草莓炭疽病研究进展
2023-09-08张舒媛
王 丰,张舒媛,卢 洁,马 跃
(沈阳农业大学a.植物保护学院,b.园艺学院,沈阳 110161)
草莓是蔷薇科的一种多年生草本植物。根据2018 年的统计数据,中国的草莓栽培面积达到173 333 hm2,成为主要的草莓生产国和出口国[1]。虽然草莓的栽培面积在不断扩大,但是病害仍然是制约草莓生产的主要因素。在草莓栽培过程中,冠腐病、根腐病、白粉病、炭疽病等真菌性病害造成草莓产量和品质的下降。其中,由炭疽菌属Colletotrichum引起的草莓炭疽病在田间的流行和蔓延更为严重。目前的研究表明,Colletotrichum主要有3 个种:C.fragariae、C.acutatum和C.gloeosporioides[2]。Colletotrichumspp.可以侵染草莓的所有器官,造成果实腐烂、叶片斑点、茎冠腐烂和根部腐烂等[3]。此外,该真菌以菌丝体和拟菌核的形式在土壤中越冬。翌年,菌丝体产生分生孢子,可以直接侵染草莓,这种病害的越冬形式不利于田间控制草莓炭疽病的发生,对草莓的产量和品质构成严重威胁[4]。本研究结合国内外研究现状,综合阐述了草莓炭疽病的致病菌的分类、病害发生情况以及防治方法。
1 炭疽菌概述
疾病控制的先决条件是明确病原。炭疽菌属是全球十大重要的真菌病原体之一,寄主范围广泛,在苹果[5]、梨[6]、桃[7]、草莓[8]等植物上均有记载。随着自然环境以及农业耕作方式的改变,炭疽菌属已经进化出许多不同的种类[9]。
1.1 炭疽菌分类
从时间的角度来看,草莓炭疽病致病菌的进化经历了一个漫长的过程。据报道,1882年,PENZIP等[10]在意大利柑橘中分离得到C.gloeosporioides,能够导致柑橘炭疽病的发生。之后,在其他物种中均发现该病原菌能够造成炭疽病的发生。1931 年,BROOKS 等[11]在美国东南部首次发现草莓感染炭疽菌(C.fragariae)。基于形态学观察,很难区分C.fragariae和C.gloeosporioides,因此在大多数情况下,草莓炭疽病在早期阶段认为是由C.fragariae或C.gloeosporioides侵染引起的。1965 年,SIMMONDS 等[12]在澳大利亚确定C.acutatum为草莓果实腐烂的病原体,通过形态学观察将C.acutatum从C.gloeosporioides中单独分类出来。除了C.acutatum与C.gloeosporides复合体的物种之外,1973年美国报告C.dematium对草莓有致病性(图1)。在1983年,HOWARD 等[13]认为C.gloeosporioides与C.fragariae在草莓上的致病性是一样的,所以将C.fragariae归类到C.gloeosporioides中。
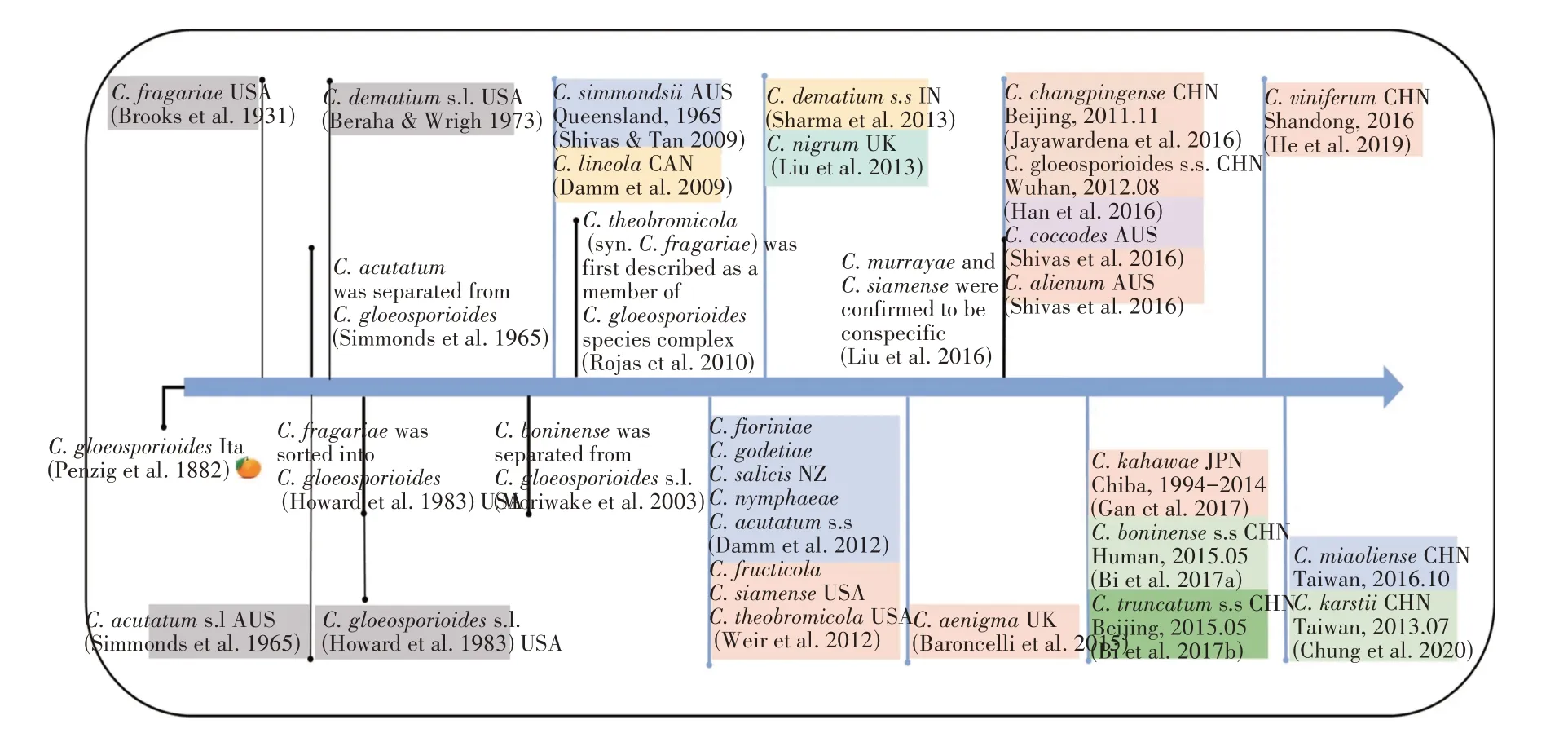
图1 草莓炭疽病致病菌种发现时间轴线图[25]Figure 1 Timeline of strawberry anthracnose pathogenic species discovery
直到20世纪末,分子生物学鉴定技术被广泛使用。MORIWAKI等[14](2003)通过分子鉴定将C.boninense从C.gloeosporioides中分离出来。SHIVAS等[15]在2009年将C.simmondii重新评估为C.acutatums.l的成员。此外,DAMM 等[16]在2009 年将以C.truncatum命名的菌株被重新确定为C.lineola。在2010 年,ROJAS 等[17]将C.theobromicola确定为C.gloeosporioides的一个成员。同年,C.dematium被认为是一个具有弯曲分生孢子的物种复合体,该复合体中的C.lineola在加拿大从患病的草莓中首次分离出来。次年,印度再次报告C.dematiums.s.对草莓有侵染性[18]。至此,C.dematium复合体中的两个种(C.lineola、C.dematiums.s.)被报告为草莓致病菌。
2012 年,在系统发育分析的基础上,人们对Colletotrichunspp.的遗传特性的了解更加深入,分类方式和标准也在不断完善。DAMM 等[19]通过ITS、GAPDH、ACT、CHS、HIS3和TUB2的多位点系统发育分析明确的区分了C.acutatum复合体中的7 个种(C.acutatums.s.、C.fioriniae、C.godetiae、C.miaoliense、C.nymphaeae、C.salicis和C.simmondsii)。随后,C.gloeosporioides复合体有9 个种(C.aenigma、C.alienum、C.changpingense、C.fructicola、C. gloeosporioidess.s.、C. kahawae、C. siamense、C. viniferum和C. theobromicola)被报告为草莓炭疽病的致病菌。2015 年,BARONCELLI 等[20],报道C.gloeosporioides复合体的C.aenigma在英国感染草莓植株。2016 年,从患病的草莓叶片、叶柄和匍匐茎中分离出致病真菌为C.gloeosporioidess.s.,C.murrayae也被发现为该病的致病菌。同一年内,LIU 等[21]证明C.murrayae与C.siamense是同种。JAYAWARDENA 等将从中国北部患病草莓根茎中分离得到的致病真菌C.changpingense确定为C.gloeosporioides复合体中的一个新种,同时发现C.alienum也可致病。2017 年,GAN 等[22]首次在日本患病草莓中分离得到C.kahawae致病菌。2019 年,HE 等[23]在中国山东省首次发现C.gloeosporioides复合体的另一个种C.viniferum能够侵染草莓叶片。CHUNG 等[24]在2020 年报道C.boninense复合体中的C.karstii能侵染草莓叶片,而C.boninense复合体中的C.boninenses.s.被报道在中国湖南省娄底市可引起草莓果实腐烂,故C.boninense复合体中的两个种(C.boninenses.s.和C.karstii)被确定为致病菌。在同一年内C.truncatum复合体中的C.truncatums.s.在北京昌平区的患病草莓中首次被分离出来,至此C.truncatums.s.以及2个单体C.nigrum和C.coccodes被确定为草莓炭疽病的致病菌。
截止目前,共有23 个炭疽菌的致病种被发现可引起草莓炭疽病,即C.gloeosporioides复合体的9 个种(C.aenigma、C. alienum、C. changpingense、C. fructicola、C. gloeosporioidess.s.、C. kahawae、C. siamense、C. viniferum和C.theobromicola),C.acutatum复合体的7 个种(C.acutatums.s.、C.fioriniae、C.godetiae、C.miaoliense、C.nymphaeae、C.salicis和C.simmondsii),C.boninense复合体中的2 个种(C.boninenses.s.和C.karstii),C.dematium复合体(C.lineola和C.dematiums.s.),C.truncatum复合体的1 个(C.truncatums.s.)以及2 个单体C.nigrum和C.coccodes(图2)。毫无疑问,引起草莓炭疽病的致病种种类将在未来继续增加。
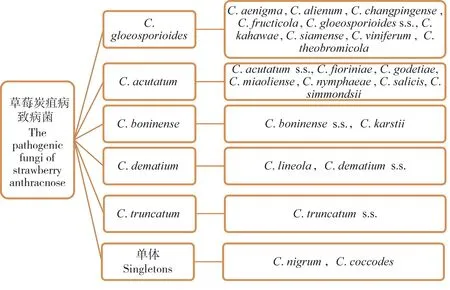
图2 草莓炭疽病致病菌分类Figure 2 Classification of the pathogenic fungi of strawberry anthracnose
1.2 炭疽菌形态特征以及生物学特性分析
随着草莓炭疽菌种类的不断增加,不同炭疽菌的形态特征和生物学特性各不相同,形态特征主要表现在菌落颜色、分生孢子形态等方面(图3)[26-27],生物学特性主要表现为温度、光照、酸碱度、碳源和氮源对菌丝生长速度和产孢量的影响。
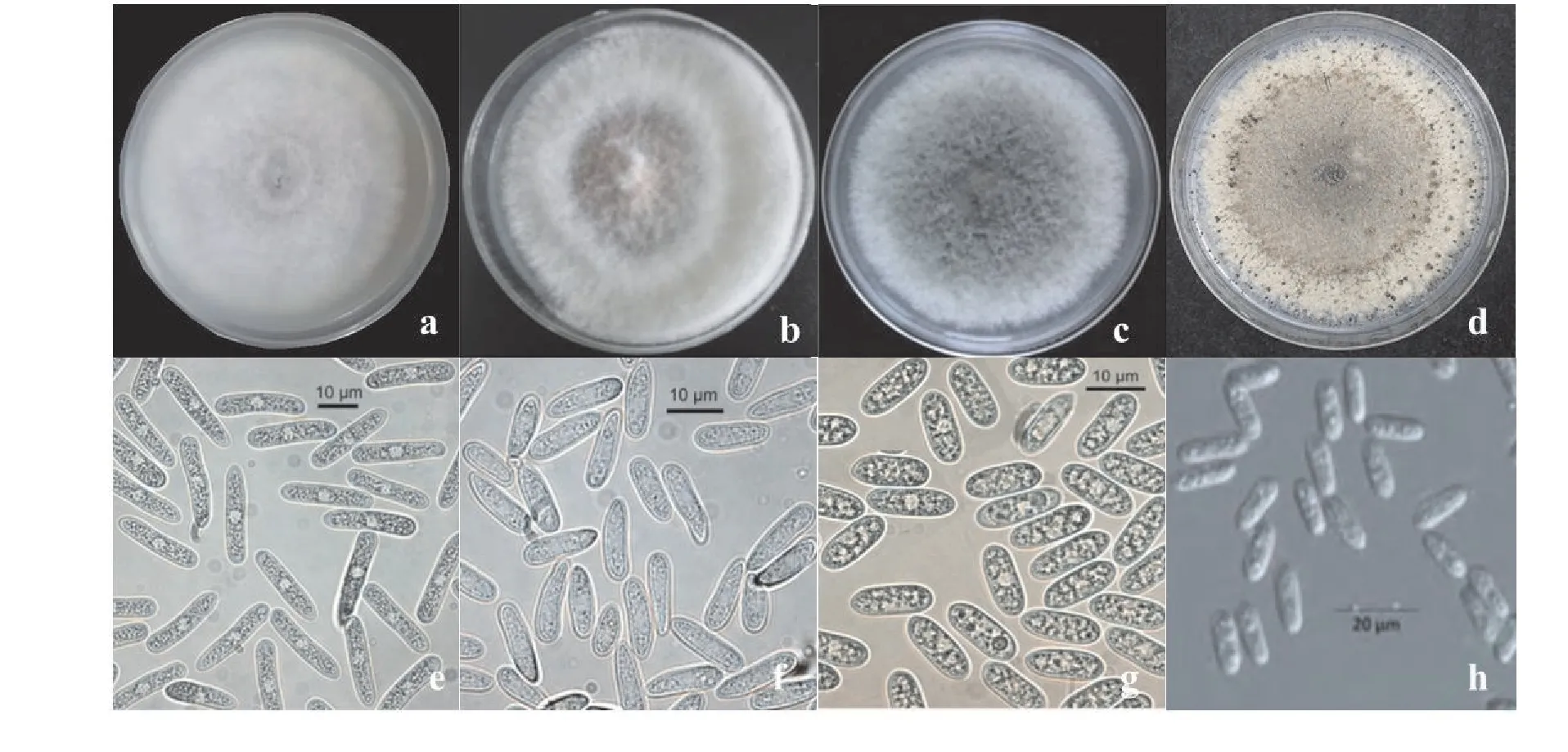
图3 草莓中分离出的Colletotrichum spp.的菌落以及孢子形态[32-33]Figure 3 Morphology of mycelium and conidia of Colletotrichum spp. isolated from strawberry
草莓炭疽菌(C.fragariae)菌落呈现地毯状,气生菌丝,菌落颜色初期为米色,逐渐变为深灰色。分生孢子为倒卵形,分生孢子堆为粉色或者橙红色,无菌核,附着孢为棒状[28]。
胶孢炭疽菌(C.gloeosporioides)菌落白色,菌丝生长密集,菌落颜色初期为白色,后期变为深灰色。刚毛有或者无,分生孢子团为浅粉色,分生孢子为柱形,两端为钝圆形,无色。研究表明,胶孢炭疽菌在PDA 和PSA 培养基中均可正常生长,培养基中使用果糖作为碳源,蛋白胨作为氮源最适宜胶孢炭疽菌生长。PSA培养基有助于胶孢炭疽菌产生孢子堆,菌丝和孢子最适宜的生长温度为25 ℃,最适pH值为6,适当光照能促进产生孢子[29]。
暹罗炭疽菌(C.siamense)菌落颜色在培养初期为白色,后期变为灰色,菌丝絮状较为稀疏,菌落背面中央为黑褐色,边缘颜色较淡。分生孢子团为橙红色,分生孢子为柱状。研究表明,暹罗炭疽菌在28 ℃下菌丝生长速度最快,能够适应各种碳源和氮源。同时,使用纤维素和蛋白胨作为碳源和氮源,pH为10,持续光照的条件下最适宜产生孢子[30]。
尖孢炭疽菌(C.acutatum)菌落颜色在4~5 d 内呈现白色,后期变成灰褐色。分生孢子为梭形,分生孢子堆为橙色或者粉红色,分生孢子为椭圆形或者梭形,有稀疏刚毛,附着孢具有色素且形态大小不一致[31]。研究表明,尖孢炭疽菌在26 ℃时菌丝生长速度最快,在30 ℃时产孢量最多,同时,适当光照有利于产生孢子堆。培养基中添加可溶性淀粉、甘露醇和葡萄糖能够促进菌丝生长和孢子产生[32-34]。
1.3 炭疽菌危害草莓症状
草莓炭疽病是制约草莓生产的关键因素之一,严重影响草莓的产量和品质[35]。在田间栽培过程中,该真菌病害蔓延十分迅速,并且病原菌对环境的适应性很强。研究表明,炭疽菌能够在整个草莓生长过程中引起病害,尤其在高温高湿的条件下病情快速发展。草莓在夏季主要处于幼苗期,抗性相对弱,而夏季高温高湿的环境条件导致草莓炭疽病的流行较为严重[36]。草莓炭疽病主要发生在幼苗期和定植初期,不同炭疽病致病菌的感染症状也存在差异。草莓炭疽菌(C.fragariae)可以侵染草莓果实,在收获后的储存期间严重限制了水果的保质期[37](图4)。草莓炭疽菌和胶孢炭疽菌(C.gloeosporioides)均可以侵染草莓的匍匐茎以及叶柄造成黑色病斑,并且草莓炭疽菌在叶柄上的危害症状比胶孢炭疽菌严重[38-39]。胶孢炭疽菌还可以侵染草莓叶片和根部,产生毒性物质进而破坏草莓根部木质部导管,最终使草莓萎蔫死亡[26,40-41](图5)。然而,暹罗炭疽菌(C.siamense)作为胶孢炭疽菌的复合种之一,对草莓叶片和根部也有致病力[42]。研究表明,在中国东部草莓栽培产区,暹罗炭疽菌是造成草莓根腐病的病原菌之一[43-45](图6)。尖孢炭疽菌(C.acutatum)主要造成果实黑斑,也可以侵染根和叶片[46](图7)。

图4 草莓炭疽菌危害草莓症状[26,37]Figure 4 Symptoms of C. fragariae infected strawberry

图5 胶孢炭疽菌危害草莓症状[40-41]Figure 5 Symptoms of C. gloeosporioides infected strawberry

图7 尖孢炭疽菌危害草莓症状[46]Figure 7 Symptoms of C. acutatum infected strawberry
1.4 致病因子研究
植物受到病原真菌侵染是多种因子调控的结果,主要有细胞壁降解酶、黑色素、毒素、信号传递途径、活性氧含量和微量元素等。明确上述致病因子在侵染过程中的作用是研究病原菌侵染机制的基础。炭疽菌通过活体营养或者死体营养的方式侵染植物,在死体营养过程中主要由细胞壁降解酶降解植物组织,表现为寄主细胞中炭疽菌生长产生大量次生菌丝并分泌细胞壁降解酶,为之后的生长获得营养物质[31]。
植物细胞壁是植物的天然保护屏障,能够抵制病原菌的入侵。植物细胞壁主要由果胶、纤维素和半纤维素构成,而降解植物细胞壁需要特殊的酶类参与。当致病菌接触植物表面时,病原菌能够产生识别并降解细胞壁结构的细胞壁降解酶,比如果胶酶、纤维素降解酶、蛋白酶和多酚氧化酶类等。其中,炭疽菌产生的果胶裂解酶和多聚半乳糖醛酸酶是降解植物组织的关键酶类。在炭疽菌和植物相互作用的过程中,炭疽菌还能够产生果胶酶,而果胶酶是病原菌入侵植物组织造成植物病斑和腐烂的胞外酶。研究表明,当胶孢炭疽菌在植物体内大量繁殖时,该致病菌的胶酸盐裂解酶基因pel、pelB以及果胶裂解酶基因pnlA的表达量升高,导致植物细胞壁受到损害,植株最终死亡[47]。除此之外,由于致病菌的刺激,植株自身可能会出现过敏性坏死反应。
植物病原菌还可以产生许多特异性和非特异性的毒素,参与宿主识别过程、寄主植物生理生化过程、影响植物组织结构进而使细胞结构变异[26]。C.capsici病原菌分泌的毒素和培养滤液能够抑制水稻种子根部生长、辣椒种子萌发以及果实腐烂。康子腾等[48]研究发现,炭疽菌的附着孢内含有黑色素,能够参与病原菌的侵染过程,是关键的致病因子。与黑色素合成相关基因PKS1、SCD1和THR1缺失的炭疽菌菌株丧失了入侵植物的能力。当C.trifolii侵染植物早期,均可以检测到THR1和SCD1基因的表达。关于炭疽菌的致病机制的研究目前主要的关注点集中在细胞壁降解酶和钙信号通路上,而关于毒素和黑色素的研究相对较少。炭疽菌的致病过程是比较复杂的,涉及多种基因的调控,对其致病因子以及致病机制还有待研究。
2 病害流行规律
炭疽病病原菌以菌丝或者拟菌核的形式依附在果实和枯枝烂叶中越冬,在土壤中也可以安全存活[9]。来年产生分生孢子堆,当环境条件适宜时,分生孢子萌发并且凭借雨水、风或者昆虫广泛传播,随机落在寄主植物表面。萌发的孢子可以通过自然孔口直接感染叶片,也可以通过伤口入侵,造成叶片、果实或者根系发生病变。炭疽菌引发草莓植株发生病变首先需要分生孢子依附于植株表面在条件适宜时萌发并产生芽管,芽管顶端膨大形成附着孢,随后附着孢分泌黏液将芽管固定在植物表面,最终从附着孢上产生侵染丝穿透植物表皮细胞获取营养,破坏植物细胞结构,导致植物产生病变最终死亡[49]。病变部位在感染后期还会产生分生孢子堆作为新的侵染源头,加快炭疽病的蔓延和流行。
3 草莓炭疽病的发生与分布
从地理角度来看,炭疽病的流行已经影响了全世界的草莓栽培产区(图8)。一般来说,对草莓致病的C.acutatum复合菌属在欧洲、非洲和大洋洲流行[50-51],而C.gloeosporioides复合菌种在东亚和北美的草莓田中普遍存在[52-53]。其中,C.acutatum复合体中的C.nymphaeae和C.acutatums.s.是侵染草莓的全球性病原体,在六大洲均有报道。在非洲和欧洲,C.nymphaeae在草莓田里比其他物种更盛行。在亚太地区东部的中国北部、韩国和日本,C.fructicola很普遍。2019-2022 年,C.siamense在美国[54]和中国[55]越来越盛行。中国作为草莓生产大国,C.siamense(226个分离物,占总数的45.93%)是中国草莓栽培地中最主要的病原菌,几乎分布在所有省份,其次是C. fructicola(189 个分离物,38.41%),同样分布在整个中国(图9)。而C. acutatums.l.、C. boninenses.l.和C.truncatums.l.物种在中国基本上是随机出现的。值得注意的是,在湖北省和台湾省C.siamense的致病性占据主导地位,C.fructicola在四川省普遍存在,在上海、山东和安徽省,C.fructicola和C.siamense在草莓田里的发生率相当高。然而C.nymphaeae和C.miaoliense只出现在河北和北京在内的温带地区或者是海拔约1 000 m 的亚热带地区,C.boninenses.l.只出现在湖南省和台湾省。HAN 等[56]研究发现由于各地温度湿度等气候条件的不同,炭疽菌的流行性和致病性也存在差异。预计在全球变暖的背景下,随着农业系统的更新,草莓炭疽病病原体的多样性和全球分布正在经历丰富的变化。
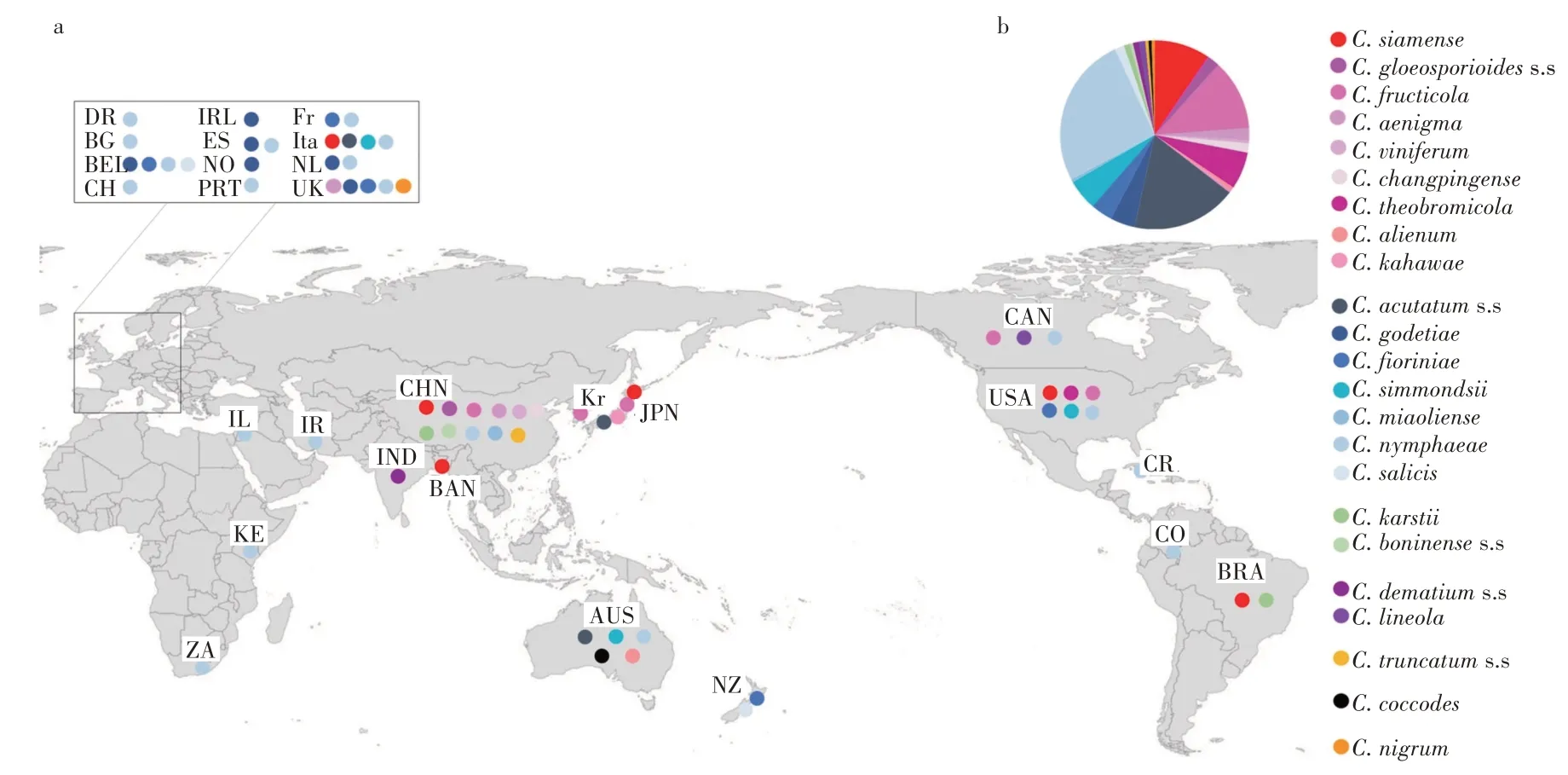
图8 危害草莓的炭疽菌在全球分布现状[25]Figure 8 Current status of the global distribution of Colletotrichum species affecting strawberry
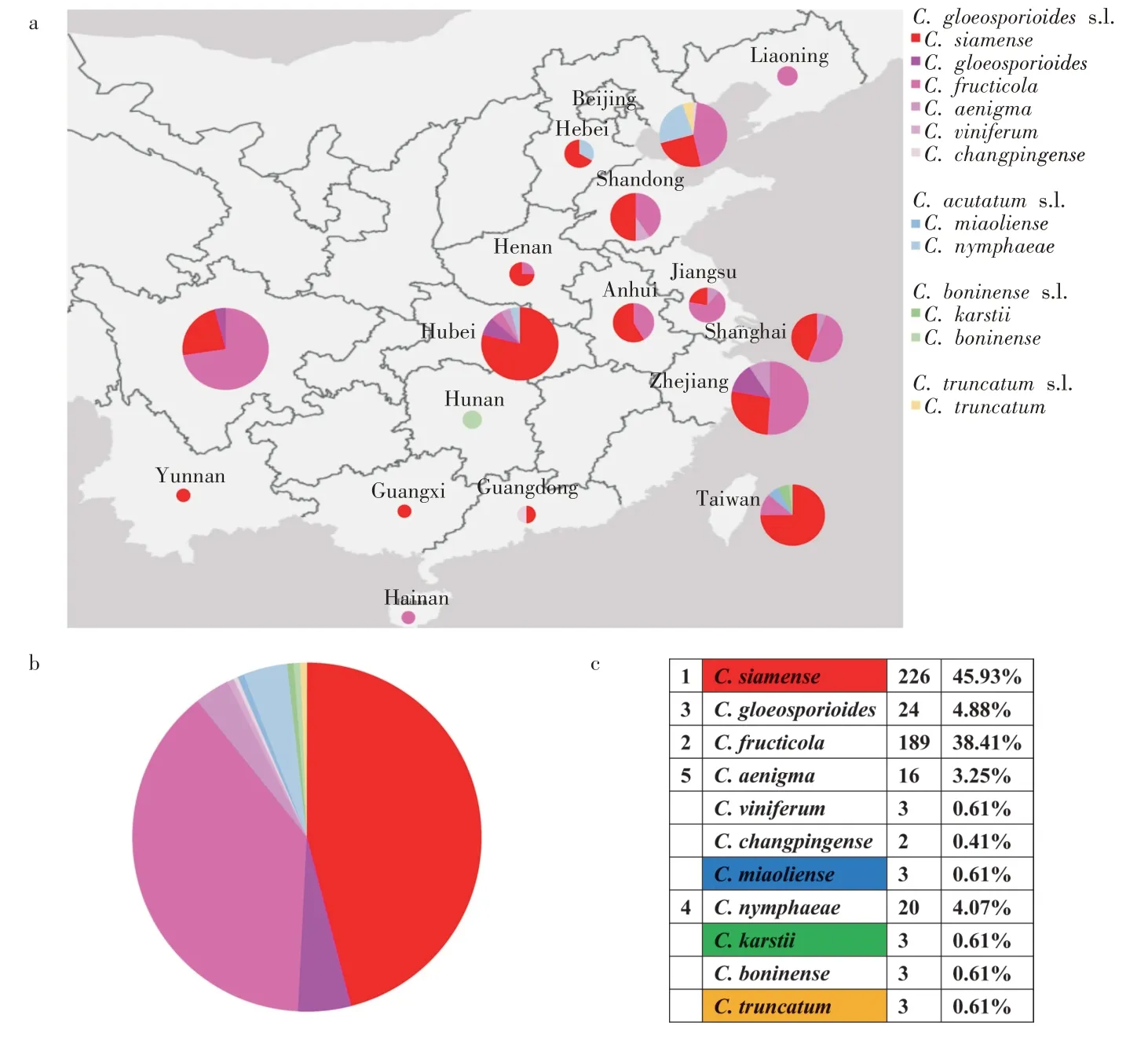
图9 草莓炭疽病病原菌在中国的分布[25]Figure 9 Distribution of strawberry anthracnose pathogens in China
4 防治方法
草莓炭疽病已经成为危害草莓生产的主要病害之一[57-58]。在高温高湿条件下,草莓炭疽病会在短时间内爆发,造成毁灭性的损失。草莓的叶片、匍匐茎、叶柄、花瓣和果实都可能被炭疽菌感染[59]。目前主要从农业防治、化学防治和生物防治3个方面控制草莓炭疽病在田间的流行和蔓延。
4.1 农业防治
4.1.1 选用抗病品种 我国南北方气候条件差异较大,各个地区栽种的草莓品种各不相同。然而,不同的草莓品种,对炭疽病的抗性也存在差异,因此需要评价各个品种对草莓炭疽病的抗性强弱。‘甜查理’品种对炭疽病的抗性较好,而‘红颜’和‘丰香’容易感染炭疽病。真菌侵染植物通常会引起相关酶活性的改变,研究表明,‘晶玉’品种在接种炭疽菌后,由于生理因子含量增加,对炭疽病有较强的抵抗性[60]。‘申阳’品种在生长过程中耐受高温,在幼苗期和结果期对炭疽菌具有一定的抵抗能力[61]。靳宝川等[62]通过研究11个草莓品种在田间对草莓炭疽病的抗性发现,同一个品种的草莓不同部位的发病情况不同,‘莫哈维’和‘叙利亚’品种的叶柄比叶片和匍匐茎更容易感染炭疽菌,‘波特拉’品种的匍匐茎更容易感病。
4.1.2 加强田间管理 清除病株:及时清理病果、落叶并集中烧毁。尽量减少病原体的数量,降低炭疽病在田间的流行与蔓延。
轮作栽培:草莓栽培过程中建议进行4 年以上的轮作栽培,目的是减少土壤中的致病真菌的数量,提高草莓的产量和品质[63]。
土壤消毒:对于栽培时间较长的土壤需要进行高温消毒灭菌[64],以降低炭疽病发生的几率。溴甲基、适量氯化钙对炭疽病都有一定的防治效果。
温湿度管理:温室栽培草莓对温度和湿度要求较高。草莓栽培初期,白天棚内温度应该保持在25~28 ℃,夜间棚内温度保持在8~12 ℃。开花期和结果期时,夜间栽培温度应该保持在5~10 ℃[64]。合理改善温室中通风透光条件,避免栽培过程中高温高湿现象出现,合理灌溉,及时排水。
4.2 化学防治
温室栽培过程中可以使用化学杀菌剂控制草莓病害的发生[65]。目前生产上广泛使用的杀菌剂主要有咪鲜胺、氟啶胺、代森锰锌和苯醚甲环唑等。姜莉莉等[66]通过室内平板试验研究表明,咯菌腈对草莓炭疽菌的抑制效果最好,咪鲜胺和苯醚甲环唑次之。田间盆栽试验结果表明,肟菌·戊唑醇和咪鲜胺防效均高于苯醚甲环唑水分散粒剂、40%福美锌可湿性粉剂和80%代森锰锌可湿性粉剂[67]。除此之外,酚类酸中反式肉桂酸(TA)具有控制草莓炭疽冠腐病发生的潜力,可以通过调节反式肉桂酸的含量来控制草莓炭疽病、冠腐病的发生,而不影响根系生长[68]。化学药剂的使用效果与气象条件存在一定关系。张叶根等[69]研究发现,在相同的气候条件下,考虑到经济成本,42.2%唑醚·氟酰胺(健达)和22.5%啶氧菌胺(阿砣)在温室中的防治效果较好。化学杀菌剂具有高效和便捷等优点,是农户的首要选择。但随着人们对食品安全和环境保护意识的增强,化学杀菌剂在农业中的使用逐渐受到限制[70]。因此,使用安全绿色的方法来控制疾病的发生和传播是至关重要的。
4.3 生物防治
随着草莓栽培面积的增加和人们对食品安全问题的关注,生产上迫切需要一种安全健康的病害防治措施。生物防治由于其安全和持久的特点,已成为一个新的研究热点[71-72]。目前用于防治草莓炭疽病的生物菌剂主要包括木霉菌和芽孢杆菌[73]。
木霉菌防治草莓炭疽病主要通过竞争作用、抗生作用和重寄生作用等方式实现。张雪等[74]发现使用绿色木霉菌剂处理感染胶孢炭疽菌的草莓叶片后,草莓叶片的病情指数降低并且激活了植物的防御系统。赵玳琳等[75-76]研究发现棘孢木霉可以接触、缠绕并穿透病原真菌菌丝,其发酵代谢物也能抑制病原菌生长最终导致菌丝死亡。MERCADO 等[77]报道哈茨木霉能够增加草莓对冠腐病的抗性,并且木霉菌剂对环境友好,作用时间长。陈娟等[78]在露地栽培和设施栽培两种方式下测定木霉菌Tr78对红颜草莓炭疽病的防治效果发现,Tr78菌株可以定植在草莓植株根部,并且定植数越高,对草莓炭疽病的防治效果越好。
芽孢杆菌被广泛用作农业的重要生物制剂[79],主要通过诱导系统抗性、拮抗作用及竞争作用防治草莓炭疽病。目前使用较多的芽孢杆菌主要有枯草芽孢杆菌、解淀粉芽孢杆菌和贝莱斯芽孢杆菌等[37,80]。吉沐祥等[81]研究发现枯草芽孢杆菌可以产生相关抑菌物质抑制病原菌的生长,并且室内平板试验的防治效果远高于盆栽试验,在田间浇施3次枯草芽孢杆菌后,草莓炭疽菌在田间的危害症状明显减弱。解淀粉芽孢杆菌具有提高草莓植株免疫力的能力[82]。解淀粉芽孢杆菌也具有控制炭疽病的能力,它可以产生脂肽类抗生素提高草莓的抗病性。研究表明,解淀粉芽孢杆菌抑制胶孢炭疽菌的能力主要取决于其生产Iturin A 的能力[83]。除此之外,贝莱斯芽孢杆菌含有与抗生物质合成相关基因srfAA、bymB、fenD、bacA和ituC,这些基因参与合成脂肽类抗生物质从而抑制炭疽病的发生。值得关注的是,有研究者将不同芽孢杆菌菌株混合使用去防治草莓田间病害。陈哲等[84]研究5 种芽孢杆菌按照不同比例施用对草莓炭疽病的防治效果发现,盆栽试验中复合菌对草莓炭疽病的防治效果为68.69%,与化学药剂防治相比有明显的抑制效果。同时,使用复合菌株有利于促进草莓植株的生长。
5 问题与展望
草莓炭疽病一直是草莓栽培过程中难以控制的危害性强的病害之一,由于地理位置和气候条件不同,病原菌的进化呈现多样性。随着病害的蔓延和流行,炭疽菌的分类越来越复杂,危害草莓的症状也各不相同。近年来,有许多学者致力于研究不同草莓炭疽病原菌的形态学鉴定,但是对病原菌的致病机制的认识非常有限。植物受到病原菌刺激后,通过自身细胞膜表面的受体蛋白识别致病菌激发自身第一层免疫系统(PTI)。除此之外,植物还存在第二层免疫系统(ETI),植物细胞内的一类受体蛋白能够特异性地识别病原菌的毒性蛋白植物,激发植物的特异性免疫。然而,当前对病原菌的致病机制还停留在病原菌触发植物的PTI 免疫途径中,对于ETI途径中植物抵抗病原菌侵染的机制还有待深入研究。
病害防治是生产中的关键问题。农业防治是综合防治的前提,选用抗病品种、采用适当的栽培措施是抗病栽培的重要手段。虽然农业防治容易推广,经济成本低,但地域性和季节性的差异,防治效果不稳定。化学防治中大量使用杀菌剂,虽然见效快、防治效果好,但是在杀死有害病菌的同时,会造成药剂残留,危害生态环境。随着人们食品安全意识的提高,化学药剂的使用范围逐渐受到限制。生物防治渐渐成为研究热点,使用有益微生物防治病害具有长期效应,在防治病害的同时还能促进植株健康生长。在生产中,生防菌剂的使用方法有待深入研究,在田间栽培管理的防治环节中仍然需要广泛推广,从而实现在草莓的生产栽培过程中对草莓炭疽病的有效预防及控制,提高草莓的产量及品质。
