(6)顶鳞1对,与正前方额鳞相切;(7)鼻鳞二分,前大后小;(8)颊鳞1 枚,扁长,长约2 倍于高;(9)颞鳞3列,2+2+3式,第一列全入眶;(10)上唇鳞6 对,第4~5 枚入眶;(11)下唇鳞5~6 对,第1 对在颏鳞末端相切,左右前3 枚与前颔片相切;(12)肛鳞完整;(13)背面呈棕黑色,略具金属光泽,腹面乳白色;(14)脊线不明显,从顶鳞后方发出向后延伸至尾端(图2)。其中,雌性个体TaL/TL 百分比为24.2%,LSBP>LSBI(17.7 mm vs. 11.6 mm),下唇鳞6 枚,腹鳞145 枚(加2 枚前腹鳞),尾下鳞65 枚;雄 性 幼 体TaL/TL 为24.5%,LSBP>LSBI(9.7 mm vs. 8.6 mm),下 唇 鳞5 枚,腹 鳞146 枚(加2 枚前腹鳞),尾下鳞64 枚。形态上,该2 号标本与云开脊蛇鉴别特征较为一致(附表2)。经形态鉴定,此次采集的蛇类标本为云开脊蛇。

图1 四川画稿溪国家自然保护区采集的云开脊蛇(标本号:YBU 22050;李科 摄)Fig. 1 Achalinus yunkaiensis from the Huagaoxi National Nature Reserve,Sichuan(voucher No.:YBU 22050;photoed by Li Ke)

图2 四川画稿溪云开脊蛇(标本号:YBU 22050)的形态特征(李科 摄)Fig. 2 Morphological characteristics of Achalinus yunkaiensis (voucher No.:YBU 22050)from the Huagaoxi National Nature Reserve,Sichuan (photoed by Li Ke)

附表2 不同产地的云开脊蛇特征比较Appendix 2 Comparison of the characteristics of Achalinus yunkaiensis from different regions
2.2 系统发育关系
由于雄性幼体较小,为保证标本的完整性未采集分子样,只采用1 号雌性样本构建系统发育树。经MEGA 7.0 比对剪切后获得654 bp 的COⅠ基因片段(GenBank登录号:OR344062)。基于PartitionFinder v2.1.1 对数据集进行密码子分区选取的最适模型分别为K80+I+G(第1 密码子),TIM+I(第2 密码子)和TIM+G(第3 密码子);构建的贝叶斯树和最大似然树的拓扑结构基本一致(图3)。结果显示所有脊蛇属物种聚为一个高支持率的分支(PP=1.00,BS=100),明确了脊蛇属内部分物种间关系;此次采集的标本与云开脊蛇聚为一支,该支系与黑脊蛇、阿里山脊蛇A. nigerMaki,1931 和台湾脊蛇A. formosanusBoulenger,1908 又形成单系。四川个体与云开脊蛇其他个体间遗传距离为3.1%~3.3%(表1),该遗传分化普遍小于脊蛇属属内物种间遗传距离(≥3.4%;Yanget al.,2022)。
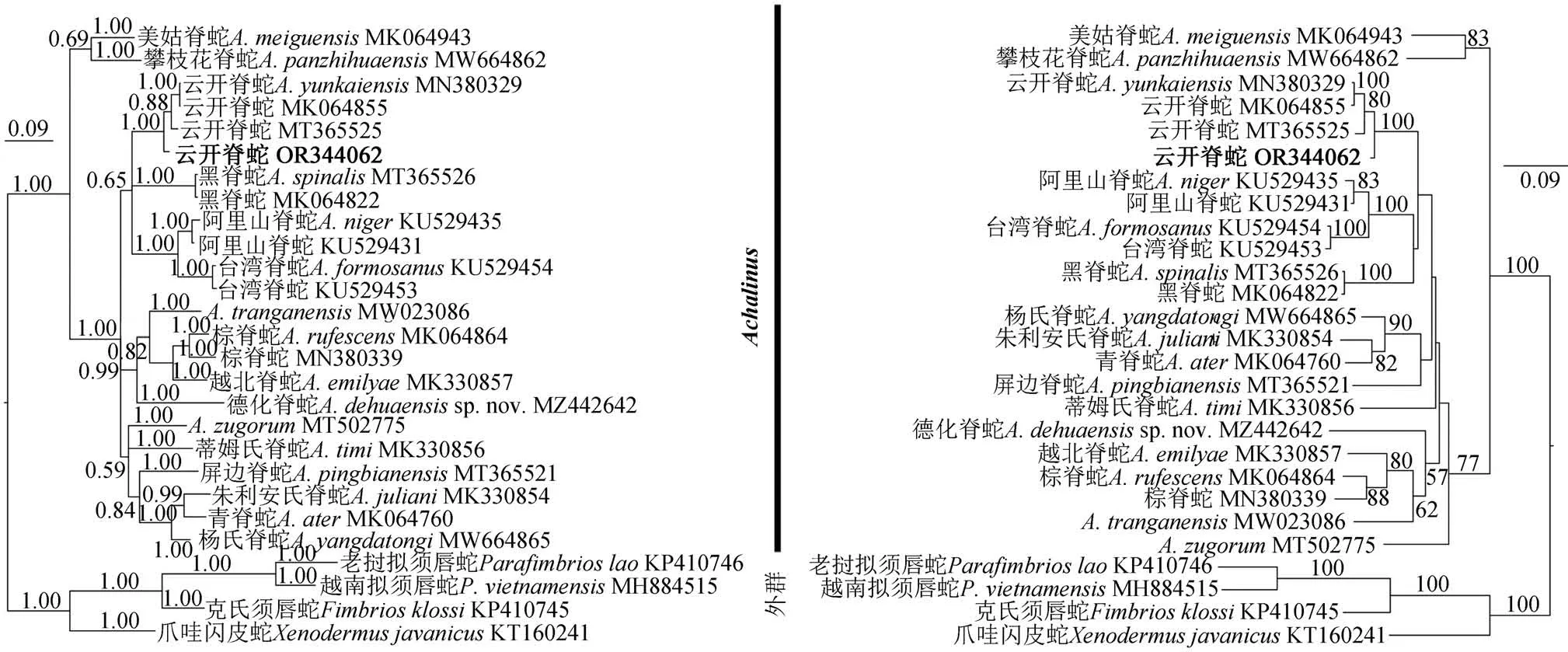
图3 基于线粒体COⅠ基因构建的脊蛇属系统发育树(左:贝叶斯树;右:最大似然树)Fig. 3 Phylogenetic tree of Achalinus based on mitochondrial COⅠ gene(left:Bayesian phylogenetic tree;right:Maximum Likelihood tree)
结合形态学比较和系统发育分析,确定此次采集的蛇类标本YBU 22050 和YBU 22148 为云开脊蛇,系四川省蛇类新分布记录种。
3 讨论
云开脊蛇首先发现于我国广东省云开山大雾岭地区(Wanget al.,2019),喻敏等(2020)报道了该物种在广西猫儿山的新分布记录。广东的样本采集于4月、6月和8月,广西的样本采集于8月,四川样本则采集于6 月。与四川相比,广东纬度更低,4 月气温可能就已达到适合该蛇活动的温度。并且此次发现的2 条云开脊蛇均采集于四川泸州画稿溪国家级自然保护区内的一处竹林与公路相接的浅草丛中(图4),该采集地位于四川东南部,地接云贵高原,为典型的丹霞地貌景观,植被类型以常绿阔叶林为主,与以前采集地植被类型相同、生境相似,同时具有丰富的其他同域物种分布。

图4 四川画稿溪国家级自然保护区云开脊蛇栖息生境(王朝英 摄)Fig. 4 Habitat of Achalinus yunkaiensis in the Huagaoxi National Nature Reserve,Sichuan(photoed by Wang Chaoying)
在形态上,四川的雌性个体在体全长(306 mm vs. 339 mm)、尾长(74 mm vs. 53 mm)、腹鳞(145 枚vs. 153枚)和尾下鳞(65枚 vs. 51枚)与广西雌性个体存在一定差异;同时,四川雄性个体TaL/TL 大于广东雄性个体(24.5% vs. 18.5%~20.0%),尾下鳞也多于广东个体(64 枚 vs. 49~56 枚),但腹鳞少于广东个体(146枚 vs. 151~162枚),且广东个体的前额鳞沟与鼻间鳞沟相等,而四川个体和广西个体的前额鳞沟均长于鼻间鳞沟,这说明云开脊蛇存在一定地理变异,这是否源于遗传差异或标本量不足仍有待进一步增加标本量核实。
云开脊蛇最大种下遗传距离达到3.3%,该结果与Yang等(2022)认为的脊蛇属最小种间遗传距离(3.4%)较为接近。脊蛇属物种属于古老的中小型穴居蛇类,活动能力十分受限,斑块化的生境极易导致各地理种群间产生隔离,从而产生遗传和外部形态上的分化,当前,有关脊蛇属的研究仍然十分缺乏,各物种的有效性还应结合更多的种群及样本进行评估。本研究扩展了云开脊蛇在我国的地理分布,丰富了四川蛇类的物种多样性,同时揭示云开脊蛇可能存在一定的地理变异,这为脊蛇属的种间关系研究提供了一定参考。
致谢:感谢云南大学硕士研究生唐奎在标本采集中提供的帮助,感谢宜宾学院动物多样性与生态保育宜宾市重点实验室本科生李莉、蒋玉莲在分子实验中的帮助。
