枸杞子多糖提取物LBP1C通过激活TFEB延缓衰老*
2023-08-14伍东立乔新华韩文生黄雨云飞孙传鑫丁畅1
伍东立 乔新华 韩文生 谢 婷 时 畅 黄雨云飞 孙传鑫丁 侃 陈 畅1,*
(1)西南医科大学基础医学院,泸州 646000;2)中国科学院生物物理研究所生物大分子国家重点实验室,北京 100101;3)中国科学院大学生命科学学院,北京 100049;4)中国科学院上海药物研究所受体研究重点实验室,药物研究国家重点实验室,上海 201203)
枸杞子是中国传统名贵中药,在欧美及亚洲很多国家有“长寿果”之称。《神农本草经》把它列为上品,历代本草均有记载。作为典型的药食同源中药[1-2],枸杞子一直被纳入在国家卫健委发布的食药同源(既是食品又是中药材)的物质目录中。《本草纲目》中记载枸杞子具有“久服坚筋骨,轻身不老,耐寒暑,易颜色,变白”的功效,在“坚筋骨”方面,前期工作已经进行了科学解读和机制研究[3-5],但枸杞抗衰老及美白的科学内涵和机制尚不完全清楚。从枸杞子中已分离出多种具有生物活性的成分,如多糖、类胡萝卜素、类黄酮和酚类等,研究表明枸杞多糖具有抗氧化、抗炎、抗细胞凋亡和细胞保护等作用[6]。研究表明,枸杞粗多糖可以通过提高果蝇的抗氧化能力延长寿命[7]。枸杞粗提物可以延长线虫的寿命并提高线虫对百草枯、紫外线诱导的氧化和热应激的耐受性水平[8]。枸杞粗多糖可以提高D-半乳糖造模的衰老小鼠的认知,降低大脑中脂质过氧化及脂褐质的水平[9],还可以提高皮肤超氧化物歧化酶(SOD)的活性,降低皮肤丙二醛(MDA)含量[10]。在皮肤美白和保护方面,枸杞根提取物能有效抑制细胞内酪氨酸酶活性,降低黑色素含量[11]。皮肤长时间暴露在紫外线辐射下,会导致表皮和真皮的过早老化,紫外线照射诱导丝裂原活化蛋白激酶(MAPK)信号通路,导致胶原降解。枸杞糖复合物(LbGp)具有抗凋亡和抗氧化的作用,其中的一种提取物LbGp5,可以促进人成纤维细胞的活力,提高I 型胶原的含量[12]。枸杞多糖组分(LBPF)可以保护小鼠皮肤由于紫外照射导致的胶原纤维断裂的损伤[13]。枸杞延缓衰老及美白的机制目前大多数研究集中在其抗氧化的功能中,如枸杞可以提高SOD 和过氧化氢酶(CAT)活性,丙二醛水平降低。枸杞抗衰老涉及到的相关信号通路主要为丝裂原活化蛋白激酶MAPK 信号通路的激活[11],其他方面的机制研究得很少。
维持适当自噬水平的机体更加健康和长寿[14],饮食限制和锻炼部分是通过适度激发自噬产生延缓衰老的作用[15]。自噬是真核细胞利用溶酶体清除异常蛋白质和受损细胞器以维持细胞稳态的重要代谢过程,自噬过程中细胞能处理一些对细胞健康有损伤的物质[16]。转录因子EB(TFEB)是自噬-溶酶体途径的重要转录调节因子,它正向调节自噬和溶酶体生物合成相关基因的表达,从而促进自噬体形成、自噬体-溶酶体融合以及自噬降解[17]。研究还发现TFEB通过溶酶体胞吐作用促进细胞内有害底物的清除,此系统功能的下降会导致疾病,年老的帕金森病模型小鼠自噬-溶酶体系统受损导致肾脏中脂褐素颗粒异常堆积[18]。脂褐素是一种老年色素,由蛋白质、脂质和碳水化合物组成,含量随着年龄的增长而增加,并导致细胞功能障碍。脂褐素在皮肤中的过量和异常分布会导致黑斑的出现,是皮肤衰老的一个标志物[19]。衰老过程中神经元最显著的形态学变化之一是脂褐素聚集体以及神经松弛素色素的积累,这些会导致神经元损失、神经胶质增殖和激活[20]。衰老的另一个显著特征是衰老相关分泌表型(SASP),SASP 是一种和衰老相关的促炎反应,包括促炎因子、蛋白酶、促纤维化因子和干扰组织微环境的生长因子,如白介素-1β(IL-1β)、IL-6 和诱导型一氧化氮合酶(inducible nitric oxide synthase,iNOS) 等[21-22]。转录因子GATA4是一个新的衰老调节基因,是SASP所必需的,GATA4可以激活转录因子NF-κB以启动SASP并促进衰老。正常情况下GATA4 会被p62 介导的选择性自噬降解,但这种调节在衰老过程中会受到抑制,因为衰老过程中自噬水平的下降导致p62的积累从而稳定了GATA4。GATA4 在多种组织中积累,包括老化的大脑、皮肤,并可能导致衰老及其相关的炎症[23]。因此,该工作旨在研究枸杞子是否可以通过激活TFEB 提高自噬水平降低衰老的SASP,同时降低细胞内脂褐质,从而延缓衰老及具有潜在美白效应。
我们从枸杞子中获得一种组分多糖LBP1C,在人成纤维细胞及线虫水平对其延缓衰老和清除脂褐质的功能和机制进行了研究。发现LBP1C 可以通过增加TFEB 入核从而提高自噬水平,降低SASP 可以延缓人成纤维细胞的衰老,提高线虫的运动能力,并降低细胞及线虫的脂褐质水平,从而促进健康衰老。本研究揭示了LBP1C 抗衰老和清除脂褐质的功效及其机制,为枸杞抗衰老及产品研发提供了新的科学依据。
1 材料与方法
1.1 LBP1C的制备
取5 kg干燥的枸杞子药材,用粉碎机粉碎。取枸杞粉末(1.0 kg)加入到20 L 去离子水中,于55℃下分别加入30 g 纤维素酶、5 g 木瓜蛋白酶及10 g 淀粉酶,搅拌提取1 h。将溶液温度升高至100℃使酶失活后,4 000 r/min 离心10 min,上清液加热浓缩后流动水透析1~2 d,再加热浓缩透析内液、4 000 r/min 离心10 min。上清液加入5 倍体积的95%工业乙醇并且不断搅拌,静置过夜。4 000 r/min离心10 min并收集沉淀,无水乙醇和丙酮交替洗涤3次,置于50~60℃烘箱中烘干,得酶-水联提枸杞粗多糖(LBP1)。
采用DEAE SepharoseTMFast Flow分离粗多糖。取约6 g LBP1 溶解于60~80 ml 去离子水中,4 000 r/min离心10 min除去不溶物,取上清上样于DEAE Sepharose TM Fast Flow 柱,依次用去离子水和不同离子强度的NaCl 溶液(0.05、0.1、0.2、0.4、0.6、0.8、1.0、2.0 mol/L)进行梯度洗脱,自动收集器收集样品,流速为12 ml/15 min,硫酸-苯酚法检测多糖含量,测定其在490 nm 处的吸光度值(A490),绘制洗脱图,合并相同组分,减压浓缩、透析、冷冻干燥,得到0.05 mol/L 洗脱组分LBP1A1,0.1 mol/L洗脱组分LBP1B,0.2 mol/L洗脱组分LBP1C。LBP1C 为下一步实验用的组分多糖。
细胞处理:用培养液配制干粉LBP1C 终浓度为0.4 g/L 处理约20 代次细胞,处理2 个代次,约6 d;线虫处理:将5 g/LLBP1C 混合进OP50 菌液中喂食线虫,从第1 天(Day1)开始处理,处理10 d。
1.2 人成纤维细胞培养
人成纤维细胞(human dermal fibroblasts,HDF)(美国细胞科学研究中心(ScienCell Research Laboratories),编号:2320)用于细胞衰老的实验研究。细胞培养在含有10% 胎牛血清(fetal bovine serum,FBS)(Gibco,货号:10099-141),加入100 U/ml 青霉素,100 mg/L 链霉素的DMEM 培养基,在5%的CO2,37°C,95%湿度的二氧化碳培养箱内进行培养。
1.3 线虫培养
N2 野生型线虫温度为20℃,湿度为约50%的条件下培养线虫,使用大肠杆菌菌株OP50 喂养线虫。
1.4 蛋白质免疫印迹(Western blot)
用5×SDS-聚丙烯酰胺凝胶电泳(PAGE)上样缓冲液(loading buffer)制备电泳样品后,用8%的SDS-PAGE 胶进行蛋白质分离,然后转移到NC膜。膜用5% 的脱脂奶粉(TBST 配制,tris buffered saline+Tween 20)室温封闭1.5 h(当检测蛋白是带biotin 标签时不用牛奶封闭,直接用TBST封闭)。用相应的一抗在4°C孵育过夜。使用HRP 偶联的二抗标记一抗,用增强型化学发光试剂盒和ChemiDoc XRS+(BioRad)仪器进行显色,显色发光液为超敏发光液(Thermo Scientific,89880)。抗体:Anti-iNOS antibody(Santa Cruz,sc-7271), Anti-LC3B antibody (Cell Signaling Technology,2775S),Anti-P62 antibody (MBL,PM045)。
1.5 实时荧光定量PCR
用SYBR Green法,在PCR反应体系中,加入过量SYBR荧光染料,SYBR荧光染料特异性地掺入DNA 双链后,发射荧光信号,而不掺入链中的SYBR染料分子不会发射任何荧光信号,从而保证荧光信号的增加与PCR 产物的增加完全同步。反应体系见表1。

Table 1 The reaction system for Q-RCR
在ABI7500 上进行定量PCR 检测,引物见表2。对于每个待检测样品,选取3 个独立制备的RNA样本反转录cDNA,并且每个cDNA样品重复检测3次,最后所得的Ct值取平均值,并将目的基因与内参基因做归一化处理后,用2-△△Ct法计算相应基因mRNA 的相对变化。定量PCR 条件:94°C变性10 min;40 个循环;94°C 变性10 s、60°C 退火15 s和72°C延伸20 s,72°C延伸90 s;同时检测产物的溶解曲线。
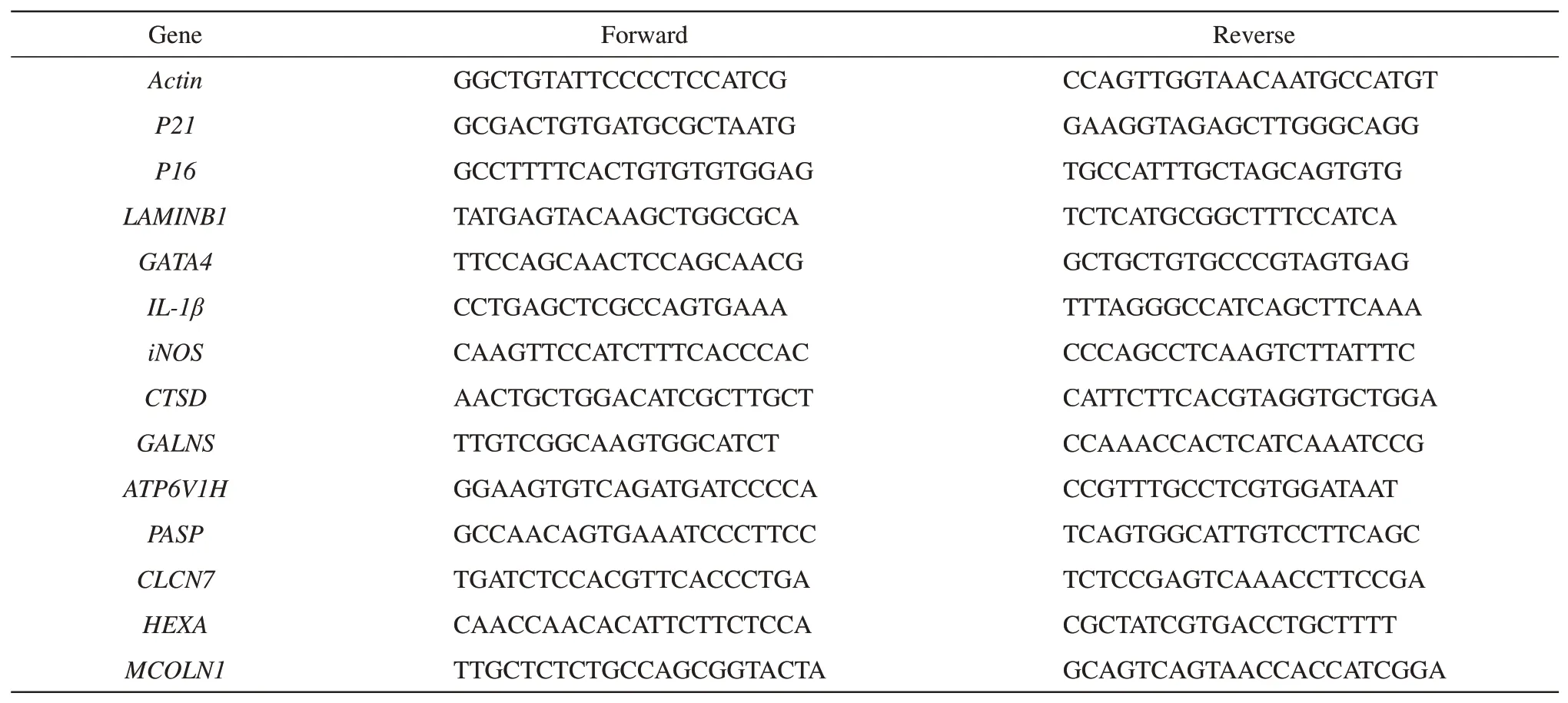
Table 2 Sequences of primers used for the amplification of qPCR
1.6 SA-β-gal染色
吸除细胞培养液,用PBS 或HBSS 洗涤1 次,加入1 ml β半乳糖苷酶(β-gal)染色固定液,室温固定15 min。吸除细胞固定液,用PBS或HBSS洗涤细胞3 次,每次3 min,然后吸除PBS 或HBSS,每孔加入1 ml染色工作液。37ºC孵育12~16 h后用普通光学显微镜下观察。每孔细胞取上、下、左、右、中5个视野,统计被染为蓝色的细胞数,每孔计数>200个。
1.7 脂褐质检测
细胞:苏丹黑染色法,去除细胞培养基,PBS洗1遍,加4%多聚甲醛(PFA),室温固定15 min,PBS 洗3 次,每次2 min。70%乙醇孵育2 min 后用70%乙醇配置0.7%的苏丹黑(现配现用),孵育2~8 min,显微镜下实时观察染色效果,防止沉淀生成。去除苏丹黑,50%乙醇孵育2 min。蒸馏水洗3次,每次2 min,后加入40%甘油,普通显微镜下观察、拍照。
线虫:体内荧光光谱法是检测线虫脂褐质的经典方法[24]。线虫在340 nm激发和430 nm发射下线虫肠道的自发荧光为脂褐质,用高速转盘活细胞荧光成像系统Andor 进行成像,图像荧光用Imaris 软件进行统计分析。
1.8 免疫荧光染色
人成纤维细胞培养于玻璃底皿,去掉培养基后用PBS洗3次。用4%的多聚甲醛室温固定15 min,PBS 洗3 次,每次5 min。细胞用0.5%的Triton X-100(PBS 配)室温通透10 min,用PBS 洗3 次每次5 min。封闭:1% BSA(PBS+0.05% Tween配制)室温封闭1 h 后,一抗(Anti-TFEB antibody(CST,D2O7D)4°C 孵育过夜。用PBST 洗涤3次,每次10 min,去除非特异性结合后,细胞与带有荧光素标记的二抗在室温结合1 h。PBST 洗涤3次,每次10 min,用激光共聚焦显微镜63 倍油镜观察。
1.9 线虫运动检测
在未涂菌的3.5 cm NGM培养基上滴1滴M9缓冲液,挑取一只待检测的线虫放置于M9 的液滴中,让线虫适应30 s,然后在体式显微镜下统计线虫30 s 之内身体弯曲(body bend)的次数;在线虫自然进食条件下对其30 s 之内吞咽次数(pumping)进行统计,每次实验统计15只线虫。
1.10 统计分析
实验所得数据采用SPSS 22.0 统计软件进行统计学分析。计量资料以x±s 表示且均符合正态分布,两组间比较采用两独立样本t检验。
2 结果
2.1 LBP1C延缓成纤维细胞衰老
枸杞成分复杂,各组分可能发挥的功能不同,LBP1C是从枸杞子中提取的组分多糖(图1a)。首选利用干燥成熟的枸杞果实通过酶(纤维素酶、木瓜蛋白酶、淀粉酶)-水联合提取的方法获得枸杞粗多糖(LBP1),枸杞的干燥成熟果实(5.0 kg)经酶-水联合提取,并用5 倍体积的95%工业乙醇沉淀后得到LBP1(129 g,得率为2.58%)。枸杞粗多糖LBP1 采用阴离子交换法(DEAE)以及凝胶过滤色谱法(Sephacryl S-200 HR 及Sephacryl S-300 HR)分离纯化得到组分多糖LBP1C,LBP1经阴离子交换柱梯度洗脱后,得到0.05 mol/L NaCl洗脱组分LBP1A1(11.9 g,得率为9.22 %),0.1 mol/L NaCl 洗脱组分LBP1B(12.0 g,得率为9.30%)以及0.2 mol/L NaCl 洗脱组分LBP1C(13.9 g,得率为10.78%)[25],接下来对其抗衰老及清除脂褐质方面的功能进行研究。复制衰老指体外培养的正常细胞经过有限次数的分裂后,停止生长,细胞形态和生理代谢活动发生显著改变的现象。除了大部分肿瘤细胞和胚胎干细胞,不同年龄阶段不同生物供体来源的细胞均存在复制衰老现象。以来源于人真皮的成纤维细胞复制型衰老模型研究LBP1C 在抗衰老方面的作用。首先对复制型衰老细胞模型的年轻及衰老的细胞进行表征,用SA-β-gal酶检测试剂盒对HDF P10和HDF P25代次细胞进行检测,P25代次的SA-β-gal 阳性细胞数比例显著高于P6 代次(图1b),结果表明P6 细胞为年轻细胞,定义为年轻细胞(early passage,EP),P25代次为衰老细胞(late passage,LP)。用LBP1C 处理HDF P20 代次的细胞,处理2代次(6 d)后进行衰老相关指标的检测,发现与对照组相比,LBP1C 处理组中的衰老标志物P16、P21 的mRNA 表达量显著降低(图1c,d)。LAMIN B1 的丢失是细胞衰老的标志物[26],LBP1C 处理组中LAMIN B1 水平显著高于对照组(图1e)。SA-β- gal阳性细胞数在LBP1C处理组中也显著下降(图1f)。以上结果表明LBP1C延缓了人成纤维细胞的衰老。
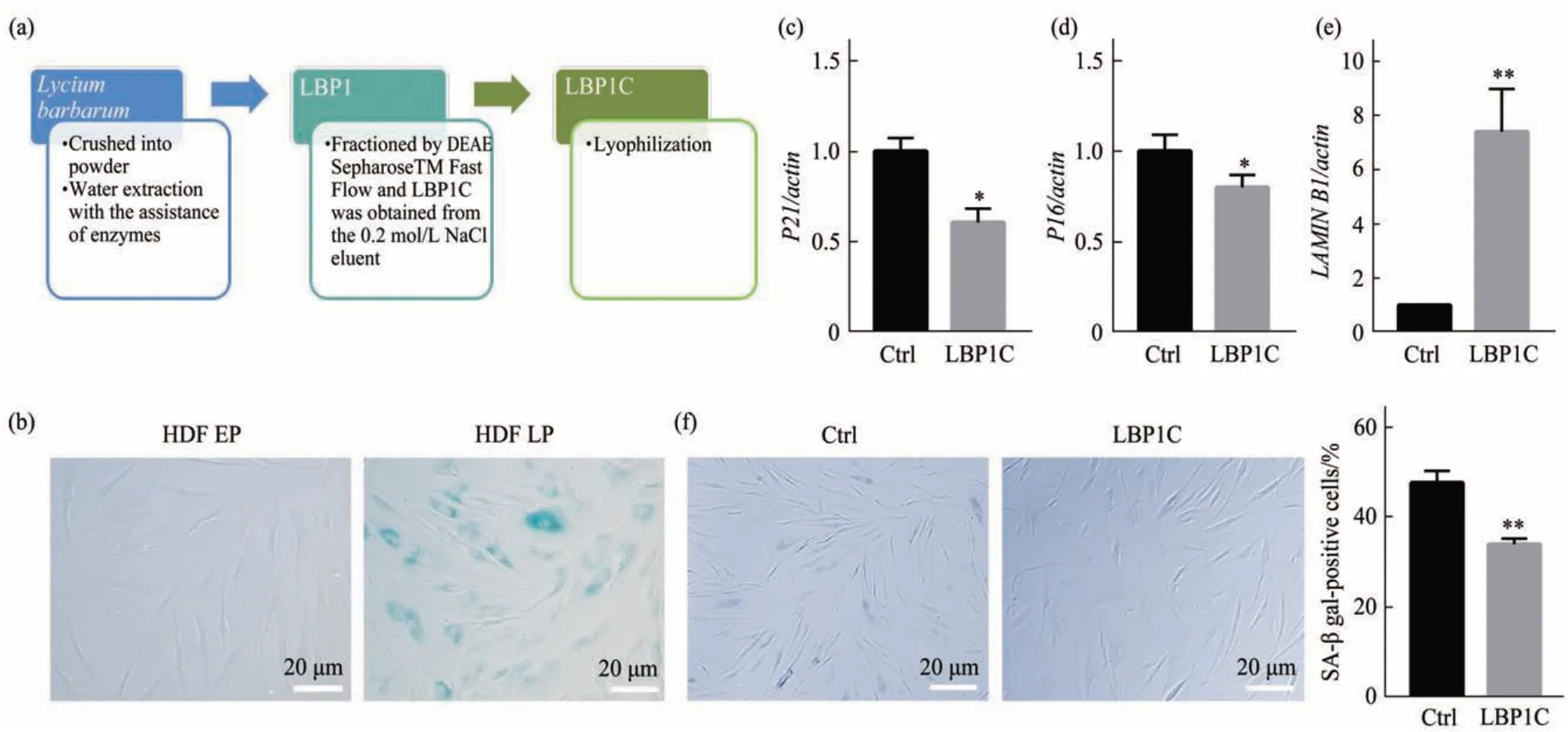
Fig. 1 LBP1C delayed fibroblasts senescence
2.2 LBP1C降低GATA4水平,抑制SASP水平
接下来对LBP1C 延缓细胞衰老的机制进行研究。GATA4 在激活衰老过程中起着关键作用。GATA4 受到自噬的抑制,但当细胞衰老时自噬水平会降低,GATA4 开始积累,其表达诱发了与SASP相关基因的表达[23]。结果发现,相比于对照组,LBP1C 处理组中GATA4 的水平显著下降(图2a),SASP因子IL-1β在LBP1C处理组也显著下降(图2b),iNOS的mRNA和蛋白质水平也都显著降低(图2c,d)。以上结果表明,LBP1C 降低了GATA4水平,抑制了衰老细胞中的SASP。
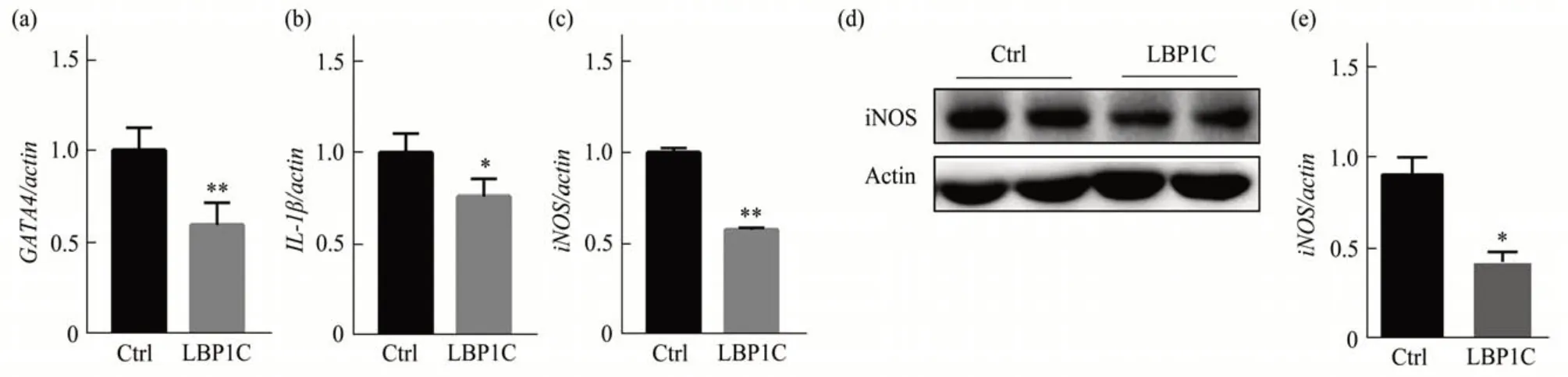
Fig. 2 LBP1C reduced GATA4 level and suppressed SASP
2.3 LBP1C 增加TFEB入核,激活自噬
LBP1C 可以降低GATA4水平,而GATA4受到自噬活性的抑制,因此推测是LBP1C激活了自噬,从而降低了GATA4 的水平。首先检测了溶酶体生物发生和自噬的关键转录调节因子(TFEB)的入核水平。免疫荧光结果表明,相比于对照组,LBP1C 处理组增加了TFEB 入核比例(图3a)。进一步用溶酶体染料检测溶酶体的数量,发现LBP1C 处理组中溶酶体数量显著高于对照组(图3b)。对TFEB下游基因检测表明,溶酶体水解酶、溶酶体膜蛋白及功能相关的囊泡性ATP 酶在LBP1C 处理组都显著上调(图3c),进一步表明LBP1C 促进了自噬的形成。进一步对自噬水平进行检测,发现相比对照组,LC3II/I 的水平显著升高,表明细胞自噬水平显著升高(图3d)。以上结果表明,LBP1C通过增加了TFEB入核从而激活了细胞自噬。
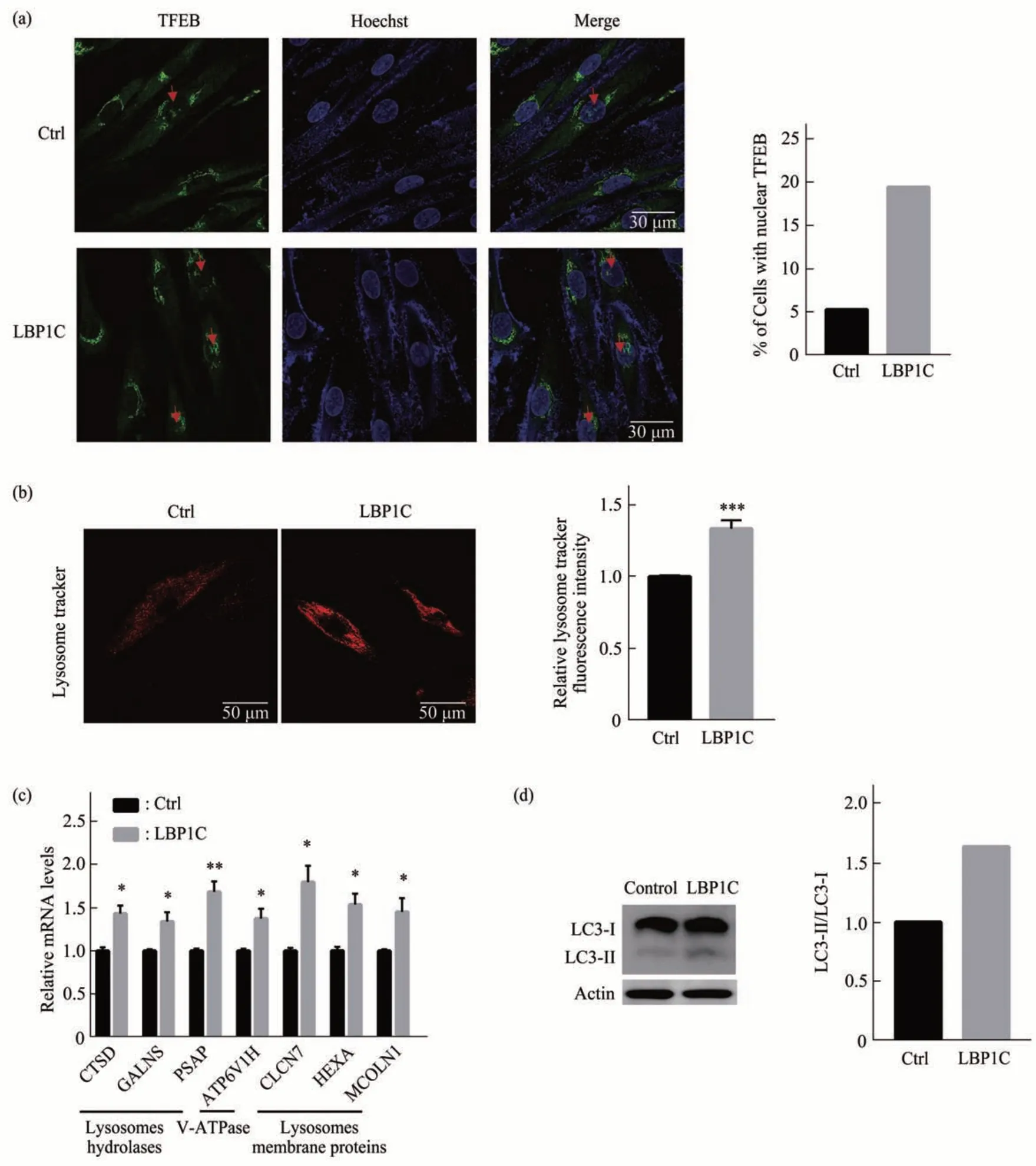
Fig. 3 LBP1C increased TFEB nuclear translocations and activated autophagy
2.4 LBP1C降低衰老细胞及线虫的脂褐质水平
LBP1C 提高了自噬水平,因为自噬可以清除细胞中的聚集物比如脂褐质,所以进一步在细胞及个体水平对LBP1C 如何影响脂褐质的水平进行了研究,发现LBP1C 处理组的细胞中的脂褐质显著低于对照组(图4a)。在线虫水平,将5 g/L 的LBP1C 混合进OP50 菌液中,从线虫Day1 开始进行喂食,对线虫的脂褐质水平及线虫的运动能力进行检测。LBP1C 处理的Day8 线虫中脂褐质水平显著低于对照组(图4b),表明LBP1C可以降低线虫的脂褐质。Day10线虫进入衰老阶段,线虫的身体运动相关指标从Day10 开始出现差异,LBP1C 显著提高了线虫的运动频率和进食速度(图4c,d)。以上结果表明,LBP1C 减缓了由于衰老导致的脂褐质积累及线虫运动能力和进食能力的下降,促进了线虫的健康衰老。
综上所述,本研究表明,LBP1C 通过增加TFEB入核,提高了溶酶体发生,增强了细胞自噬水平。自噬水平的提高一方面使转录因子GATA4降低,从而抑制了衰老相关的SASP表型,另一方面自噬水平的提高降低了衰老过程中产生的脂褐质,因此LBP1C延缓了衰老(图4e)。
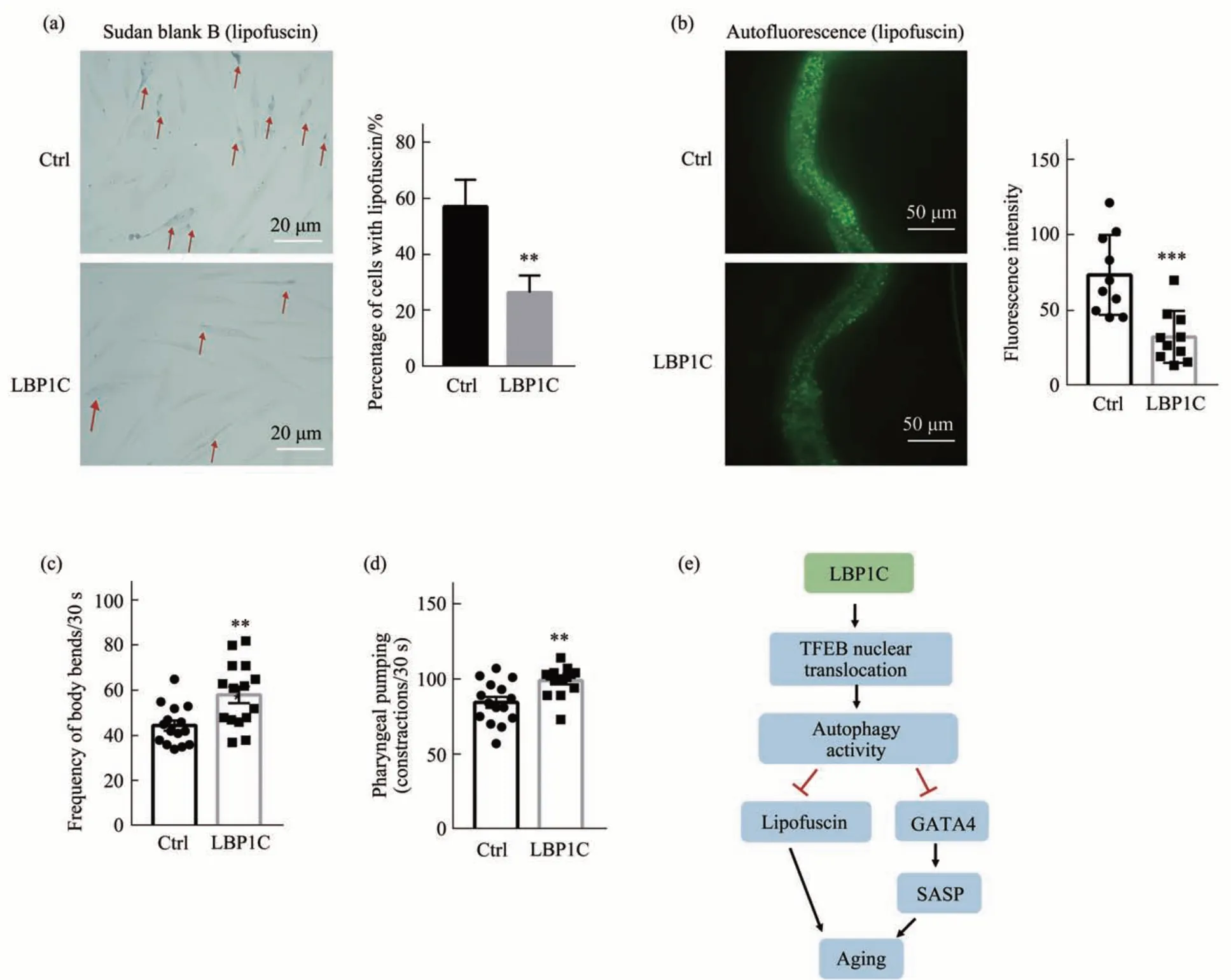
Fig. 4 LBP1C reduced lipofuscin in HDF cells and C. elegans’ and enhanced motility of C. elegans
3 讨论
本研究对从枸杞子中分离得到的一种组分多糖LBP1C 在细胞及线虫体系水平上其抗衰老及潜在美白的效应及机制进行了研究。研究发现,LBPC1降低了自然衰老细胞标志物水平,并降低了SASP,减少了细胞及线虫中脂褐质水平,提高了线虫的运动能力,促进了健康衰老。机制上发现LBPC1 促进了TFEB入核,提高了溶酶体的含量及细胞的自噬水平,从而降低了SASP的调控因子GATA4的水平,进而减少了SASP。同时自噬-溶酶体系统的激活也促使细胞内聚集的底物脂褐质的降解,因此降低了脂褐质的水平。通过本研究发现了枸杞抗衰老和抑制脂褐质的有效成分LBP1C,并揭示了其新的机制,为枸杞的广泛应用提供了科学依据。
3.1 利用自然衰老的模型研究枸杞LBP1C抗衰作用,并发现其促进健康衰老的功效
本研究利用自然衰老的人成纤维细胞模型及自然衰老的线虫模型进行了LBP1C 功效及机制的研究,发现其降低SASP及降低脂褐质的作用。相比以往很多研究是通过加速衰老的小鼠模型[10]或者一些氧化剂处理或刺激进行研究[8],本研究更能模拟枸杞在日常生活中的自然应用,为使用者提供实用的参考信息。抗衰的新宗旨是“健康衰老”,要实现“老而不衰”,即:老龄时也能保持相较于普通水平更高的身体机能水平。之前的研究揭示了锻炼缓解内质网还原应激从而促进健康衰老的机制[27],本研究中发现LBP1C 可以使衰老的线虫的运动能力得到提高,说明枸杞促进了线虫健康衰老,作为药食同源的中药在促进健康衰老方面具有重要的实用价值。
3.2 发现枸杞LBP1C通过TFEB激活自噬延缓衰老新机制
本研究揭示了枸杞子的有效成分LBP1C 发挥功能的新机制:LBP1C激活了TFEB,促进溶酶体的发生及自噬活性从而产生抗衰老及清除脂褐质的效应。研究表明,衰老过程中自噬能力是显著下降的[28],自噬是蛋白质质量控制和维持细胞稳态的重要组成部分,自噬能力下降直接导致SASP的重要转录因子GATA4的积累,从而激活SASP,导致衰老[29]。LBP1C 显著提高自噬水平从而具有抑制衰老的潜能,而且与SASP 相关的IL-1β 及炎症相关的iNOS水平显著下降,相应衰老的标志物P16、P21和β-gal 活性显著下降,线虫由于衰老导致运动能力的下降也得到了挽回;自噬水平的提升也是降低脂褐质的原因之一,该研究为枸杞抗衰的功效理解提供了新的科学证据。以往研究多从其抗氧化的角度出发,清除自由基、抑制脂质氧化从而抑制脂褐质;本研究揭示了枸杞子可以通过激活自噬清除脂褐质前体物质从而减少脂褐质。总之,本文从枸杞子中提取了全新的化合物,并为其抗衰老、潜在美白的功效提供了新的机制。
美白实际包含了两方面内容:美是细胞不老;白是脂褐质等沉积少。本研究从延缓衰老和清除脂褐质沉积两个方面揭示了枸杞子功效成分LBP1C的新作用和新的分子机制,阐释了《本草纲目》记载的枸杞子轻身不老、易颜色、变白的功效的科学内涵。未来将深入研究LBP1C调控TFEB入核的机制,并在哺乳动物中进行验证本研究发现的效应及机制,为枸杞在未来的转化及应用中提供充分的科学依据。
4 结论
本研究使用人类成纤维细胞和线虫作为自然衰老的模型,发现LBP1C 可以降低人成纤维细胞中P16、P21的表达水平以及SA-β-gal 染色阳性细胞的数量,表明LBP1C 可以延缓细胞衰老。进一步研究表明,LBP1C 发挥抗衰老作用是通过降低SASP 和脂褐质水平。机制上,LBP1C 通过促进TFEB 入核增加自噬,并减少细胞和秀丽隐杆线虫中SASP和脂褐素的积累。同时,LBP1C提高了线虫衰老过程中的运动能力,促进健康衰老。本文研究了从枸杞子中提取的LBP1C 的新功能和机制,表明其通过促进自噬来延缓细胞衰老,从而达到抗衰老和皮肤美白的效果。
