MCC950 通过抑制NLRP3 炎症小体通路引发的焦亡缓解原发性痛经*
2023-09-21汪少华潘思安袁菡钰岳增辉
汪少华 潘思安 薛 晓 袁菡钰 李 娟 岳增辉 刘 余
(湖南中医药大学针灸推拿与康复学院,长沙 410208)
原发性痛经 (primary dysmenorrhoea, PDM) 被定义为没有盆腔器质性病变的月经期疼痛性疾病,通常以痉挛性耻骨上疼痛为特征[1,2]。PDM 影响50%~90%年轻女性的生活,包括人际关系、学习和工作,而且引发焦虑[3,4]。目前治疗PDM 的一线药物为非甾体抗炎药 (non steroidal anti-inflammatory drugs, NSAIDs),存在着恶心、呕吐、食欲减退甚至消化道溃疡等不良反应[5,6]。目前,没有足够的证据说明单独使用NSAIDs 是最有效且安全的[7]。因此,寻找安全有效治疗或缓解PDM的药物是有必要的。
核因子-κB (nuclear factor-κB, NF-κB) 蛋白复合物家族在B 细胞成熟过程中被诱导,并作为二聚体发挥作用[8]。NF-κB 家族包括五种蛋白质二聚体,存在于典型的p50/p65 型中[9]。p50/p65 二聚体可在淋巴细胞和巨噬细胞中触发NF-κB 诱导的转录作用,从而在炎症的病理生理过程中发挥作用[10,11]。在对外源性信号的反应中,IκB 的翻译后修饰导致IκB 磷酸化,导致NF-κB 二聚体进入细胞核并激活靶基因,从而进一步加剧炎症[12]。总之,NF-κB 是介导炎症反应的关键因子。
含核苷酸结合寡聚化结构域核苷酸结合寡聚化结构域样受体蛋白3 (nucleotide-binding oligomerization domain-like receptor protein3, NLRP3) 炎症小体是由NLRP3、半胱氨酰天冬氨酸特异性蛋白酶-1 (cysteinyl aspartate specific proteinase-1, caspase-1)和含有半胱天冬酶募集结构域的凋亡相关斑点样蛋白 (apoptosis-associated specklike protein containing a caspase recruitment domain, ASC) 组装而成的细胞质多蛋白复合体[13,14]。PDM 中出现NLRP3 蛋白寡聚化,NLRP3 选择性抑制剂MCC950 通过NF-κB/COX-2/PG 通路抑制PDM 大鼠子宫组织NLRP3 炎症小体激活并减轻炎症[15]。NLRP3 的寡聚化驱动caspase-1 依赖性的蛋白裂解以及IL-1β 和IL-18 的成熟和释放[16,17]。激活的caspase-1 将gasdermin D(GSDMD) 裂 解 成GSDMD-N 末 端 (GSDMD-N),GSDMD-N 进入质膜,导致细胞膜破裂,最后细胞焦亡[18]。因此,NLRP3 炎症小体作为炎症反应的核心,参与了PDM 的发生发展,可能是治疗或缓解PDM 的新靶点。
PDM 中发生了NLRP3 炎性小体的激活,但PDM大鼠子宫组织损伤的机制尚不完全清楚。因此,为进一步研究PDM 大鼠子宫损伤的机制,本研究使用苯甲酸雌二醇和催产素构建PDM 大鼠模型,探讨MCC950 治疗或缓解PDM 的机制。
方 法
1.实验动物和分组
健康未孕雌性SPF 级Sprague-Dawleg (SD)大鼠30 只,6~8 周龄,体质量180~220 g,购自北京华阜康生物科技有限公司,饲养在湖南中医药大学实验动物中心,实验动物许可证号:SCXK(京)2019-0008。大鼠被饲养在20℃~25℃的适宜温度下,自由饮水和进食。1 周后采用随机数表法将30 只大鼠随机分为3 组:对照组、模型组(PDM组)和MCC950 组,每组10 只。所有动物实验均经湖南中医药大学实验动物伦理委员会审查批准(伦理批号LL2021040703),符合中国国家标准《实验动物-动物福利伦理审查指南》(GB/T 35892-2018)。
2.材料和方法
(1)主要药品和仪器
苯甲酸雌二醇 [生产批号:(2020) 11025,宁波第二激素厂,中国];催产素(货号:P1029,批次:07;分子式:C45H70N12O14S2;分子量:1067.2;纯度:98.52%)和MCC950(货号:S7809,批次:07;分子式:C20H23N2O5S.Na;分子量:426.46;纯度:99.40%)均购自美国Selleck;大鼠PGE2 ELISA 试剂盒(生产批号:20211111-J2944)和大鼠PGF2α ELISA 试剂盒(生产批号:20211111-J3106)均购自湖南艾方生物科技有限公司;NF-κB p65 抗体(货号:GB12142),phospho-NF-κB p65 抗体(货号:GB11142-1),HRP-山羊抗鼠二抗(货号:GB-23301)和HRP-山羊抗兔二抗(货号:GB23303)均购自武汉赛维尔生物科技有限公司;NLRP3 抗体(货号:ab263899),GSDMD 抗体(货号:ab-219800)和N-terminal GSDMD 抗体(货号:ab215-203)均购自英国abcam;caspase-1 抗体(货号:#2225),cleaved caspase-1 抗体(货号:#89332)和IL-1β 抗体(货号:#12242)均购自美国Cell Signaling Technology;ASC 抗体(货号:bs-6741R)购自美国Bioss Antibodies;IL-18 抗体(货号:10663-1-AP)购自美国Proteintech;β-actin(货号:AF5001)购自上海碧云天生物技术有限公司。
酶标仪(型号:RT-6100,Rayto,美国),光学显微镜(型号:BA410E,Motic,中国),切片机(型号:UC7,Leica,德国),高速冷冻离心机(型号:Sorvall ST 16R,Thermo Fisher,美国),电泳槽(Bio-rad,美国),化学发光仪(型号:6300,CLINX,中国),灰度分析软件(alphaEaseFC,Alpha Innotech,美国)。
(2)PDM 大鼠模型的建立
采用苯甲酸雌二醇和催产素联合给药建立大鼠PDM 模型。雌激素通过调节细胞增殖、血管生成和增加催产素受体 (oxytocin receptor, OTR) mRNA表达来提高子宫敏感性[19,20]。催产素能有效诱导子宫收缩,引起子宫相对缺血和缺氧,导致疼痛[21]。对照组大鼠皮下注射0.9%氯化钠注射液,连续注射10 天,第11 天腹腔注射0.9%氯化钠注射液。另外两组大鼠每日早上9 时皮下注射苯甲酸雌二醇(剂量:第1 天2 mg/kg,第2~9 天0.8 mg/kg,第10 天2 mg/kg)。苯甲酸雌二醇的剂量和时间参考其体内药代动力学研究[22]。第11 天上午9 时,PDM 组和MCC950 组每只大鼠腹腔注射2 U 催产素(浓度5 mg/ml,体积0.4 ml)。MCC950 组第11 天在注射苯甲酸雌二醇30 min 后腹腔注射10 mg/kg 的MCC950(浓度5 mg/ml)。
(3)大鼠扭体反应的测定
大鼠扭体反应参考Wei 等[23]的报道。记录大鼠在注射催产素后30 min 内的扭体次数和扭体潜伏期。扭体潜伏期是从注射催产素到第1次扭体的时间。发生扭体反应后,使用2%的戊巴比妥钠麻醉大鼠,腹主动脉采血,切除子宫组织。最后,对老鼠实施安乐死。用无菌手术剪刀将约5 mm×5 mm×5 mm大小的子宫组织置于4%多聚甲醛溶液中固定。其余的组织被放置在液氮中进行后续检查。扭体评分标准:正常运动、探索行为以及躯干和后肢伸展记0 分;腹部凹陷、身体倾斜记1 分;后肢伸展、后爪背屈、骨盆旋转的身体伸展记2 分;腹肌收缩、后肢向后伸展、身体有僵直的扭转评分记3 分。
(4)显微镜检查
采用随机数表法每组选取5 只大鼠,从每只大鼠的同侧采集同一节段的子宫组织进行显微镜观察。将大鼠子宫组织在4%多聚甲醛中室温固定24 h,脱水,二甲苯透明,石蜡包埋,以厚度5 μm切片。组织随后用二甲苯和乙醇脱蜡,干燥并用苏木精-伊红染色。用中性树胶封片后,光学显微镜观察大鼠子宫的组织形态结构。大鼠子宫病理损伤评分参考Wei 等[24]的实验。根据显微镜下子宫的形态特征,无病理的子宫记为0 分;子宫内膜变性坏死记为1 分;固有层水肿记为2 分;固有层腺体增加记为3;炎性细胞浸润固有层评分记为4 分;子宫肌层炎症评分记为5 分。
(5)酶联免疫吸附试验(ELISA)检测大鼠血清PG 变化
取腹主动脉的血液在室温下自然凝固10~20 min,离心20 min 左右 (2000~3000 r/min),仔细收集血液上清液。根据ELISA 试剂盒的说明进行血清中PGE2 和PGF2α 的测定。
(6)Western blot 检测
采用随机数表法每组选取5 只大鼠,取大鼠同侧同段子宫组织约30 mg,用匀浆器低温研磨。加入预冷的RIPA 裂解缓冲液、蛋白酶抑制剂和磷酸化酶抑制剂,置于冰上30 min。4℃,12,000 r/min离心10 min,除去含有蛋白的上清,加入样品缓冲液,在沸水浴中加热10 min,使蛋白完全变性。采用BCA 蛋白测定试剂盒测定蛋白质标准曲线,计算蛋白质浓度。经电泳、PVDF 膜转移和阻断后,用Tris 缓冲盐水Tween-20 (TBST) 稀释特异性一抗。膜与以下主要抗体在4℃孵育过夜:NF-κB p65 (1:2000),phospho-NF-κB p65 (1:3000),NLRP3(1:1000),caspase-1 (1:3000),cleaved caspase-1(1:3000),ASC (1:3000),IL-1β (1:3000),IL-18(1:3000),β-actin (1:2000),GSDMD (1:1000) 和GSDMD-N (1:1000)。然后,用TBST 冲洗膜,并与稀释的二抗孵育60 min,ECL 法曝光。
3.统计学分析
所有实验数据均采用 IBM SPSS Statistics 26.0统计软件进行分析。所有数据的统计描述均采用均数±标准差(±SD)表示。如果数据满足正态性和方差齐性,则使用单因素方差分析;若不满足正态性,使用Kruskal-Wallis H 检验。显著性水平α =0.05,P< 0.05 为差异具有统计学意义。
结 果
1.MCC950 可减少PDM 大鼠的扭体次数,延长扭体潜伏期
在注射催产素30 min 内,对照组没有扭体反应。与对照组相比,PDM 组大鼠的扭体次数(P< 0.001,见图1A)和评分(P< 0.001,见图1B)明显增加,说明PDM 大鼠模型构建成功。与PDM 组相比,MCC950 组的扭体次数(P< 0.05,见图1A)和评分(P< 0.05,见图1B)显著降低。由于无扭体反应,对照组无扭体潜伏期。与PDM 组相比,MCC950组扭体潜伏期明显延长(P< 0.05,见图1C)。

图1 各组大鼠注射催产素30 min 后扭体次数、扭体评分及扭体潜伏期比较(n = 10,±SD)Fig.1 Comparison of writhing times, writhing scores and writhing incubation periods 30 min after oxytocin injection in each group (n = 10,±SD)
2.MCC950 可减轻PDM 大鼠子宫内膜细胞损伤,降低子宫病理损伤评分
对大鼠子宫进行HE 染色,观察MCC950 对大鼠子宫内膜细胞损伤的保护作用。光学显微镜下,对照组子宫未见明显组织学改变。子宫内膜上皮细胞排列整齐,柱状,生理性变性少,未见明显炎症细胞浸润。PDM 组子宫内膜上皮细胞大量空泡化坏死,多螺旋小动脉充血,可见中性粒细胞浸润。MCC950 组子宫上皮出现少量空泡变性坏死,无充血,有少量中性粒细胞浸润(见图2A-C)。病理评分以显微镜评价为基础,对符合标准的评分进行累积评价。与对照组相比,PDM 组子宫组织病理学损伤评分显著升高(P< 0.01,见图2D)。与PDM 组相比,MCC950 组病理评分明显降低(P< 0.05,见图2D)。综上所述,MCC950 对PDM 大鼠子宫损伤有保护作用。
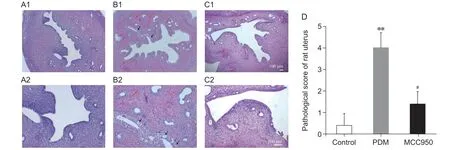
图2 各组大鼠子宫组织组织病理学变化及病理损伤评分比较(n = 5,±SD)Fig.2 Histopathological changes and comparison of pathological damage scores in the uterine tissue of rats in each group (n = 5,±SD)
3.MCC950 下调PDM 大鼠血清中PGF2α 含量及PGF2α/PGE2 比值
前列腺素(prostaglandin, PG)是PDM 的发病机制之一。此外一项人体试验表明,PGF2α 抑制剂对治疗PDM 有效[25]。因此,PG 可以用来评价PDM动物模型的成功与否。与对照组相比,PDM 组大鼠血清中PGF2α(P< 0.01,见图3A)及PGF2α/PGE2 比值(P< 0.001,见图3B)显著升高。与PDM 组比较,MCC950 组大鼠血清中PGF2α(P<0.05,见图3A)及PGF2α/PGE2 比值(P< 0.001,见图3B)明显降低。
4.MCC950 抑制PDM 大鼠子宫组织NF-κB 的激活
采用Western blot 法测定大鼠子宫组织中NF-κB的含量。与对照组相比,PDM 大鼠子宫组织中NF-κB p65 和 磷 酸 化NF-κB p65 (phospho-p65) 蛋 白 的 表达水平显著升高(P< 0.001,见图4)。此外,与PDM 组相比,MCC950 组大鼠子宫组织NF-κB p65(P< 0.01)和磷酸化NF-κB p65(P< 0.001,见图4)水平明显降低。
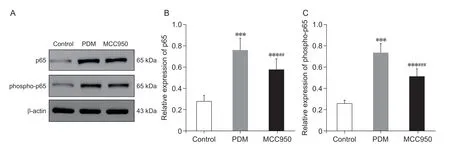
图4 各组大鼠子宫组织NF-κB p65 及磷酸化NF-κB p65 蛋白表达水平比较(n = 5,±SD)Fig.4 Comparison of protein expression levels of NF-κB p65 and phosphorylated NF-κB p65 in the uterine tissue of rats in each group (n = 5,±SD)
5.MCC950 抑制NLRP3 炎症小体蛋白的表达和下游cleaved caspase-1 的释放
为了证实MCC950 对炎症小体及其下游caspase的作用,通过Western blot 分析了炎症小体蛋白和cleaved caspase-1(见图5A)。与对照组相比,PDM组大鼠子宫组织NLRP3(P< 0.001,见图5B)、ASC(P< 0.001,见图5C)、caspase-1(P< 0.01,见图5D)、cleaved caspase-1(P< 0.01,见图5E)蛋白表达水平显著升高。与PDM 组比较,MCC950组NLRP3(P< 0.001,见图5B)、ASC(P< 0.01,见图5C)、caspase-1(P< 0.05,见图5D)、cleaved caspase-1(P< 0.05,见图5E)蛋白表达水平显著降低。
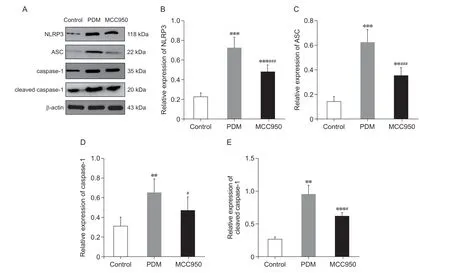
图5 各组大鼠子宫组织NLRP3 炎症小体组件NLRP3、ASC、caspase-1、cleaved caspase-1 蛋白表达水平的比较(n = 5,±SD)Fig.5 Comparison of the protein expression levels of the NLRP3 inflammasome components NLRP3, ASC, caspase-1 and cleaved caspase-1 in the uterine tissue of rats in each group (n = 5,±SD)
6.MCC950 下调焦亡蛋白和炎症因子的表达
为了确定PDM 大鼠子宫组织损伤和焦亡的原因,通过Western blot 检测GSDMD、GSDMD-N、IL-1β 和IL-18 的蛋白表达水平(见图6A)。与对照组相比,PDM 组大鼠子宫组织中GSDMD(P<0.001,见图6B)、IL-1β(P< 0.001,见图6C)、GSDMD-N(P< 0.01,见图6D)、IL-18(P< 0.001,见图6E)蛋白表达水平明显升高。与PDM 组比较,MCC950 组大鼠子宫组织中GSDMD(P< 0.001,见图6B)、IL-1β(P< 0.001,见图6C)、GSDMD-N(P< 0.05,见图6D)、IL-18(P< 0.001,见图6E)蛋白表达水平显著降低。
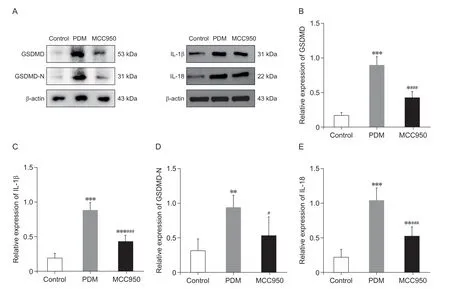
图6 各组大鼠子宫组织中焦亡蛋白及炎症因子的比较(n = 5,±SD)Fig.6 Comparison of pyroptotic proteins and inflammatory factors in the uterine tissue of rats in each group (n = 5,±SD)
讨 论
NLRP3 炎症小体活化而引起的炎症是许多疾病的基础[26]。NLRP3 炎症小体介导炎症、组织损伤和妇科疾病如子宫内膜异位症、妇科肿瘤和多囊卵巢综合征等[27]。NLRP3 炎症小体的组装是由模式识别受体对危险相关分子模式和病原体相关分子模式做出反应决定的[13]。本研究结果显示,PDM大鼠子宫组织中NLRP3 炎症小体组件蛋白表达上升,NF-κB 和磷酸化NF-κB 的表达也上升。NLRP3炎症小体的激活包括两个过程:一是上调NLRP3 的炎症小体成分(包括NLRP3、ASC 和caspase-1),并对NLRP3 进行翻译后修饰,获得激活条件;二是对NLRP3 激活子的识别和NLRP3 炎症小体的形成[28]。在PDM 中,NLRP3 炎症小体激活可能由toll 样受体等模式识别受体引起,导致NF-κB 激活和基因转录,或由线粒体功能障碍引起[29~31]。因 此PDM 中 出 现NLRP3 炎 症 小 体 和NF-κB 通路的活化。本研究结果显示PDM 大鼠子宫组织GSDMD 和GSDMD-N 显著升高。NLRP3 炎症小体激活后,caspase-1 被剪切形成具有活性的cleaved caspase-1,基因敲除实验证实了这一点[32]。cleaved caspase-1 剪切GSDMD 形成GSDMD-N,GSDMD-N在细胞膜上形成孔,引起焦亡[18]。此外,ASC 可以增加GSDMD 的含量,促进GSDMD 活化[33]。相反,靶向敲低GSDMD 基因阻断了细胞焦亡,而在增加外源性GSDMD 表达后,焦亡被重新激活[34]。这些实验表明GSDMD 是焦亡的执行蛋白。因此可以推断,在PDM 大鼠子宫组织中,NLRP3 炎症小体发生活化并引发了焦亡。此外,本研究结果显示,在PDM 大鼠子宫组织中,IL-1β 和IL-18 蛋白表达升高。PDM 模型大鼠血清中PGF2α 以及PGF2α/PGE2 表达明显升高。Lim 等的研究表明[35],分娩时人的子宫收缩,NLRP3 蛋白表达增加,PGF2α 表达也增加。靶向敲除NLRP3 后,PGF2α 表达明显降低。PG 中特别是可引起子宫收缩的PGF2α,参与了PDM 的发病[36]。随着PGF2α/PGE2 比值升高,导致子宫平滑肌收缩加重[37],从而加重PDM。这些结果提示,NLRP3 激活引起的PG 升高是PDM 的发病机制之一。细胞死亡命名委员会将细胞焦亡定义为依赖caspase-1 导致染色质碎裂、质膜透化的细胞肿胀和炎性内容物IL-1β 和IL-18 释放的程序性细胞死亡[38]。通常,细胞焦亡会引起组织损伤,炎症物质IL-1β 和IL-18 本身就是致痛源[39~41]。因此,PDM 的组织损伤和疼痛可能是由焦亡过程中释放的IL-1β 和IL-18 介导的。
MCC950 是一种选择性NLRP3 抑制剂,对全身多个器官具有药理作用[42]。MCC950 作用于NLRP3 蛋 白NACHT 结 构 域 的Walker B 位 点,从 而 阻 断ATP 水 解 和NLRP3 活 化[43]。NLRP3是一种包含氨基端pyrin 结构域、羧基端富含亮氨酸重复结构域和中央NACHT 结构域的三联蛋白。刺激诱导NLRP3 寡聚化通过NLRP3 蛋白中央NACHT 结构域的同型相互作用招募ASC,ASC 中的caspase 招募结构域招募caspase-1[28]。在生理条件下,NLRP3中LRR-LRR与PYD的相互作用保持稳定,避免过早激活[44]。最近的报道显示[45],MCC950 与天然NLRP3 蛋白形成稳定的十聚体,提示MCC950可以有效结合NLRP3 并发挥抑制作用。本研究结果 显 示,MCC950 干 预 后NF-κB 通 路 和NLRP3炎症小体组件蛋白的表达均下降。NLRP3 蛋白是NLRP3 炎症小体的传感器[46]。抑制NLRP3 蛋白后,ASC 和caspase-1 的招募能力下降,NLRP3 炎症小体受到抑制,不能有效产生cleaved caspase-1,进而抑制IL-1β 的分泌。MCC950 能有效抑制NLRP3 蛋白激活,但MCC950 干预如何抑制NF-κB 的转录还不完全清楚。有证据表明,NLRP3 下游因子IL-1β 可通过磷酸化位点Ser316、Ser529 和Ser536 激活NF-κB信号转导[47,48],这可能与MCC950 干预抑制NF-κB有关。由于NLRP3 炎性小体和cleaved caspase-1 被抑制,导致下游GSDMD 的数量和活性降低,因此MCC950 干预抑制PDM 大鼠的子宫焦亡和炎症物质的释放。本研究结果表明,MCC950 抑制了PGF2α的表达。由于NLRP3 被直接抑制,PGF2α 的产生也减少。与对照组相比,MCC950 组行为学和部分Western blot 蛋白表达差异有统计学意义,子宫病理损伤评分、血清PG 和部分Western blot 蛋白表达差异无统计学意义,说明经过MCC950 的干预后,PDM 大鼠得到缓解。
本研究存在潜在的局限性:首先,没有测量基因水平上的表达,没有分离子宫细胞以获得更精准的药物控制。此外,PDM 中NLRP3 炎症小体激活的启动机制目前尚不清楚。因此,未来可以从以下方面开展研究:①从遗传学角度出发,不排除基因编辑;②从子宫或其他组织中分离细胞并施加药物干预,以获得更精准的靶点;③确定PDM 中NF-κB通路和NLRP3 炎症小体激活的机制。
综上所述,MCC950 可抑制PDM 中NF-κB 通路和NLRP3炎症小体的活化,减少焦亡和组织损伤,从而缓解PDM。
利益冲突声明:作者声明本文无利益冲突。
