芋扩展蛋白基因家族的全基因组鉴定及其在球茎膨大中的表达分析
2023-08-22李必聪李慧英周庆红黄英金朱强龙
李必聪,李慧英,肖 遥,罗 莎,周庆红,黄英金,朱强龙,*
(1.江西农业大学 农学院,江西 南昌 330045; 2.江西省薯芋生物学重点实验室,江西 南昌 330045)
扩展蛋白(expansin)是一类广泛存在于各种植物细胞壁中的伸展蛋白,已被普遍认为是植物细胞生长过程中细胞壁延伸的关键调节因子[1]。芋球茎膨大与其细胞生长和细胞壁延伸密切相关。因此,深入研究芋扩展蛋白基因家族可为芋球茎的生长发育、增产提质以及优良新品种选育提供理论依据。
扩展蛋白长度一般为250~275个氨基酸(aa),由两个主要的功能结构域(DPBB_1和Pollen_allerg_1)和氨基末端的一个信号肽组成,信号肽的长度通常为20~30个aa[2]。扩展蛋白基因家族可以根据其系统发育关系划分为EXPA、EXPB、EXLA和EXLB共4个亚家族(也称α、β、γ和δ亚家族)[3],其中EXPA和EXPB亚家族主要功能是促进植物细胞壁的伸展,并参与其他生长发育过程和抗性,但是关于EXLA和EXLB两个亚家族功能研究较少,尚未有明确的实验证据发现它们有增加细胞壁柔韧性的作用[2]。自1992年McQueen-Mason等[4]首次发现黄瓜幼苗细胞壁的粗蛋白提取物具有诱导离体细胞壁扩张的能力,从而鉴定出2个扩展蛋白以来,随着各种植物的基因组测序数据的不断完成与发表,在越来越多的物种中鉴定出了扩展蛋白,如早期在拟南芥中鉴定到36个扩展蛋白基因、水稻有58个[2],玉米有88个[5]、大豆有75个[6]、番茄有38个[7]、烟草有52个[8]、马铃薯有34个[9]、冬枣有30个[10]、毛竹有82个[11]、茶树有40个[12]和油菜有109个[13]等,为进一步深入研究扩展蛋白功能奠定基础。
扩展蛋白基因在植物的许多发育过程中都发挥重要作用,如对地下根茎的调控作用,在马铃薯中发现StEXPA7和StEXPA8在幼小的膨大块茎中具有较高的基因表达水平,可能在马铃薯早期的块茎发育中发挥重要作用[14];蔡兆琴等[15-16]发现,扩展蛋白基因IbEXPA2参与甘薯块根生长膨大调控,而IbEXPA4可以促进块根的纵向生长和横向生长。对植物细胞生长的调控作用,Lv等[17]的研究表明,分别存在3个、9个和2个扩展蛋白基因在棉花纤维细胞发育的起始阶段、快速伸长阶段和纤维生长的过渡阶段发挥重要作用;也有研究发现,AtEXPA10和PnEXPA1的转基因烟草植株与野生植株相比,其叶片、茎干和花朵显著增大,显微观察结果表明,膨大器官受细胞增殖刺激所致,且对PnEXPA1基因的过表达也会激活细胞扩增过程[18]。对植物细胞壁的调控作用,Wei等[19]研究发现,拟南芥中的AtEXPA1基因可以通过改变保卫细胞的细胞壁结构来调节气孔的开放;在中药材三七直根发育的研究中发现,PnEXP15、PnEXP17、PnEXP22和PnEXP24可能通过参与调控细胞壁松弛来促进其主根的膨大[20]。
芋[Colocasiaesculenta(L.)Schott]是天南星科家族中的一员,也是世界上最古老的作物之一,以其重要的食用、药用和经济价值而广受人们的喜爱[21]。据FAO(联合国粮食及农业组织)最新统计数据显示,2021年全球芋头产量超过1 200万t,是世界第五大根茎类农作物,其中我国大陆地区的芋头年产量为160多万t,仅次于尼日利亚,是世界第二大芋头产出国。2021年已经公布的芋全基因组测序数据[21]为进一步深入研究芋扩展蛋白基因家族的进化和功能特征奠定了重要的研究基础。芋头作为重要的非谷类粮食作物,其球茎富含易消化的淀粉颗粒且能量值高,并含有钾、磷、镁、锌和铁等对身体有益的微量元素[22-23],对于全球粮食安全起到重要的调节作用。但与产量直接相关的球茎(芋头)形成分子机理研究鲜有,而在马铃薯[9]、甘薯[15]、萝卜[24]、葛根[25]等同为根茎类作物的扩展蛋白基因的研究已经比较深入了,所以对芋中的扩展蛋白基因进行系统而深入的研究具有重要意义。
本研究基于芋全基因组测序数据,对芋扩展蛋白基因家族进行筛选鉴定,从基因结构特征、蛋白理化特性、系统发育进化、基因共线性关系、基因转录表达模式及实时荧光定量PCR(qRT-PCR)验证等方面对芋扩展蛋白基因家族的特征特性、系统进化及其在球茎膨大中的分子功能进行分析,为进一步研究扩展蛋白在芋球茎膨大发育等生物学过程中的功能奠定基础。
1 材料与方法
1.1 芋扩展蛋白基因家族成员的鉴定与结构分析
为获得全部的芋扩展蛋白基因,本研究从国家基因库核酸序列归档系统(CNSA)(https://ftp.cngb.org/pub/)中获取了芋基因组及氨基酸序列。扩展蛋白基因特有的保守结构域DPBB_1(pfam03330)和Pollen_allerg_1(pfam01357)均从网站http://pfam.xfam.org/获取。然后利用TBtools软件[26]中的Simple Hmm Search功能,基于隐马尔可夫模型,在芋蛋白质序列数据库中搜索扩展蛋白基因,选取满足E-value≤1×10-5条件的蛋白序列作为候选序列,再利用Pfam[27]和SMART[28]网站共同检验这些候选基因序列的特征结构域,并剔除未同时包含DPBB_1和Pollen_allerg_1两种特有保守结构域的基因序列。最后使用TBtools软件中的Visualize BCBI CDD Domain Pattern功能对扩展蛋白基因进行结构域的可视化分析,最终确定芋扩展蛋白基因的数量。此外,再利用芋全基因组的注释文件,通过TBtools软件中的Visualize Gene Structure(Basic)分析每个基因的内含子与外显子结构并对其进行可视化。
1.2 芋扩展蛋白序列分析及理化特征分析
将鉴定出的31个芋扩展蛋白基因家族成员的蛋白序列通过生物信息学网站ProtParam tool[29]分析每个蛋白的物理与化学参数;接着再利用网站Softberry(http://www.softberry.com)预测每个蛋白的亚细胞定位信息;在SignalIP-6.0[30]在线网站上分析信号肽的长度;并在MEME[31]信息网站上分析芋扩展蛋白基因的基序结构,将最大模体数量设为10,其他参数默认。
1.3 芋扩展蛋白基因系统发育树的构建及基因命名
首先通过MEGA11软件[32]中的Muscle序列比对功能对芋、拟南芥、水稻和马铃薯的扩展蛋白基因家族的氨基酸序列进行序列比对,然后将比对后的结果利用MEGA11软件中的Construct /Test Neighbor-Joining Tree功能(bootstrap值设置为1 000)构建系统发育树,再利用FigTree v1.4.4软件对系统发育树进行美化,从而将鉴定出的芋扩展蛋白基因家族成员进行亚家族分类,然后在不同的亚家族内根据其在染色体上的位置顺序进行命名。
1.4 芋扩展蛋白基因染色体定位和基因复制共线性分析
通过TBtools中的Blast Compare Two Seqs [sets]功能对芋扩展蛋白基因家族的氨基酸序列进行自我比对,并根据串联重复基因的筛选标准:(1)基因序列的比对率大于70%;(2)基因序列的比对相似性大于70%;(3)基因在染色体上的位置小于100 kb,鉴定出其中存在的串联重复基因,再利用国家基因库核酸序列归档系统(CNSA)中的染色体注释文件,使用TBtools软件中的Gene Location Visualize from GTF/GFF功能获得全部基因在染色体上的位置信息并进行可视化;利用MCScanX软件[33]对芋和拟南芥进行扩展蛋白基因成员倍增模式(基因复制共线性)分析,再利用Circos软件[34]绘制出两个物种的扩展蛋白基因在染色体上的物理位置及基因间的共线性关系。
1.5 芋扩展蛋白基因在不同组织中的转录表达分析
本实验室前期已经对地方芋品种江芋2号的叶片、根和3个不同发育时期的球茎(结芋期后8~10 d的子芋、50~60 d的子芋和90~100 d的子芋)进行了转录组测序。从转录组测序结果获得的芋不同组织中的芋扩展蛋白基因的转录表达谱数据(FPKM值),并分析家族各成员的表达趋势,再使用Clust vis在线网站[35]绘制出基因表达热图。
1.6 qRT-PCR分析
通过对芋扩展蛋白基因的转录表达谱热图和系统进化发育的分析,最终挑选出在芋球茎中表达差异最显著的基因(上调的有CeEXPA11、CeEXPA13和CeEXPA23;下调的有CeEXPA2和CeEXLA1)。再利用Prime Primer 5.0软件[36]对它们的CDS序列的保守部分设计特异性引物(表1)并送至北京擎科生物科技有限公司进行合成。使用TaKaRa MiniBEST Plant RNA Extraction Kit试剂盒对与转录组测序取样一致的芋球茎超低温保存材料的总RNA进行提取,再用TaKaRa PrimeScriptTMRT Master Mix试剂盒反转录成cDNA,以稀释4倍的cDNA为模板用于随后的qRT-PCR检测基因表达量。在BIO-RAD CFX96 Real-Time System仪器上完成实验,qRT-PCR反应体系为10 μL,包括2 μL的cDNA样品,各0.5 μL的正反向引物,5 μL的2×Realtime PCR Super mix,2 μL灭菌水。反应程序为:95 ℃ 30 s;95 ℃ 15 s,62 ℃ 15 s,72 ℃ 1 min,Plate Read,35个循环;熔解曲线在 65~95 ℃,increment 0.5 ℃ 0.05 s,Plate Read;结束。以actin基因作为内参对照,构建熔解曲线,使用2-ΔΔCT方法计算相对表达量,分析各基因的表达特征。使用SPSS 20进行单因素方差分析,Origin 2018进行图像绘制。

表1 扩展蛋白基因qRT-PCR引物
2 结果与分析
2.1 芋扩展蛋白基因的筛选
在芋全基因组中共筛选出31个扩展蛋白基因家族成员,经过BCBI batch CD-search 上的结构域分析,最终确定这31个基因均具有扩展蛋白基因的保守结构域。其中,EXPA亚家族有23个基因,EXPB亚家族有5个基因,EXLA亚家族有1个基因,EXLB亚家族有2个基因。按各基因在染色体上的位置分布,将这31个扩展蛋白基因依次命名为:CeEXPA1~CeEXPA23、CeEXPB1~CeEXPB5、CeEXLA1、CeEXLB1~CeEXLB2(表2)。
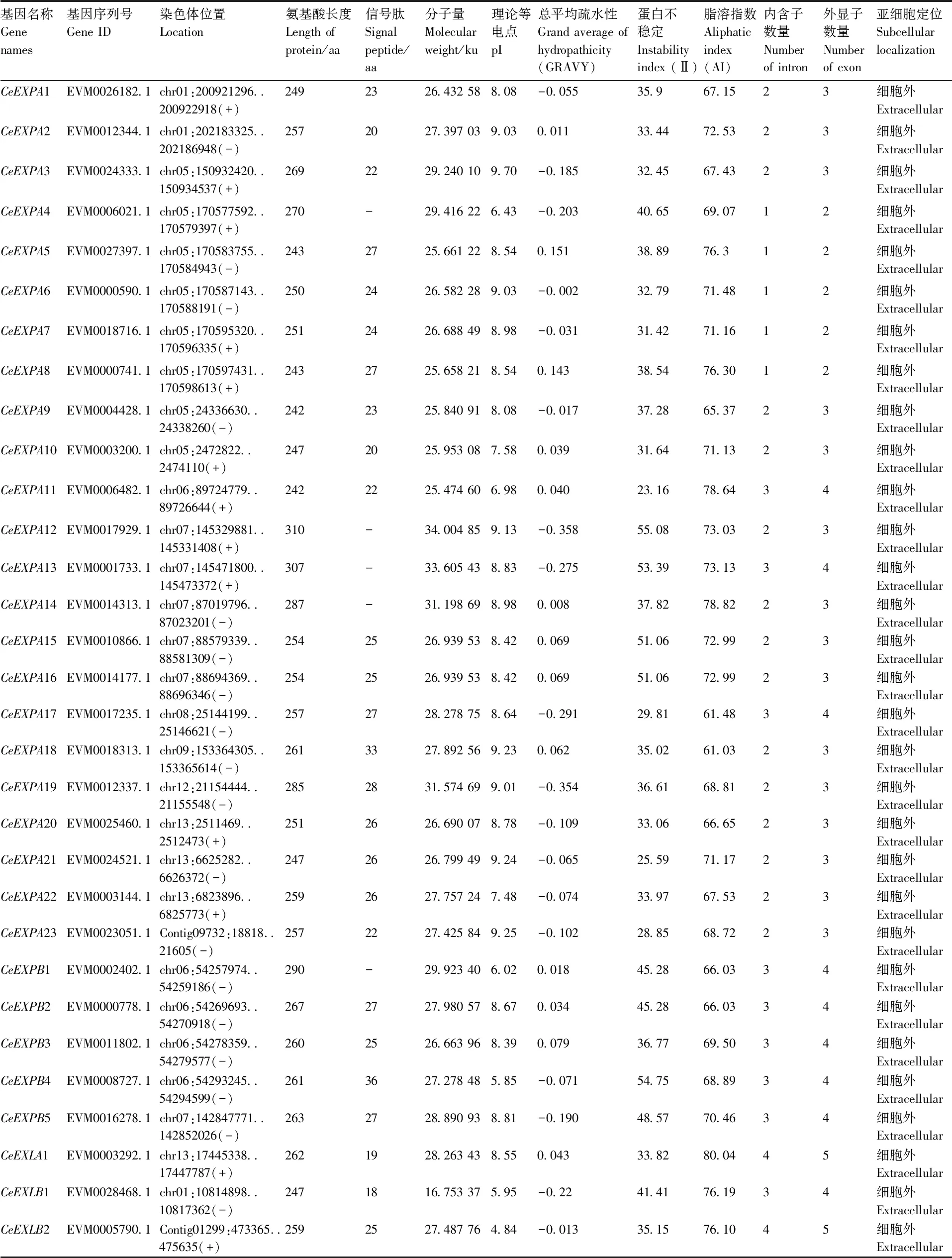
表2 芋扩展蛋白基因基本理化性质分析
2.2 芋扩展蛋白理化特征分析
蛋白质的功能特征与其理化特性密切相关。本研究鉴定出的31个扩展蛋白的分子量从16.75 ku到34.00 ku不等,差距达17.25 ku,平均值为27.64 ku;其蛋白质的最短氨基酸长度为242个,最长达310个,平均长度为261个,跨度也较大;经SignalP网站预测,共发现26个蛋白序列含有信号肽,长度范围在18~36个氨基酸之间,平均长度为25;利用Softberry网站预测,发现31个扩展蛋白均定位在细胞外。
结果表明,31个扩展蛋白的理论等电点最小值为4.84,最大值为9.70,平均值为8.18,其中有6个扩展蛋白的理论等电点均小于7,为酸性蛋白,而其余25个均为碱性蛋白。对总平均疏水性的计算发现,大于零的值(0.011~0.151)有13个,表明它们属于疏水性蛋白,而其余的18个蛋白(-0.358~-0.002)则属于亲水性蛋白。鉴定出的扩展蛋白不稳定指数在23.16~54.75,平均值为38.34,其中大于40的蛋白有10个,表明大部分扩展蛋白的结构稳定性较好(>40则不稳定)。这些基因的脂溶指数为61.03~80.04,平均值为70.84,相对较高的脂溶指数,保证该蛋白在各种环境中具有良好的稳定性,有利于其功能的正常发挥,其中CeEXLA1的脂溶指数高达80.04,属于嗜热型蛋白。
2.3 芋扩展蛋白基因结构分析
通过对芋扩展蛋白基因结构的分析,可以发现其中具有的内含子数量较少(表2、图1),但全部都具有内含子,为1~4个,平均为2.3个;其中5个基因(16.13%)含有1个内含子,15个基因(48.39%)含有2个内含子,9个基因(29.02%)含有3个内含子,只有2个基因(6.45%)含有4个内含子。而外显子数目多于内含子数量,为2~5个,平均数量为3.26个,且含有3~4个外显子的基因数量较多,达到24个(77.42%)。同时芋扩展蛋白基因的内含子与外显子结构呈现明显的亚家族特异性,4个亚家族的平均内含子数目分别为EXPA类1.91个,EXPB类3个,EXLA类4个,EXLB类3.5个。进一步分析发现,往往相同亚家族的基因都具有相似的长度和相同的内含子和外显子数量以及相似的保守结构域,如EXPA亚家族成员的大部分基因含有2个内含子,EXPB亚家族成员的基因都只有3个内含子,EXLA和EXLB两个亚家族更趋向于4个内含子,表明它们在进化上具有一定的保守性。
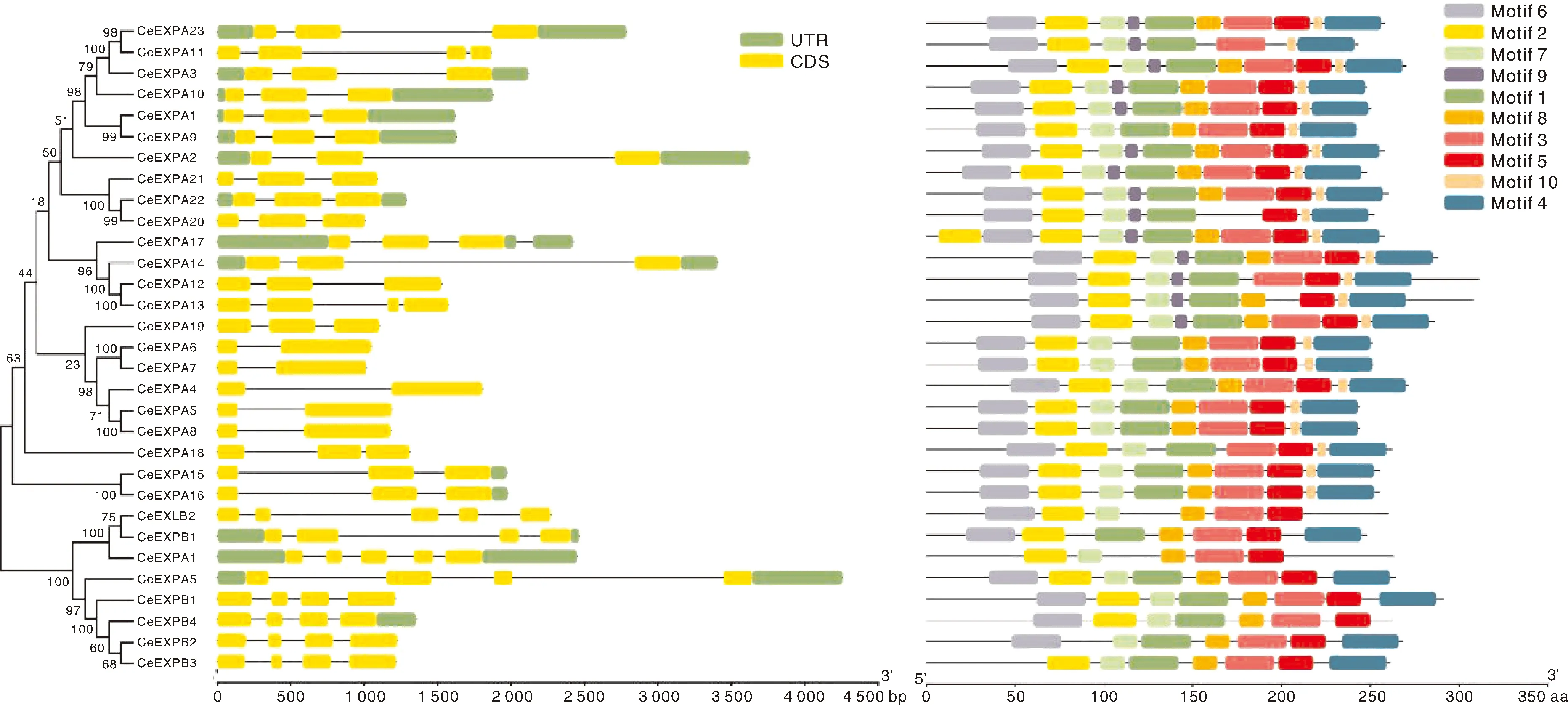
图1 芋扩展蛋白基因的进化与结构分析
通过MEME在线网站分析31个芋扩展蛋白基因的氨基酸序列,结果显示,在芋扩展蛋白基因中共找到269个预测的基序,每个基因含有5~10个基序,平均每个基因含有8.68个基序,其中CeEXLA1具有的基序数量最少,只有5个,而有10个EXPA亚家族的基因含有10个基序。4个亚家族中仅有EXPA亚家族含有Motif 9和Motif 10,且该亚家族中的成员都含有Motif 1、Motif 2、Motif 3、Motif 6和Motif 7,而其余3个亚家族则均含有Motif 3、Motif 5和Motif 8。可以发现,芋扩展蛋白基因在不同亚家族之间含有的基序数目和种类具有较大差异,这种基序的多样化也表明了芋扩展蛋白基因所具有的多种生物学功能。
2.4 芋扩展蛋白基因的染色体定位
根据芋扩展蛋白基因在染色体上的物理位置信息,分别将31个芋扩展蛋白基因定位于芋的8条染色体(chr1、chr5~chr9和chr12~chr13)以及2个重叠群(Contig01299和Contig09732)上。因此,31个扩展蛋白基因在染色体上的分布不均匀,其中5号染色体含有最多的基因(8个),7号、6号、13号、1号染色体分别含有6个、5个、4个和3个基因,而8号、9号和12号染色体以及Contig01299(CeEXLB2)和Contig09732(CeEXPA23)上分别只存在1个基因。从图2中还可以发现,CeEXPA4~CeEXPA8这5个基因在5号染色体上属于串联重复基因,同样在6号染色体上的CeEXPB1~CeEXPB4这4个基因也是如此,而且这些在染色体上紧密相邻的基因的序列相似性也比较高,所以它们在系统发育树上大都被聚类到一起。
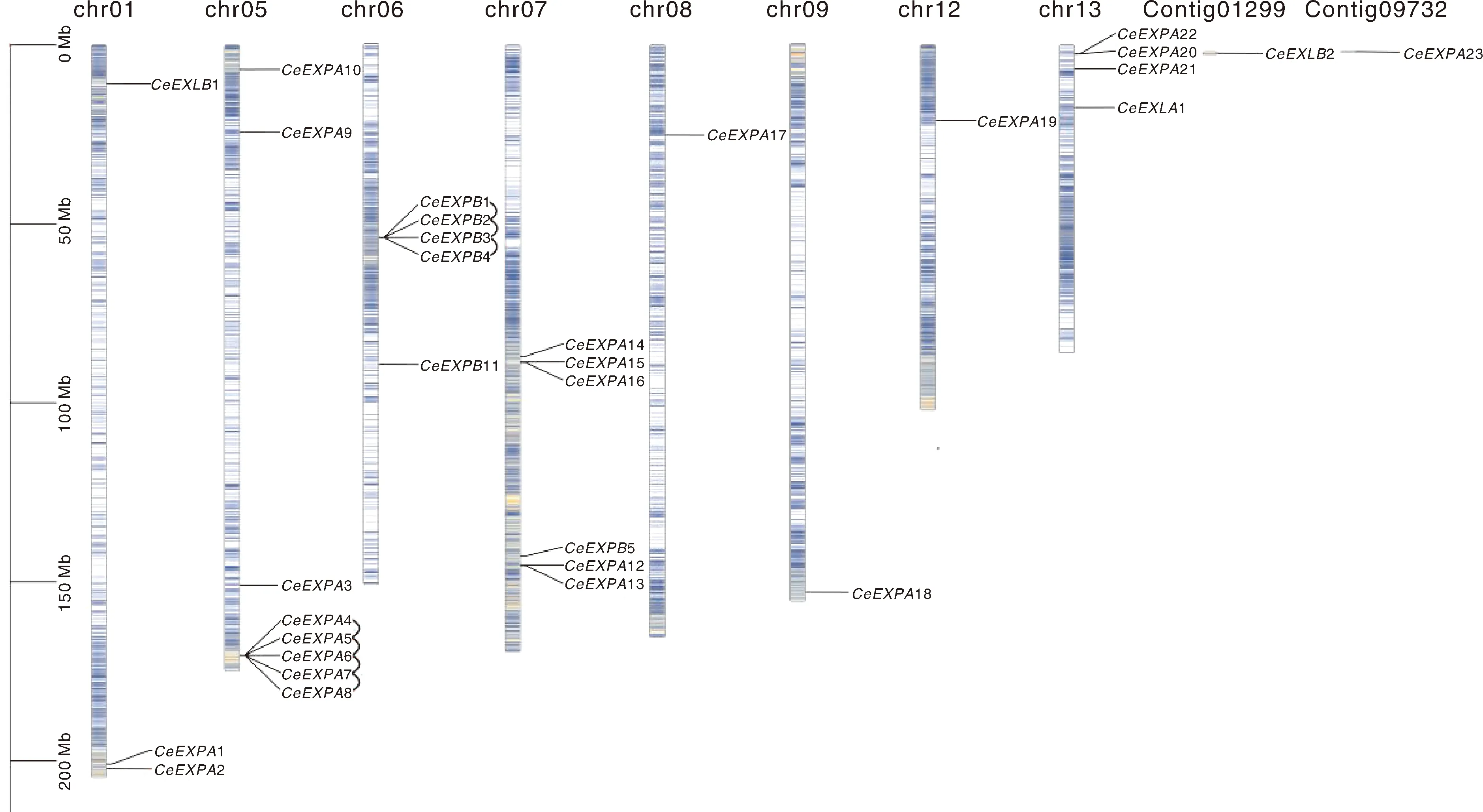
基因密度以热图的形式在染色体呈现,红色和蓝色区域分别表示高、低基因密度。黑线连接表示串联重复基因对。
2.5 芋扩展蛋白基因的系统发育树分析
利用MEGA 11软件对芋扩展蛋白基因进行了氨基酸多序列比对和系统进化树构建,分析了31个芋扩展蛋白基因家族成员之间的进化关系。结果表明,31个扩展蛋白基因被划分为EXPA、EXPB、EXLA和EXLB共4个亚家族(图3),其中EXPA亚家族有23个基因,EXPB亚家族有5个基因,EXLA亚家族有1个基因,EXLB亚家族有2个基因。从系统进化树中共发现芋中含有9个同源基因对,其中只有3个同源基因对的步长值小于90%,有4个同源基因对(CeEXPA5和CeEXPA8、CeEXPA11和CeEXPA23、CeEXPA12和CeEXPA13、CeEXPA15和CeEXPA16)的自展值为100%,而且都属于EXPA亚家族,2个同源基因对(CeEXPA6和CeEXPA7、CeEXPA20和CeEXPA22)的自展值为99%,其余3个同源基因对(CeEXPA1和CeEXPA9、CeEXPB2和CeEXPB3、CeEXPB1和CeEXPB4)的自展值分别为58%、53%和24%。
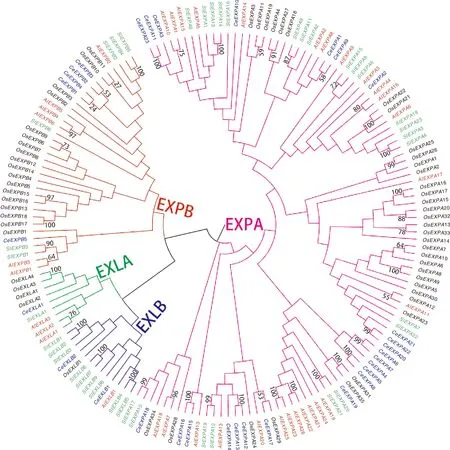
蓝色、红色、黑色和绿色的字体分别代表芋、拟南芥、水稻和马铃薯中的扩展蛋白基因。
为分析不同物种间扩展蛋白基因家族的进化关系,本研究选择芋、拟南芥、水稻和马铃薯的扩展蛋白基因序列构建系统进化树(图3)。通过系统进化树分析,4个物种的扩展蛋白基因均无一例外地被分到4个亚家族中,每一个亚家族均包含来自这4个物种的成员,这表明这一基因家族基本特征的形成早于单子叶与双子叶植物分化。通过进一步分析,在4个物种里面共鉴定出同源蛋白46对,其中物种间的直系同源蛋白2对(CeEXPA17和OsEXPA29、OsEXPA12和AtEXPA11),它们的蛋白序列相似性分别为53%和55%,而物种内的旁系同源蛋白44对,其中8对来自拟南芥,15对来自水稻,12对来自马铃薯,9对来自芋,它们各自的蛋白序列相似性为64%~100%、27%~100%、72%~100%和24%~100%,以及它们的平均蛋白序列相似性为86.00%、85.07%、95.58%和81.44%。进一步分析表明,被划分入4个亚家族的4个物种的扩展蛋白基因的分布都具有一定的相似性,其中EXPA亚家族含有最多的扩展蛋白基因(106个,占65.03%),EXLA亚家族中的扩展蛋白基因(9个,占5.52%)数量最少。还发现,芋的4个亚家族比例与拟南芥的比例最相似,其中EXLA和EXLB亚家族所占比例较小,但是同为块茎类作物的马铃薯却含有9个EXLA亚家族的扩展蛋白基因,所占比例也高达23.08%,是这4个物种里面最高的,而芋EXLA亚家族所占的比例(6.45%)在这4个物种中则仅次于马铃薯。
2.6 芋与拟南芥的扩展蛋白基因共线性分析
由于拟南芥是植物研究中最重要的模式植物之一,而且到目前为止对拟南芥中的扩展蛋白基因功能的研究也比较深入[37-40],所以将芋和拟南芥的扩展蛋白基因一起进行共线性分析,以便后续对芋的扩展蛋白基因功能进行预测。如图4,共线性分析的结果显示,在芋和拟南芥之间含有7对共线性基因(AtEXPA2和CeEXPA21、AtEXPA10和CeEXPA1、AtEXPA10 和CeEXPA9、AtEXPA13和CeEXPA10、AtEXPA18和CeEXPA1、AtEXPA18和CeEXPA9、AtEXLA3和CeEXLA1),在芋中发现1对共线性基因(CeEXPA1和CeEXPA9),在拟南芥中鉴定出9对共线性基因(AtEXPA3和AtEXPA5、AtEXPA1和AtEXPA4、AtEXPA7和AtEXPA8、AtEXPA9和AtEXPA14、AtEXPA10和AtEXPA18、AtEXPA8和AtEXPA17、AtEXPA14和AtEXPA17、AtEXPB4和AtEXPB5、AtEXPB3和AtEXPB6)。
2.7 扩展蛋白基因在不同芋组织中的转录表达量分析及qRT-PCR分析
利用从转录组数据中获取的芋扩展蛋白基因在不同组织中特异表达的FPKM值,绘制基因表达量热图并分析基因表达模式。在图5-A中,根据芋扩展蛋白基因在不同组织中的表达情况,大致可分为两类表达模式,一类是在球茎中表达量高且有显著性变化,同时在根和叶片中表达量相对较低的基因,如CeEXPA2、CeEXPA11、CeEXPA13、CeEXPA23和CeEXLA1等;第二类是在球茎中表达量低且没有显著性变化,但在叶片中高表达的基因,如CeEXPA10和CeEXPB5。进一步分析球茎膨大中第一类差异表达基因的表达趋势,可以发现,CeEXPA2和CeEXLA1在芋球茎发育过程中表达量先下降后略有升高;而CeEXPA11、CeEXPA13和CeEXPA23则在芋球茎发育过程中表达量呈不断升高的趋势。
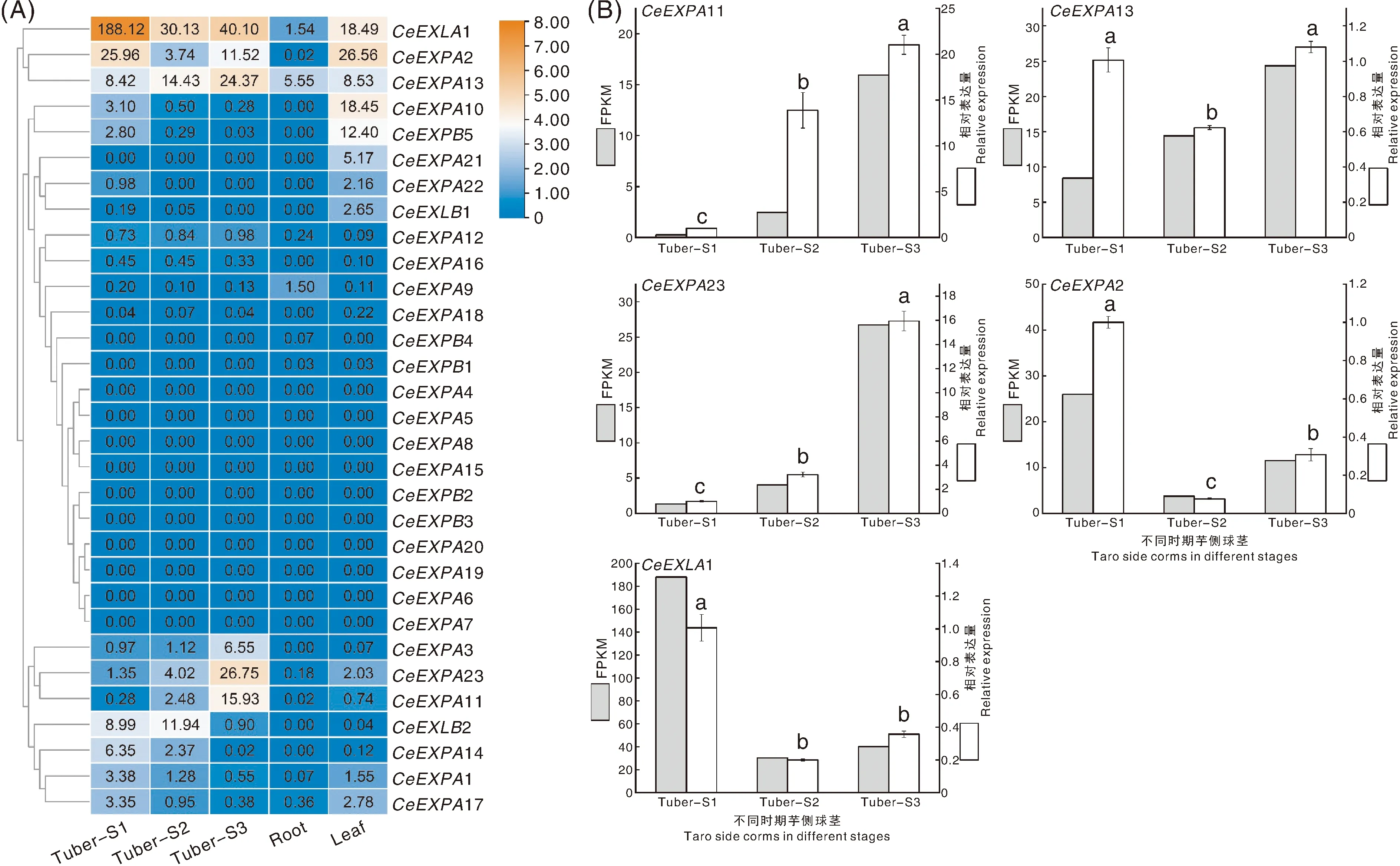
Tuber-S1,球茎膨大前期;Tuber-S2,球茎膨大中期;Tuber-S3,球茎膨大后期。不同小写字母表示不同发育时期芋球茎的相对表达量在0.05水平上的差异显著性。
然后利用qRT-PCR对芋球茎中存在的上调表达基因CeEXPA11、CeEXPA13和CeEXPA23,下调表达基因CeEXPA2和CeEXLA1进行基因相对表达量的验证,并与FPKM值进行对比、制图分析。本实验中验证的各基因的定量分析结果都与FPKM值的变化趋势基本保持一致,唯有CeEXPA13的定量结果变化趋势为先下降后升高,与FPKM值升高的趋势有所不同(图5-B)。
3 结论与讨论
扩展蛋白基因在植物基因组中是广泛存在的,其主要作用方式是疏松植物细胞壁的组分,增加细胞壁的柔韧性,从而调控植物的生长发育[41]。本研究在芋中鉴定了31个扩展蛋白基因,其中含有23个EXPA、5个EXPB、1个EXLA和2个EXLB。芋扩展蛋白基因数量与拟南芥(36)、杨树(36)、葡萄(29)、菜豆(36)、番茄(38)和马铃薯(36)相近。通过对拟南芥、水稻、马铃薯和芋4个物种扩展蛋白基因亚族大小的比较分析,发现每个扩展蛋白亚家族在物种间的分布是不均匀的,扩展蛋白亚家族基因在物种间的拷贝数差异可能是由于植物为适应不同环境需求导致的演化[7]。经对4个物种的扩展蛋白基因系统进化树分析,发现芋和同为单子叶植物的水稻之间具有更近的亲缘关系,且同为双子叶植物的拟南芥和马铃薯的扩展蛋白基因也具有更近的亲缘关系,由此推测,早在单子叶和双子叶分化之前,就有一组与每个亚家族相对应的祖先扩展蛋白基因[42]。芋EXPA亚家族基因占扩展蛋白基因家族总成员数的比例为74%,这一比例显著高于同为单子叶植物的水稻(59%)[43]和玉米(41%)[5]。此外,本研究鉴定出的9个芋扩展蛋白基因分布在2个串联重复基因对中,表明片段重复在一定程度上促进了芋扩展蛋白基因家族的扩展,该结论与Chen等[44]对其他40种代表性植物中扩展蛋白基因家族分布与扩张的研究结论相一致。
不同亚家族之间在基因内含子外显子结构和基序模式等方面具有一定的保守性,代表了亚家族的重要特征[45]。芋的29个扩展蛋白基因(93.54%)含有1~3个内含子,约77.42%含有3~4个外显子,尤其是其中EXPB亚家族成员都为4外显子/3内含子的结构,这一结果与番茄EXPB亚家族中内含子和外显子结构一致[7],这在一定程度上说明2个物种的EXPB亚家族基因在结构上具有较高的保守性;EXPA、EXPB、EXLA和EXLB共4个亚家族都具有的基序结构为Motif 2、Motif 3、Motif 5、Motif 7和Motif 8,而在马铃薯中则发现4个亚家族均含有Motif 3和Motif 6[45],可见芋的扩展蛋白基因在基序模式上具有更高的保守性;还发现EXLA亚家族唯一的家族成员CeEXLA1只含有5个基序,少于其他扩展蛋白基因。相同亚家族中的扩展蛋白基因含有相似的内含子与外显子结构和基序特征,而不同亚家族之间基序特征的多样性可能体现了芋扩展蛋白基因在细胞内的多种代谢途径和反应中都有广泛的参与度。
扩展蛋白基因在植物茎的生长发育过程中具有重要的调控作用[14]。CeEXPA2在芋球茎膨大前期高表达,然后随着球茎的膨大显著下调表达,同时,该基因也在芋叶片中有较高的表达量,而在根中的表达量较低,通过系统进化树的比较发现该基因与拟南芥中的AtEXPA3和AtEXPA16具有较近的亲缘关系,有研究发现AtEXPA3和AtEXPA16参与拟南芥根部的合胞体形成过程中的细胞生长和细胞壁分解,且在合胞体中特异表达,但在周围根组织中不表达[40],据此推测CeEXPA2可能对芋球茎和叶的生长发育都有重要的调控作用,在球茎膨大过程中可能通过调控细胞发育和细胞壁修饰,从而促使球茎膨大。CeEXLA1在叶片中的表达量高于根,但都远低于球茎,且在芋球茎膨大过程中下调表达,作为芋EXLA亚家族唯一的基因,其在系统进化树中与OsEXLA1、OsEXLA2和OsEXLA3的亲缘关系较近,在水稻中就发现OsEXLA3的表达与细胞伸长相关,且受到赤霉素的诱导[46],表明CeEXLA1可能在芋球茎膨大过程中调控细胞伸长增大,从而促使球茎膨大。CeEXPA11和CeEXPA23在聚类分析中的亲缘关系最近,且它们的表达量都在芋球茎发育过程中呈现不断增高的趋势,而在叶片和根中的表达水平较低,在聚类上发现它们与StEXPA1和StEXPA5存在较近的亲缘关系,有研究结果显示,StEXPA1在马铃薯块茎发育过程中的表达量先下降后上升,StEXPA5的表达量则呈现不断升高的趋势[14],这与本研究结果相似,表明CeEXPA11和CeEXPA23可能共同调控芋球茎的膨大生长,且在膨大后期发挥着重要的作用。CeEXPA13的表达量从芋球茎膨大初期开始不断升高,且在根和叶中均存在一定的表达量,但显著低于球茎中的表达量,从聚类分析又发现其与StEXPA12和StEXPA19具有较近的亲缘关系,理向阳等[45]对马铃薯扩展蛋白基因在15个不同组织中的表达分析,发现StEXPA12在块茎中的表达量非常高,推测CeEXPA13可能对芋球茎膨大也有重要的调控作用。
综上所述,从芋中鉴定出的31个扩展蛋白基因在基因结构上具有一定的保守性,与拟南芥、水稻及马铃薯中的扩展蛋白基因存在较近的亲缘关系。芋扩展蛋白基因表达模式和功能预测分析发现该家族基因不同成员在芋不同组织中有不同的表达模式,其中CeEXPA2、CeEXPA11、CeEXPA13、CeEXPA23和CeEXLA1可能对芋球茎的膨大发挥重要的调控作用。本研究将为进一步解析芋球茎膨大形成机理及选育高产优质芋品种奠定理论基础。
