浙北地区红美人杂柑成花过程及其生理生化特征
2023-08-22王智豪奚昕琰杨淑娜高志远殷益明贾惠娟
王智豪,奚昕琰,王 莉,杨淑娜,高志远,殷益明,邹 辉,贾惠娟,*
(1.浙江大学 农业与生物技术学院,浙江 杭州 310058; 2.湖州市农业科技发展中心,浙江 湖州 313000; 3.南浔双林得源家庭农场,浙江 湖州 313012)
红美人是从日本引进的橘橙类杂交新品种,杂交母本为南香,父本为天草,登记名为爱媛果试第28号,简称爱媛28。红美人最适宜种植在年平均气温16~22 ℃,年光照时间大于1 300 h,极端最低温度(-5 ℃)持续时间不超过24 h,土壤pH值4.5~8.0的地区[1]。果实成熟期在10月下旬,12月中旬完熟[2]。该品种果面呈橙红色,果肉化渣性好,优质高糖,有甜橙般香气,深受消费者的喜爱。不同于其他果树,柑橘具有一年抽生多次枝梢的特点,按发生时期可分为春梢、夏梢、秋梢,它们均可成为结果母枝,但成花、坐果和果实品质在不同种类结果母枝上存在差异。目前在相对冷的浙江北部地区也有红美人引种,但引种时间短,栽培管理上多参照浙南地区其他柑橘的传统方法,少有针对性的技术,存在中、后端枝梢较前端枝梢花量少、成花能力弱等问题,导致结果部位外移、树势早衰现象突出,制约了红美人产业的高质量发展。
花芽分化是果树成花过程的基础,与果实的产量和品质密切相关[3]。大量研究表明,内源激素对调控植物花芽分化有重要作用,这在杏[3]、桃[4]、梨[5]、葡萄[6]、樱桃[7]等果树上已有报道。高ABA/GA(脱落酸含量/赤霉素含量)、ZR/GA(玉米素核苷含量/赤霉素含量)有利于成花[3-7],而IAA/GA(生长素含量/赤霉素含量)对花芽分化的作用尚未有定论。另外,在桃[4]、荔枝[8]、杨梅[9]、刺梨[10]中,碳氮物质的总含量之比决定植株成花。目前,已有不少学者对脐橙[11-14]、蕉柑[15]、金柑[16]、琯溪蜜柚[17]、无籽蜜柚[18]等柑橘的花芽分化进行了研究,但多集中在对花芽形态分化进程的观察,对其调控机制鲜有报道。红美人杂柑是近年来产业价值颇高的新品种,研究其花芽分化进程、成花规律对探索配套技术、高质量引种意义重大。
本研究以红美人为材料,通过观察其花芽分化进程,并检测不同结果母枝花芽的内源激素含量和叶片碳氮总含量,分析内源激素、营养物质变化与成花之间的关系,初步阐明不同结果母枝花芽分化进程的生理机制,旨在为红美人的成花调控和栽培管理技术提供理论依据。
1 材料与方法
1.1 材料
试验于2022年在浙江省湖州市南浔双林得源家庭农场进行。该地属于亚热带季风气候,年平均气温16.9 ℃,年平均降水量1 023.3 mm,年平均日照为1 778.0 h。大棚设施栽培(10月初盖膜,翌年3月初撤膜),果园管理良好。
供试材料为6年生红美人杂柑,基砧为枳,中间砧木为温州蜜柑,于花芽萌发前随机剪取各种类型枝条各15枝,为方便表述,将其分别简写为:SSA-SP(三次梢春梢)、SSA-SU(三次梢夏梢)、SSA-AU(三次梢秋梢)、SS-SP(二次梢春夏梢-春梢)、SS-SU(二次梢春夏梢-夏梢)、SA-SP(二次梢春秋梢-春梢)、SA-AU(二次梢春秋梢-秋梢)、S-SP(单次梢-春梢)(图1)。用刀片将各类枝梢前端5个芽从采集的枝条上切下,混合均匀后各取25枚芽用于花芽形态分化的观察,称取200 mg花芽用于测定内源激素含量。将叶片从采集的枝条上剪下,用于测定碳氮含量。具体取样时间为:1月5日、1月20日、2月5日、2月20日、3月6日。叶片碳氮含量的测定取样时间在以上基础上增加3次,分别为3月21日、3月31日和4月10日。
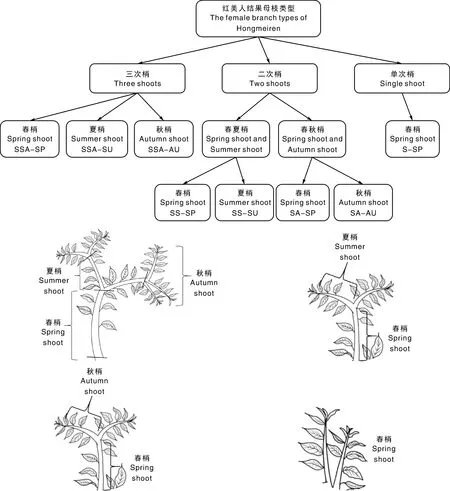
图1 取样示意图
1.2 花芽形态分化进程观察
用刀片从采集的枝条上将花芽切下,立即用O.C.T包埋液(optimal cutting temperature compound)进行包埋后放入-80 ℃冰箱冷冻保存。花芽切片使用Leica-CM1950冷冻切片机,切片厚度设置为15 μm,机箱温度-20~-22 ℃。取花芽中间段的切片在Leica DM 1000 LED显微镜下拍照,观察其花器官的形态变化、花芽分化进程并统计各个阶段的分化百分率。
1.3 花芽内源激素含量测定
将采集的花芽用液氮速冻后,使用JX-FSTPRP-24全自动样品快速研磨仪(上海净信科技)研磨成粉末。植物材料内源激素的提取方法和液相色谱-质谱联用分析方法参照文献[4]的方法。
1.4 叶片总碳含量和总氮含量测定
将叶片剪碎混合均匀,取2 g左右在60 ℃烘干至恒重,使用JX-FSTPRP-24全自动样品快速研磨仪(上海净信科技)研磨成粉末,过50目筛,冷却至室温后,准确称取4 mg(精确到0.1 mg)用vario MICRO CHN元素分析仪测定样品总碳含量和总氮含量。
1.5 数据分析
实验数据采用Excel 2020软件整理记录,使用SPSS 24.0软件进行统计分析,用Ducan’s新复极差法在α=0.05水平上进行比较,采用GraphPad Prism 9软件制图。
2 结果与分析
2.1 红美人杂柑花芽分化时期与分化特征
红美人杂柑在湖州地区的花芽分化进程共分6个时期(图2):未分化期(图2-a),此时红美人杂柑的芽外形瘦小,不突出,外层鳞片排列紧密,芽的生长锥尖狭小。分化初期(图2-b),突起的生长点先端变得宽而肥大,向上隆起呈馒头状,以后逐渐伸长、变宽,顶端逐渐扁平;鳞片开始变松,花原基形成。萼片分化期(图2-c),芽的鳞片更加松散,顶端迅速膨大,顶部增宽,四周产生突起,即萼片原基,标志着进入了萼片分化期,分化后的生长点变宽而扁平。花瓣分化期(图2-d),随着萼片原基的伸长,萼片内侧基部发生突起体,即为花瓣原基;花瓣原基不断伸展,互相抱合。雄蕊分化期(图2-e),在花瓣原基内侧分化出新的突起,即为雄蕊原基,雄蕊伸长,中心盘体出现,雄蕊原基向上生长。雌蕊分化期(图2-f),在雄蕊原基下方,生长锥中央逐渐向上突起,形成雌蕊原基,此时花芽进入了雌蕊分化期。
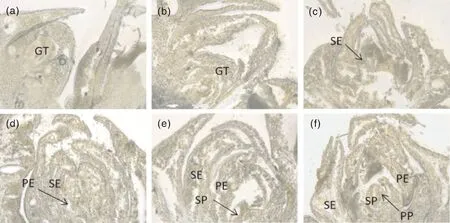
a,未分化期;b,分化初期;c,萼片分化期;d,花瓣分化期;e,雄蕊分化期;f,雌蕊分化期;GT,生长锥;SE,花萼原基;PE,花瓣原基;SP,雄蕊原基;PP,雌蕊原基。
2.2 红美人花芽分化进程
柑橘的花芽分化过程包括生理分化、形态分化、性细胞形成3个阶段。分化初期之前属于生理分化期;进入萼片分化期,标志着花芽分化进入了形态分化期[19]。在同一取样时间,同类型结果母枝上存在处于不同分化时期的花芽。
由图3可知,1月5日各种类型枝梢上存在未分化期、分化初期、萼片分化期的花芽,而SSA-AU、SS-SU、SA-AU、S-SP出现了花瓣分化期,占比分别为19.0%、8.7%、19.0%、22.7%。1月20日,SSA-SU、SS-SP、SA-SP上首次出现了花瓣分化期,SSA-SP上仍未出现,而此时SSA-AU、SS-SU、SA-AU、S-SP上花瓣分化期的占比已分别达到39.1%、23.8%、36.3%、47.6%。2月5日,SSA-AU、SS-SU、SA-AU、S-SP出现了雄蕊分化期,各自占比为33.3%、19.0%、26.1%、38.1%,此时SSA-SP出现了花瓣分化期。2月20日,雌蕊分化期最先在SSA-AU、SS-SU、SA-AU、S-SP上出现,分别占比为9.5%、9.1%、9.1%、13.6%。3月6日,SSA-AU、SS-SU、SA-AU、S-SP上分别有25.0%、22.7%、21.7%、22.7%的花芽进入了雌蕊分化期,而此时SSA-SP、SSA-SU、SS-SP、SA-SP上分别仅有4.8%、4.8%、9.1%、9.5%的花芽进入了雌蕊分化期。
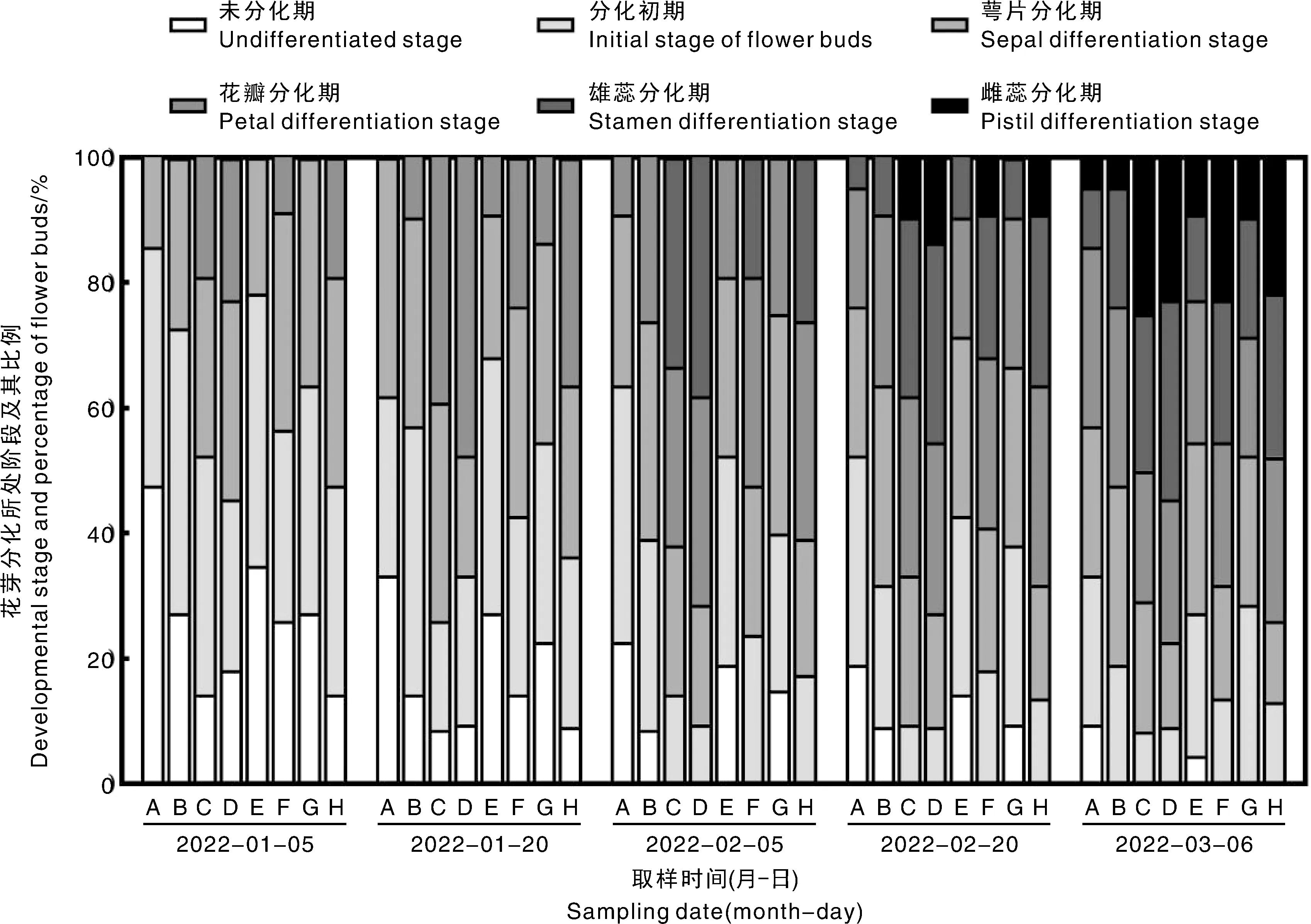
A,SSA-SP(三次梢春梢);B,SSA-SU(三次梢夏梢);C,SSA-AU(三次梢秋梢);D,SS-SP(二次梢春夏梢-春梢);E,SS-SU(二次梢春夏梢-夏梢);F,SA-SP(二次梢春秋梢-春梢);G,SA-AU(二次梢春秋梢-秋梢);H,S-SP(单次梢-春梢)。
以上结果表明,各种类型枝梢花芽分化进程存在差异,总的趋势是枝条中、后端梢较前端梢花芽分化进程慢且最后花芽处于雌蕊分化期的比例较低。即SSA-AU、SS-SU、SA-AU、S-SP进程较快,且最后花芽处于雌蕊分化期的比例较高;而SSA-SP、SSA-SU、SS-SP、SA-SP上花芽分化进程较慢,且最后花芽处于雌蕊分化期的比例较低,这可能会影响后续成花过程。
2.3 红美人成花过程中花芽激素含量变化
由图4可知,花芽中的ABA含量呈现先下降后上升的趋势。三次梢中SSA-SU、SSA-AU上ABA含量的转折点出现在1月20日,此时SSA-SU、SSA-AU上的花芽进入了花瓣分化期;SSA-SP的转折点出现在2月20日,此时SSA-SP处于花瓣分化期,仅有极少数花芽进入了雄蕊分化期。二次梢中,SS-SP、SS-SU的ABA含量在2月5日快速上升,此时SS-SP进入了花瓣分化期,SS-SU进入了雄蕊分化期;SA-SP、SA-AU的波动不大。单次梢呈现同样的规律,ABA含量先降后升,2月5日后快速上升,此时花芽处于雄蕊分化期。
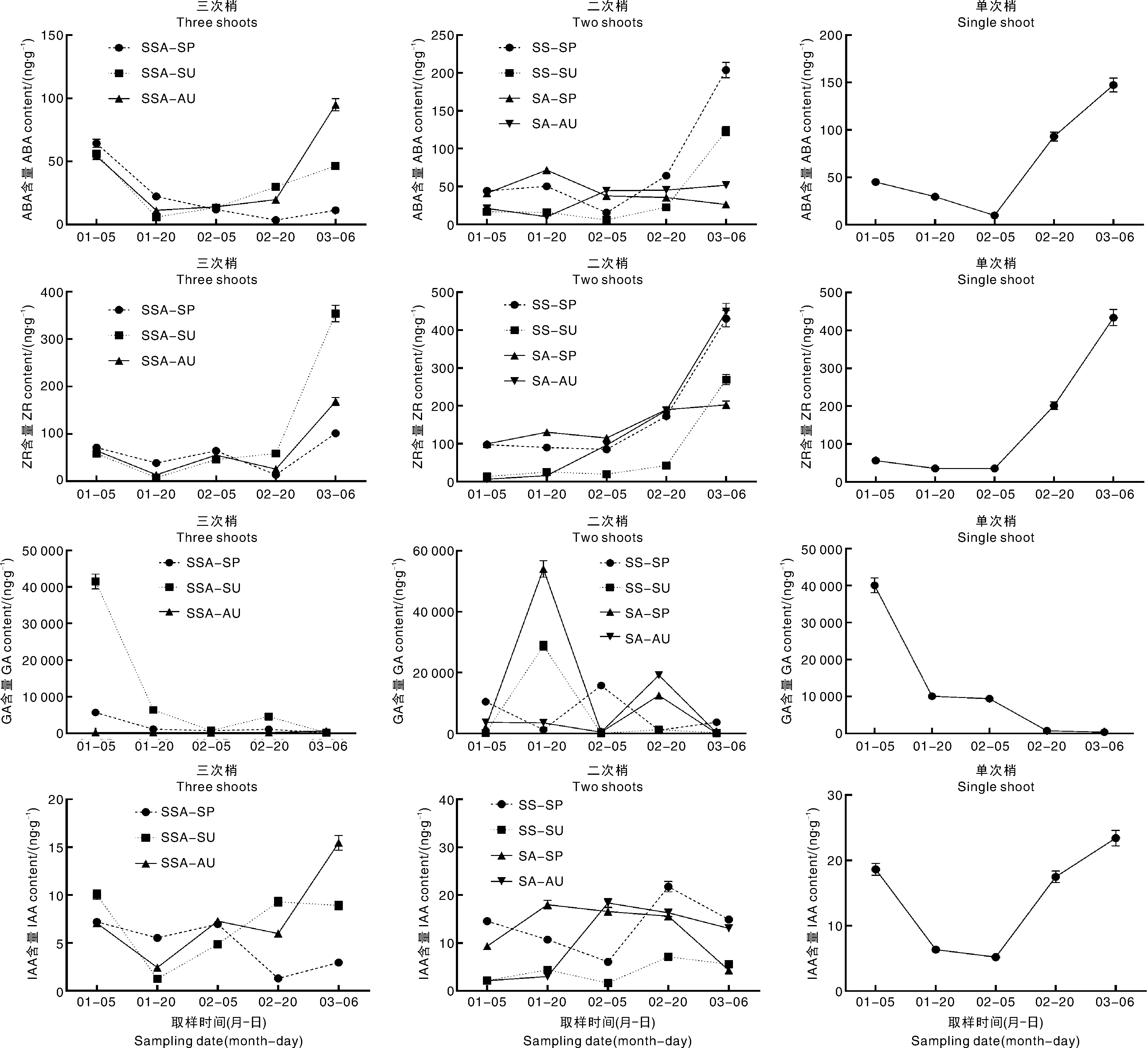
SSA-SP,三次梢春梢;SSA-SU,三次梢夏梢;SSA-AU,三次梢秋梢;SS-SP,二次梢春夏梢-春梢;SS-SU,二次梢春夏梢-夏梢;SA-SP,二次梢春秋梢-春梢;SA-AU,二次梢春秋梢-秋梢;S-SP,单次梢-春梢。下同。
花芽中的ZR含量呈现波动上升的趋势。三次梢中,各类枝条ZR含量均在2月20日快速上升,此时SSA-SP、SSA-SU刚进入雄蕊分化期,SSA-AU刚进入雌蕊分化期。二次梢中呈现类似的规律,除SA-SP外,ZR含量均在2月20日快速上升,此时SS-SP进入了雄蕊分化期,SS-SU和SA-AU进入了雌蕊分化期。单次梢中,ZR含量于2月5日快速上升,此时花芽处于雄蕊分化期。
花芽中GA含量呈现下降的趋势。三次梢中SSA-SP、SSA-SU、SSA-AU,以及单次春梢均在1月20日快速下降,随后GA一直处于较低水平。二次梢中,GA含量波动变化,大致在1月20日和2月20日出现2个峰值。
花芽中IAA含量波动变化,未发现明显规律。
以上结果表明,ABA含量升高时花芽多处在花瓣分化期和雄蕊分化期,ZR含量升高时花芽多处在雄蕊分化期和雌蕊分化期。
2.4 红美人成花过程中花芽激素比值变化
由图5可知,1月5日至2月20日,三次梢中SSA-AU的ABA/GA、ZR/GA、IAA/GA均高于SSA-SP、SSA-SU;2月20日,SSA-SP、SSA-SU中的ABA/GA、ZR/GA、IAA/GA快速升高,最终SSA-SU高于SSA-SP。就二次梢而言,ABA/GA、ZR/GA、IAA/GA呈现类似的规律:均在2月5日出现一个小高峰,2月20日,ABA/GA、ZR/GA、IAA/GA快速上升,最终SS-SU高于SS-SP,而SA-SP和SA-AU差异不显著。就单次梢而言,ABA/GA、ZR/GA、IAA/GA均在2月5日快速上升,可能与此时花芽分化进程有关。
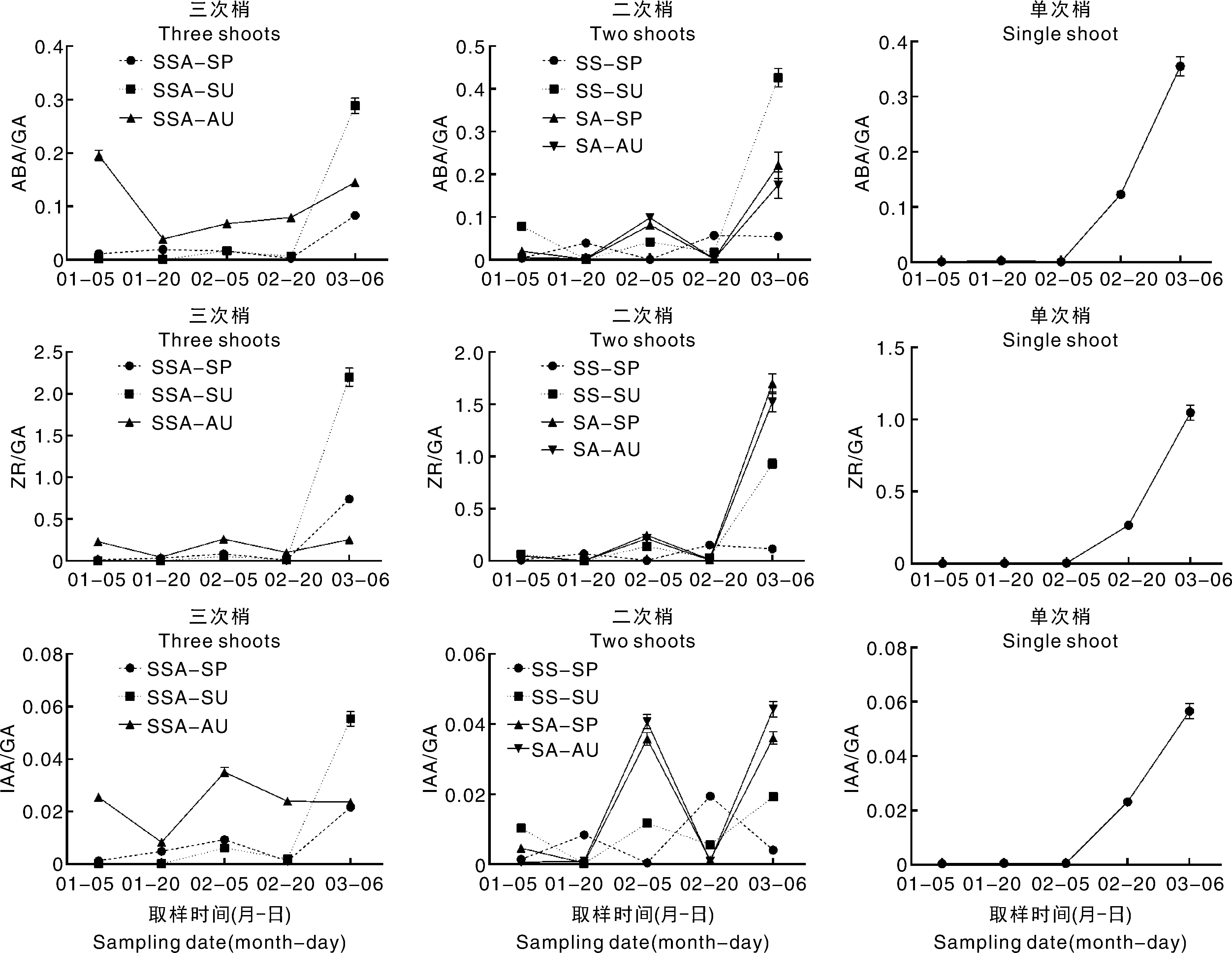
图5 红美人不同结果母枝花芽激素比值比较
2.5 红美人成花过程中叶片碳氮含量变化
由图6可知,红美人叶片C/N大致呈现波动上升的趋势。三次梢中SSA-SP、SSA-SU的C/N在2月20日和3月21日出现两个峰值,随后下降,而SSA-AU的C/N在2月20日达到最低值(19.11),在3月31日出现峰值(28.17)。二次梢中C/N同样呈现波动上升的趋势,且均在3月21日达到峰值,此时SA-AU的C/N高于SA-SP,而SS-SP和SS-SU差异不显著。就单次梢而言,红美人叶片C/N在3月6日出现最低值(18.68),分别在2月5日(24.87)和3月31日(25.73)出现峰值。
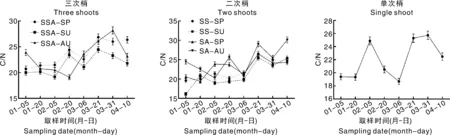
图6 红美人不同结果母枝叶片C/N比较
3 讨论
花芽分化是一个复杂的过程,生理分化完成后,植株受内源激素和外界环境的影响,逐渐分化出花芽和叶芽,随后进入形态分化期,逐渐发育形成花萼、花瓣、雄蕊和雌蕊[3]。本试验中红美人在设施条件下花芽分化过程分为未分化期、分化初期、萼片分化期、花瓣分化期、雄蕊分化期和雌蕊分化期6个时期。
本试验结果表明,红美人花芽形态分化于1月5日开始,3月6日基本完成,各时期均有重叠现象。不同类型结果母枝中,无论是三次梢或二次梢,均为前端梢较中、后端梢花芽分化进程快,且最后处于雌蕊分化期的花芽比例高,解释了前端梢成花能力强,中、后端梢成花能力弱的现象。对各种类型结果母枝花枝率调查结果也显示:三次梢的后端梢(SSA-SP)、中部梢(SSA-SU)和前端梢(SSA-AU)花枝率分别为16.1%、14.3%和57.2%;二次梢(春夏梢)的后端梢(SS-SP)和前端梢(SS-SU)分别为6.7%和37.9%;二次梢(春秋梢)的后端梢(SA-SP)和前端梢(SA-AU)分别为7.7%和45.8%;单次春梢(S-SP)为46.0%,这与花芽分化进程相吻合。基于以上研究结果,可为进一步研究红美人合理的整形修剪方式和最佳的结果枝组配比,减少结果部位外移提供参考依据。
植物内源激素是果树花芽分化的关键诱导因子之一,对花芽分化起重要的调控作用[20]。然而,内源激素在植物体内分布范围广、含量低,且花芽分化不仅受单一激素的影响,还依赖于各种激素的动态平衡,因此多数学者认为不同激素的比值能更真实地反映成花规律。张上隆[21]研究发现,IAA/GA3、ZT/GA3、ABA/GA3上升促进温州蜜柑成花,IAA对成花的效应与黄迪辉等[22]的观点不同。本试验结果表明,红美人在花芽分化进程中,GA含量逐渐下降,ZR含量逐渐上升,ABA含量呈现先降后升的趋势,而IAA的变化没有明显的规律。从激素比值来看,ABA/GA、ZR/GA、IAA/GA均呈上升趋势,且成花率高的枝梢ABA/GA、ZR/GA、IAA/GA高,解释了生产上红美人前端梢成花能力强,中、后端梢成花能力弱的原因。此外,本试验结果表明,花瓣分化期之前,GA含量高,ABA含量、ZR含量、ABA/GA、ZR/GA均处于较低水平;进入雄蕊分化期和雌蕊分化期后,ABA含量、ZR含量、ABA/GA、ZR/GA均快速上升。由此可见,浙北地区设施栽培红美人在雄蕊分化期前后(2月5日到2月20日)是激素调控花芽分化的关键时期。同时,本试验还观察到ABA含量升高多处在花瓣分化期和雄蕊分化期,ZR含量升高多处在雄蕊分化期和雌蕊分化期,即内源ABA与ZR含量的变化不同步,具体原因值得进一步研究。
碳氮物质含量是成花的基础,碳氮营养假说认为,相对碳多氮少的内环境有利于成花。该理论在红肉蜜柚[23]、砂糖橘[24]上也得到了验证。本试验结果表明,在成花过程中,红美人各类型枝梢的叶片均呈现C/N波动上升的趋势,多数成花率高的枝梢C/N较高。二次梢(春夏梢)中春梢和夏梢的C/N虽然差异不显著,但夏梢中的ABA/GA、ZR/GA均高于春梢。由此可见,浙北地区红美人不同类型结果母枝前端梢成花能力强,中、后端梢成花能力弱,这与花芽分化进程、营养物质积累、内源激素水平的差异有关,其中激素起主要调控作用。
4 结论
浙北地区设施红美人花芽分化的过程可分为未分化期、分化初期、萼片分化期、花瓣分化期、雄蕊分化期和雌蕊分化期6个时期;不同类型结果母枝前端梢成花能力强,中、后端梢成花能力弱,这与花芽分化进程、营养物质积累、内源激素水平的差异有关,其中激素起主要调控作用;雄蕊分化期前后(2月5日到2月20日),是激素调控花芽分化的关键时期。红美人杂柑在众多的栽培管理过程中,要特别重视不同类型结果母枝的合理配置,以及夏、秋梢摘心及其放梢时间节点的把握,并尝试利用激素调控花芽分化。
