circROBO1 上调KLF5 促进视网膜细胞瘤侵袭的机制研究
2023-09-23刘越峰张志伟罗卫民
刘越峰,张志伟,李 欣,罗卫民
(1.十堰市太和医院眼科(湖北医药学院附属医院),湖北 十堰 442000;2.南华大学衡阳医学院肿瘤研究所,湖南 衡阳421001;3.湖北医药学院,湖北 十堰 442000)
视网膜母细胞瘤(retinoblastoma,RB)为婴幼儿时期的一种常见恶性眼内肿瘤,好发于三岁以内的儿童,临床特征主要表现为眼红、眼痛、视力低下或白瞳症等,严重者威胁患儿的视功能及生命[1]。目前,RB 的早期诊断仍较为困难,大部分患儿被确诊时已处于疾病的晚期阶段,故探究RB 的发生、发展机制,寻找调控该过程的关键分子,可能是提高RB早期诊断、降低复发转移的关键所在[2,3]。
环状RNA(circRNAs)为一种共价闭合的单链非编码RNA 分子,在多种恶性肿瘤(如乳腺癌、肺癌、RB 等)中的表达水平存在显著异常[4-6]。circRNAs 与多种癌症的恶性进展密切相关,有潜力成为各种恶性肿瘤早期诊断或临床治疗的靶标,如circAGFG1 可增加乳腺癌细胞的增殖与迁移能力,是三阴乳腺癌的潜在治疗靶点[7]。研究显示circROBO1 可通过调控KLF5 促进乳腺癌转移[8]。本团队前期研究发现微小RNA miR-222 可通过靶向调控RB1 而促进RB 的生长和侵袭能力[9]。同时我们也发现circROBO1 可通过靶向调控miR-217 而促进RB 细胞的增殖,circROBO1 对RB 生长具有促进作用,但对其侵袭迁移能力尚未进行深入研究。因此,本研究拟探究circROBO1 对RB 侵袭的影响及其调控机制,为RB 治疗寻找潜在治疗靶点。
1 材料与方法
1.1 主要材料
人视网膜母细胞瘤细胞Y79(中科院上海生化细胞所),Lipo 3000、蛋白裂解液及抑制剂、抗体KLF5 及β-actin(北京博尔,中国),DMEM 培养基、血清和抗生素(依赛科,中国),circROBO1 干扰小RNA 及其对照、RNase R 酶(吉赛生物,中国)、细胞核/胞浆细胞组分提取试剂盒(BioVision,美国)、TaqMan 逆转录及荧光定量PCR 试剂盒(Takara,美国),智能计数仪(上海萌薇,中国),Transwell 小室和基质胶(0.8 nm,康宁,美国),Nanodrop 2000 紫外可 见 分 光 光 度 计(Thermo,美 国),OLYMPUS BX51 正置显微镜(奥林巴斯,日本)。
1.2 细胞培养及分组
根据供应商提供的说明书将Y79 细胞培养于含胎牛血清(FBS)、青霉素和链霉素(浓度分别为10%、102U/mL 及0.1 g/L)的DMEM 培养基内。实验分si-control 组、空白组和si-circROBO1 组。转染 circROBO1 干 扰 小 RNA(5'-GAAAACACAAGAUAUGAAA-3')序 列 的 为si-circROBO1组,转染其对照无关序列的为si-control 组(5'-UUCUCCGAACGUGUCACGU-3'),未转染任何序列的为空白组。
1.3 细胞转染
收集Y79 细胞,在含10% FBS DMEM 培养基中稀释成104细胞/mL 细胞悬液,并将其接种于6 孔板中,其中每孔加入2 mL 的Y79 细胞悬液,37 ℃,含5% CO2的细胞培养箱中孵育。当Y79 细胞的汇合度达80%左右时准备转染,采用无菌EP 管混匀lipofectamin 3000 和 转 染 试 剂(si-circROBO1 和si-control),20 min 后将上述混合物加入到无血清培养基(不含抗生素)中,并加入到待转染6 孔板中,37 ℃,含5 % CO2中培养7 h,10% FBS DMEM 培养48 h 后的细胞可用于后续实验。
1.4 RNA 提 取 和qRT-PCR 检 测
根据试剂商提供的RNA 抽提试剂盒提取Y79细胞的总RNA,并于紫外可见分光光度计上检测,将符合实验需求的RN 置于-20 ℃冰箱内保存备用。以Y79 细胞总RNA 为模板,根据试剂商提供的逆转录试剂盒及qSYBR-Green-containing PCR试剂盒说明书进行操作,进行相对定量分析(Bio-Rad IQTM5 Multicolor 实时荧光定量系统,美国)。circROBO1引物及其对照引物用于qRT-PCR 实验,其 中U6为 内 参,相 对 表 达 量 根 据2-ΔΔCT计 算,ΔΔCT=(CTcircRNA-CTU6)target-(CTcircRNA-CTU6)control。反应总体系20 μL,且每组实验设置3 个复孔。
1.5 划痕实验
使用marker 笔于24 孔板背后,用直尺比着在正中央划一道线,横穿过孔(所有器械均已灭菌或紫外消毒)。取1 mL 5×105对数生长期的细胞悬液加入到24 孔板中,使细胞贴壁培养一夜,次日用200 μL 枪头比着直尺垂直于背后横线划痕,PBS 清洗细胞3 次,去除划下细胞。加入无血清培养基。放入37 ℃ 5%CO2培 养 箱,培 养。24 h 后 取 样,镜 下拍照。
1.6 Transwell 实验
将转染24 h 后的Y79 配置成3×105/mL 的单细胞悬液(转染方法见1.3)。Transwell 上层上腔层内铺加基质胶,并静置6 h 待其成膜(Transwell 侵袭实验需要完成此步骤,Transwell 迁移实验无需此步)。将150 μL 单细胞悬液接种到小室上腔层,将350 μL含20%胎牛血清的培养基添加至下腔层,37 ℃,含5% CO2培养箱内孵育24 h 后进行结晶紫染色。除去上室细胞清洗后晾干,镜下观察计数。
1.7 RNase R 酶消化实验
提取Y79 细胞总RNA 后,根据是否加入Rnase R,将RNA 分为两组:Rnase R 组(加Rnase R 消化)和对照组(不加Rnase R 消化)。随后在PCR 仪中,经37 ℃反应30 min 后再经70 ℃反应10 min 将酶灭活后逆转录,使用根据circROBO1 环状接口处设计的引物及根据线性亲本基因ROBO1 设计的引物,进行qRT-PCR 检测,比较线性亲本基因ROBO1与circROBO1在Y79 细胞中的表达水平改变,明确circROBO1在Y79 细胞中的结构稳定性。
1.8 circROBO1 亚细胞定位实验
根据试剂商提供的细胞核/胞浆细胞组分提取试剂盒的说明书进行操作,分别提取Y79 细胞的细胞质及细胞核RNA,分别采用qRT-PCR 检测circROBO1、及细胞质对照和细胞核对照18 S 分别再细胞质及细胞核中相对表达量,根据其在两者中的比例作统计图。
1.9 双荧光素酶报告基因实验
将104个Y79 细胞接种到每个6 孔板中,放入37 ℃5%CO2培养箱,培养24 h。按照Lipo 3000 说明书操作分别共转染circROBO1野生型、突变型,或KLF5mRNA 野 生 型、突 变 型,与miR-885-5p或miR-NC,转染后按照试剂盒说明书进行后续操作[13],分 别验证circROBO1和KLF5与miR-885-5p之间的结合关系。
1.10 蛋白质印迹分析
实验分为si-circROBO1 组(同1.2)、si-control组(同1.2)和空白组。使用RIPA 裂解液、PMSF 从Y79 细胞中分离细胞总蛋白,用后续的SDS-PAGE凝胶分离。蛋白质转膜条件为300 mA 2 h。最后将PVDF 膜 转 移 至4 °C 冰 箱 内,使 用KLF5 及β-actin抗体处理过夜(浓度为1∶1 000),次日于室温下使用特异性二抗孵育,显影后,Image J 用于结果分析。
1.11 数据分析
使用SPSS 20.0 处理数据,结果以“均数±标准差”记录,t检验用于组间分析。P<0.05 为差异具有统计学意义。
2 结果
2.1 circROBO1 在视网膜Y79 细胞中的结构稳定性鉴定
RNase R 酶消化后的qRT-PCR 实验结果表明,与对照组相比,环状RNAcircROBO1的相对表达量在RNase R 酶处理后并无显著变化;而线性ROBO1的相对表达量在经RNase R 酶消化后,则显著降低(t=16.18,P<0.05,图1)。

图1 RNase R 酶消化处理后circROBO1 及线性ROBO1 的相对表达量Fig 1 Relative expression levels of circROBO1 and linear ROBO1 after RNase R enzyme digestion treatment
2.2 circROBO1 在视网膜Y79 细胞中的定位
分别提取Y79 细胞质及核的RNA,分析circROBO1在胞质及胞核中的定位,qPCR 结果显示,根据circROBO1在胞质与胞核之比,其在细胞质中的含量高于细胞核中(t=13.04,P<0.05,图2),即circROBO1的亚细胞定位主要为细胞质(图2),而β-actin主要位于细胞质,18S 则主要位于细胞核。
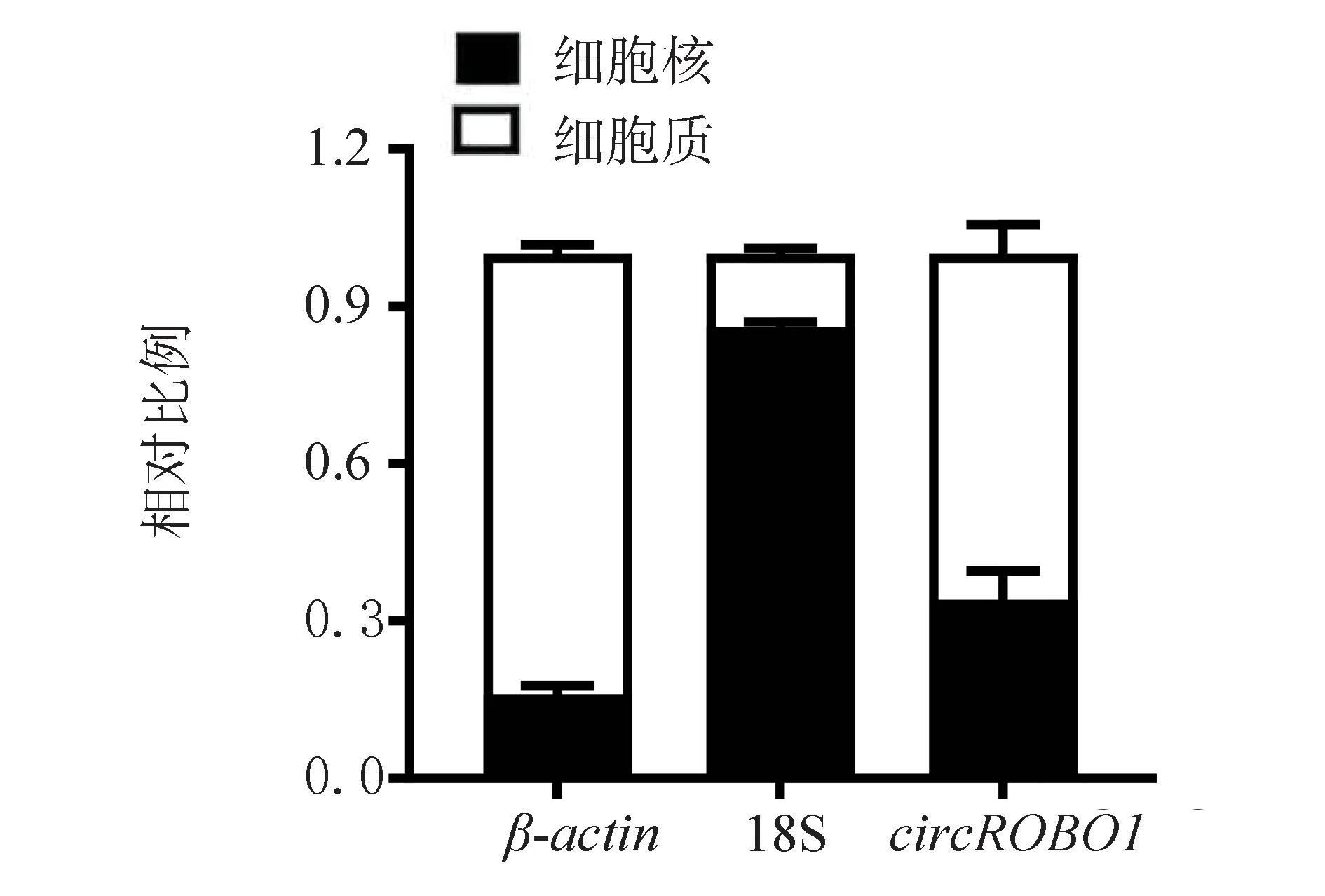
图2 circROBO1 在视网膜Y79 细胞中的亚细胞定位Fig 2 Subcellular localization of circROBO1 in retinal Y79 cells
2.3 circROBO1对视网膜Y79细胞迁移能力的影响
划痕实验结果显示:与si-control 组比较,si-circROBO1 组视网膜Y79 细胞的迁移率显著降低(t=22.54,P<0.05,图3A、B),即外源性沉默circROBO1 后,视网膜Y79 细胞的迁移能力显著降低。
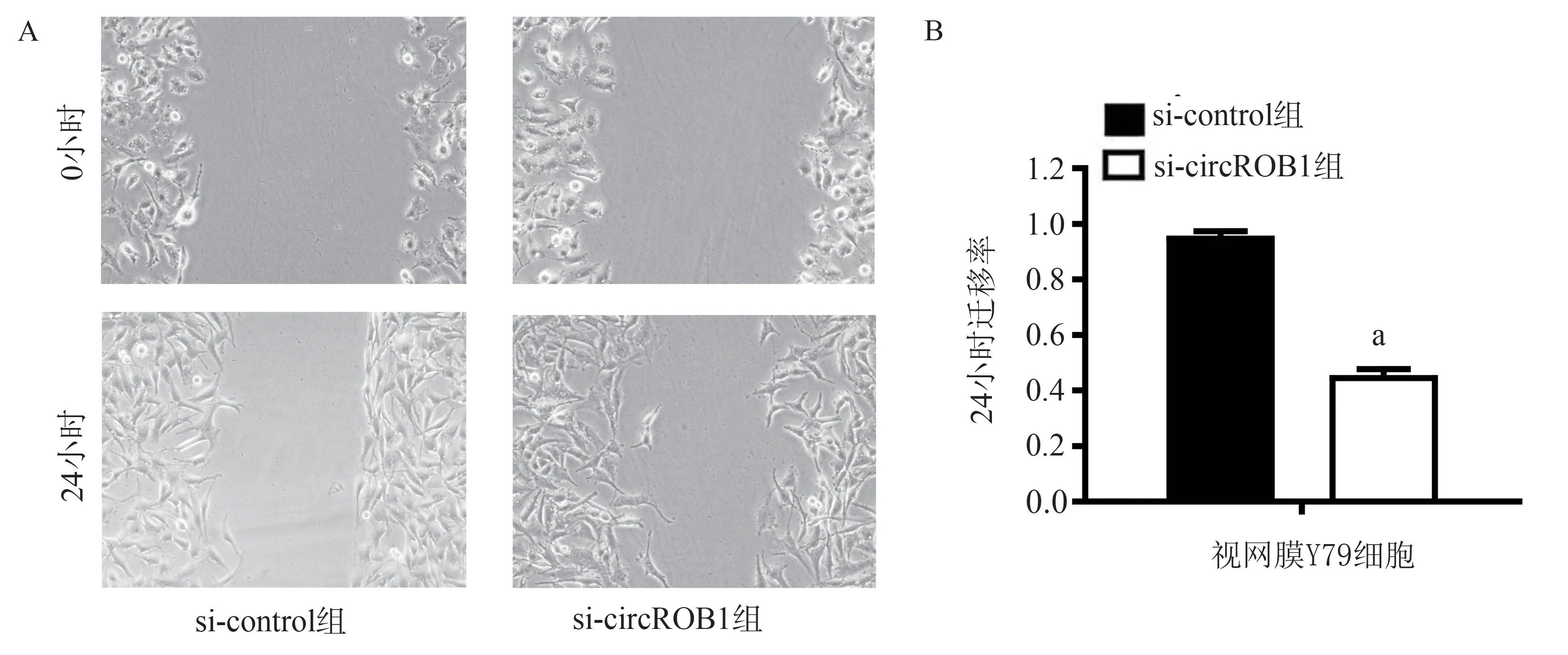
图3 沉默circROBO1 对视网膜Y79 细胞迁移能力的影响Fig 3 Effect of circROBO1 silencing on retinal Y79 cell migration
2.4 circROBO1对视网膜Y79细胞侵袭能力的影响
Transwell 结果显示:与si-control 组比较,si-circROBO1 组视网膜Y79 细胞的侵袭与迁移细胞 数 均 显 著 降 低(t侵袭=11.65,t迁移=12.06,P<0.05,图4A、B)。即外源性沉默circROBO1,视网膜Y79 细胞的侵袭能力显著降低。
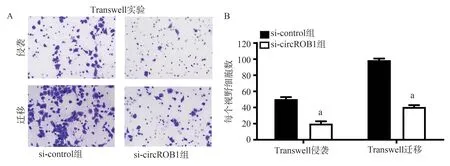
图4 沉默circROBO1 对视网膜Y79 细胞Transwell 侵袭和迁移能力的影响Fig 4 The effect of silencing circROBO1 on the Transwell invasion and migration ability of retinal Y79 cells
2.5 circROBO1 与miR-885-5p 的结合位点预测及验证
根据生物信息学软件CircInteractome 的预测,结果显示circROBO1序列中可能存在1 个miR-885-5p结合位点。双荧光素酶报告基因实验结果显示:和野生型circROBO1共转染时,与miR-NC 相比,miR-885-5p 共转染组荧光素酶活性显著降低 (t=11.39,P<0.05,图5B);和突变型circROBO1共转染时,与miR-NC 相比,miR-885-5p 共转 染 组 活 性 无 显 著 差 异 (t=1.112,P>0.05,图5B),即circROBO1能够与miR-885-5p 相互作用。
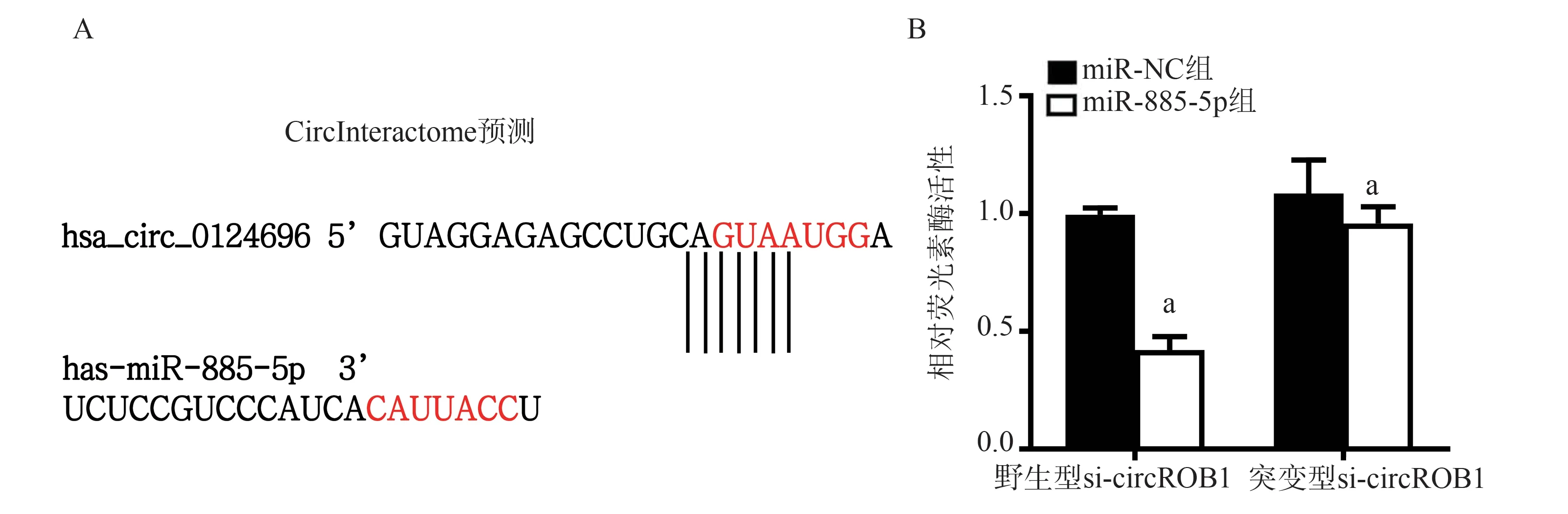
图5 circROBO1 与miR-885-5p 的结合位点预测及验证Fig 5 Prediction and validation of binding sites between circROBO1 and miR-885-5p
2.6 miR-885-5p 下游靶基因的预测及验证
根据TargetScan 软件的预测,结果显示miR-885-5p序列中可能存在1 个与KLF5mRNA 结合的位点(图6A)。双荧光素酶报告基因实验结果显示:与对照miR-NC+KLF5 野生型共转染组比较,miR-885-5p+KLF5 野生型共转染组的细胞荧光素酶活性显著降低(t=17.26,P<0.05,图6B);与对照miR-NC+KLF5 突变型共转染组比较,miR-885-5p+KLF5 突变型共转染组活性无显著差异 (t=1.684,P>0.05,图6B),即miR-885-5p能够与KLF5相互作用。

图6 miR-885-5p 与KLF5 的结合位点预测及验证Fig 6 Prediction and validation of binding sites between miR-885-5p and KLF5

图7 circROBO1 通过KLF5 促进视网膜Y79 细胞瘤侵袭Fig 7 CircROBO1 promotes invasion of retinal Y79 cell tumor through KLF5
2.7 circROBO1 通 过KLF5 促 进 视 网 膜Y79 细 胞瘤侵袭
siRNA 沉默视网膜Y79 细胞中circROBO1 表达后的蛋白免疫印迹实验,结果显示,si-circROBO1转染组KLF5 的蛋白表达量显著降低(图 7A、B)。即外源性沉默circROBO1 可显著降低KLF5 蛋白表达。
3 讨论
circRNA 为近年来新发现的非编码RNA 亚类,是通过具有共价闭环结构的反向剪接机制所产生的,不仅可充当 微小RNA (microRNA,miRNA) 和蛋白质的吸附海绵,还可以调节基因表达、进行表观遗传修饰,转化为肽段,并产生假基因[10,11]。circRNA 的表达失调可导致多种人类疾病的发生及进展,包括恶性肿瘤及眼部疾病等[12]。近年来越来越多的研究发现circRNA 在各类眼科疾病的发病机制中发挥重要作用,表明circRNA 有成为眼科疾病诊断、进展评估及预后判断生物标志物的潜力。
已有研究显示,circ_0000527 在RB 组织和细胞中高表达,circ_0000527 沉默可抑制RB 细胞的增殖、迁移和侵袭,并促进细胞凋亡,且circ_0000527敲低可抑制RB 细胞体内肿瘤生长,该过程主要是通 过circ_0000527 海 绵 化miR-98-5p,miR-98-5p 靶向XIAP 而介导RB 细胞恶性进展而实现的[13]。据报道,circMKLN1 与RB 患者较好的预后密切相关,作为环状RNA,circMKLN1 在体外对RNase R 消化具有抗性,主要富集在细胞质中发挥作用,且circ-MKLN1 过表达可抑制了RB 细胞的增殖、迁移和侵袭能力,circMKLN1 过表达可阻碍体内肿瘤的生长,该过程是通过circMKLN1 调控miR-425-5p及其靶基因PDCD4而实现的[14]。上述研究表明circRNAs 的确可通过海绵吸附miRNAs 而促进或抑制RB 的恶性进展。
本研究中通过RNase R 酶消化后的qRT-PCR实验,发现环状RNAcircROBO1在RB 细胞Y79 中结构稳定;该结果与已有研究证实的“环状RNA 因其环状结构使其结构非常稳定且对RNA 核酸外切酶具有抗性”这一结果相一致[15]。细胞核及细胞质中circROBO1 的富集分析发现其主要定位于细胞质,而沉默circROBO1 显著抑制RB 细胞的迁移及侵袭能力。进一步的蛋白免疫印迹结果表明,沉默circROBO1 显著抑制KLF5 的蛋白表达。而在乳腺癌中已经证实胞质circROBO1 可通过miR-217-5p/KLF5 轴促进肿瘤肝转移[8]。上述结果与“作为广泛分布在细胞质中的circRNA,其可作为miRNA 海绵来调控下游关键基因的表达”相一致[16]。
Krüppel 样因子(Kruppel-like factor 5,KLFs)为KLF 家族转录因子中的成员,可参与多种生物学过程及不同类型的疾病,尤其是癌症。KLF5 可调节多种靶基因的表达,如Cyclin D1、Nanog和Slug等[17-20]。研究显示KLF5 可参与多种生物学功能,如细胞干性、增殖 、细胞凋亡、自噬和迁移等[21-24]。并且KLF5 可促进多种恶性肿瘤的转移与进展,如BAP1 拮抗WWP1 介导的转录因子 KLF5 的泛素化并抑制自噬,从而促进黑色素瘤的恶性进展[25],KLF5 去泛素化可促进乳腺癌细胞的增殖与转移[26],长 链 非 编 码RNA LncRNA PVT1 可 通 过KLF5/β-catenin 信号通路而调节三阴性乳腺癌的恶性进展[27]。作为一种转录因子,KLF5 在乳腺癌和结直肠癌中过表达,是其预后不良的潜在生物标志物[28-30]。研究显示,通过生物信息学预测及双荧光素酶报告基因,发现circROBO1可吸附下游的miR-217-5p,KLF5作为miR-217-5p的直接靶基因,miR-217-5p可直接调控KLF5的表达,由circROBO1/KLF5/FUS 轴所建立的正反馈循环可促进乳腺癌的肝转移[8]。而本研究中我们发现在视网膜Y79 细胞中,circROBO1的沉默可显著抑制KLF5的蛋白表达,经过预测及荧光素酶报告基因实验验证我们发现circROBO1与miR-885-5p之间、miR-885-5与KLF5之间均存在结合位点,而该研究结果与乳腺癌中的结果较为相似,表明RB 中circROBO1也是通过靶向KLF5而促进视网膜Y79 细胞瘤侵袭的,该过程是通过circROBO1与miR-885-5p相互作用实现的。目前该机制的探究还不太完善,后续我们将纳入更多RB 细胞系进行验证,并进行RIP 等实验继续验证。
本研究初步证实circROBO1 可通过上调KLF5促进视网膜Y79 细胞瘤侵袭,该结果可为临床RB靶向药物的开发提供新的思路。
作者贡献度说明:
刘越峰:分子生物学实验及数据处理;张志伟:数据处理及作图;李欣:行政支持;罗卫民:研究思路及文章审核。
作者声明不存在利益冲突关系。
