昆虫蛀食对卷果涩荠种子萌发特征的影响
2023-08-01文晓虎张玲卫刘会良迪力夏旦木塔什买买提
文晓虎, 张玲卫, 刘会良, 张 岚, 迪力夏旦木·塔什买买提
(1.新疆农业大学资源与环境学院, 乌鲁木齐 830052;2.中国科学院新疆生态与地理研究所荒漠与绿洲生态国家重点实验室, 乌鲁木齐 830011;3.中国科学院大学, 北京 100049; 4.新疆生态与地理研究所伊犁植物园, 新疆 新源 835800)
种子萌发是植物生长发育的起点,关系到植物后期的营养生长和生殖生长,是植物能否成功建植的决定因素[1]。同时种子萌发期也是相对脆弱和敏感的时期,经常受到众多环境因素的影响[2]。如不同的温度条件会影响种子内贮藏物质的转化和能量传递,促进或抑制种子萌发[3];盐分胁迫的毒性作用导致种子萌发率和萌发速率降低,影响植物幼苗的生长发育[4];干旱胁迫致使种子内部失水,损伤细胞器和脂膜,使种子无法正常萌发[5]。在干旱和半干旱地区水分稀少、环境温度高、蒸发量大,影响作物的生长。因此,研究该区域温度和干旱胁迫对种子萌发的影响十分重要。
温度是影响种子萌发率和萌发速率的关键因素[6]。在不同植物种子萌发过程中,存在决定种子萌发的基础温度(Tb)、最适温度(To)和最高温度(Tc)[7],这3个温度决定着植物种子的萌发温度阈值,同时以最适温度为分割点,将植物种子萌发温度阈值分为亚适温和超适温:亚适温指萌发温度处于基础温度和最适温度之间,此时种子萌发速率会随着温度增加呈线性增加;超适温则指处于最适温度和最高温度之间的范围,种子萌发速率呈线性或曲线下降趋势[8]。植物种子萌发所需要的温度条件与种类及其生长所适应的生态和地理环境条件密切相关[9]。由于种子所处的环境以及种子的自身特性不同,对温度的响应也有所区别,其中荒漠植物种子萌发受到的影响更为明显。一般表现为,低温能够打破夏季一年生物种的休眠,而高温可以打破冬季一年生物种的休眠[10]。在打破休眠后荒漠植物种子萌发温度阈值较宽,如四齿芥(Tetracmequadricornis(Steph.) Bunge)种子在15~30 ℃范围萌发率达90%以上[11]。因此,温度对萌发的起始、萌发率和休眠具有显著影响,探究植物种子萌发对温度的响应,有助于了解植物与其生长环境的适应机制。
在干旱与半干旱区域,水分也是调控与影响种子生长发育的关键因素,降水随着季节变化,水资源分布不均匀[12],在一些时期容易造成水分胁迫,对植物种子和幼苗生长发育造成负面影响。干旱胁迫首先降低了种子的吸水速率和最大吸水量,造成种子延迟萌发或者萌发率降低;其次水分供应不足,也会导致种子内部物质代谢降低、酶活性降低等,不仅使种子难以萌发,也致使植物幼苗发育不良[13]。随着水分胁迫强度加剧,植物种子萌发率会持续降低,导致植物种群存活率迅速降低[14]。同时,除水分会对种子萌发造成影响外,来自不同生境的种子在干旱胁迫下表现出明显差异:生长于干旱环境的植物种子则表现出很强的干旱适应性,而从水分较为充足的环境收集的种子对水分胁迫非常敏感[15]。如生长在盐渍化荒漠的短命植物绵果荠(LachnolomalehmanniiBunge)在中低浓度PEG条件下仍能够萌发,高浓度下种子陷入休眠但复水后受抑制的种子可以重新萌发[16]。植物萌发条件是植物长期与其生长环境相适应的特性,因此即使同种植物之间也有明显差异,这种由生长环境不同导致的萌发特征差异可称为母体效应。例如,生长在内蒙古的梭梭(Haloxylonammodendron(C. A. Mey.) Bunge)种子的最适萌发温度为25 ℃;而生长于新疆吐鲁番的梭梭种子最适萌发温度为10 ℃[17]。分别萌发于春季和秋季的短命植物尖喙牻牛儿苗(ErodiumoxyrhinchumM. Bieb.)产生的种子大小不同,在高、低温胁迫下两类种子的响应也存在显著差异[18]。母体效应是由母体植物在种子成熟过程中所经历的条件所决定的,通常在控制成熟种子的萌发性方面起着重要作用。水分和母体效应也是种子休眠启动、中断和发芽的关键驱动因素[19]。
母体效应在萌发特性方面已被证实具有适应价值,植物母体差异和种子形成时环境因素可以调控子代种子萌发能力[20]。昆虫蛀食行为也是影响植物生长的重要生物因素,蛀食行为能够对植物营养生长和繁殖生长造成显著的负面效果,使得遭受蛀食和未遭受蛀食的植物母体间产生显著差异[21]。干旱区昆虫种类丰富,存在大量鞘翅目、鳞翅目和直翅目等昆虫,它们大多数为植食性昆虫,直接以包括短命植物在内的荒漠植物为食[22-24]。短命植物是生长在干旱与半干旱区的一种特殊植物类群,多以种子繁殖,生活周期短,对环境变化具有灵活、多样的适应策略[25]。短命植物在古尔班通古特沙漠中所占比例较大,占植物总数的37.1%,其绿色产量在春季达到60%以上[26],对生态系统和生物多样性的维持与稳定有重要作用[27]。但针对短命植物种子萌发相关的研究多集中在温度、水分和盐碱胁迫等非生物因素造成的影响,而对生物因素如昆虫取食后的母体效应研究较少。那么,昆虫蛀食导致的短命植物母体差异产生的后代种子其萌发特性存在何种差异,以及温度、干旱胁迫是否对后代种子造成影响,是否影响了植物生存及其生态适应性等科学问题还不明确。
以油菜筒喙象(Lixusochraceus(Boheman))蛀食和非蛀食的卷果涩荠(Malcolmiaafricana(Bunge) Boiss.)种子为研究对象,通过对比不同温度和干旱胁迫下两种类型卷果涩荠种子萌发情况,揭示两种类型卷果涩荠种子萌发特性,探究两种类型卷果涩荠种子萌发对温度和干旱胁迫的响应,为干旱区植被恢复、幼苗重建等提供数据支撑和理论基础。
1 材料与方法
1.1 供试物种种子采集与处理
本研究区位于古尔班通古特沙漠的南缘北沙窝(86°57′44″E,44°19′15″N)。于2021年5月至6月,在古尔班通古特沙漠采集卷果涩荠种子。由于卷果涩荠种子成熟后角果极易裂开,使采集种子效率降低,所以在区分油菜筒喙象蛀食/非蛀食的前提下,使用信封直接采集完整的成熟卷果涩荠植株。将装有卷果涩荠植株的信封放置在室温(25 ℃)且通风良好的位置自然晾干,然后使用按压的方法使种子从植株上脱离,在去除较大的杂质后,使用风扇吹种子的方式去除种子中的剩余杂质与不合格种子(瘪粒破损种子),将合格的种子装于信封中在室温下贮存备用。
1.2 种子萌发实验
实验共设置6个恒温条件,分别为5 ℃、10 ℃、15 ℃、20 ℃、25 ℃和30 ℃。采用纸上发芽法,以25粒种子为一组重复,共4组重复,将蛀食/非蛀食组的卷果涩荠种子分别置于直径90 mm铺有两层滤纸的培养皿中,加蒸馏水至滤纸湿润,将培养皿放置在24 h全黑暗环境的恒温室内培养箱(型号:GXZ-288 B,湿度:50%)中进行萌发实验。每日统计种子萌发数,种子胚根突破种皮长度超过种子自身一半时视为发芽。持续28 d,所有重复连续5 d无新增发芽数时结束实验。计算种子萌发率、发芽势与发芽指数。
干旱胁迫处理:目前大量研究选择用高渗溶液模拟干旱表示土壤水分胁迫,其中聚乙二醇(PEG-6000)水溶液是应用最多的高渗溶液调节剂,具有渗透压稳定、无营养、无毒性、重复性好等优点[28-29]。萌发实验设计与上述相同,同样为6个恒温处理(5 ℃、10 ℃、15 ℃、20 ℃、25 ℃和30 ℃),在加蒸馏水的步骤时,将蒸馏水替换为等量的渗透试剂,渗透试剂使用25%浓度的化学纯级的PEG-6000水溶液。以萌发温度实验为ck组,加25%浓度的PEG水溶液的培养皿为对照组。后续统计步骤相同。
发芽率/%=(发芽种子数/供试种子数)×100%;
发芽势/%=(发芽高峰期发芽种子数/供试种子数)×100%;
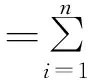
发芽率降幅/%=[(对照培养的发芽率-干旱胁迫的发芽率)/对照培养的发芽率]×100%;
发芽指数降幅与发芽率降幅相同。
1.3 种子发芽温度阈值
种子发芽温度阈值的确定应用SPSS软件中概率单位回归分析法,分别计算不同温度下种子发芽率达20%至90%时所需要的时间(tg),以种子发芽速率(1/tg, g=20%,30%,40%,50%,60%,70%,80%,90%)为纵坐标,发芽温度为横坐标,在亚适温与超适温条件下分别建立发芽速率与发芽温度的线性回归方程,回归直线与X轴交点为种子发芽的温度阈值,两条回归直线的交点所对应的温度为种子发芽所需的最适温度[30]。
1.4 数据分析
使用Excel软件对数据进行初步统计与处理,使用SPSS 19.0 统计分析软件进行相关数据分析,主要包括用单因素方差分析(One-way ANOVA)检验蛀食/非蛀食组卷果涩荠种子发芽率、种子发芽势、发芽速率的显著性差异水平。使用Origin 2017软件绘图。
2 结果与分析
2.1 温度对两种类型卷果涩荠种子发芽的影响
发芽率和发芽势是评定种子发芽能力及种子质量等级的重要指标之一,而发芽指数是衡量种子活力的重要指标。在本实验中,温度对蛀食组和非蛀食组卷果涩芥种子的发芽率均无显著影响,而对种子发芽势和发芽指数有显著影响(p<0.05)。蛀食组中,除15 ℃和25 ℃外,其余温度下发芽率均在90%以上;而非蛀食组的种子发芽率除30 ℃外,均高达90%。表明卷果涩荠种子发芽温度范围较广,两种类型卷果涩荠种子均能够在多种温度下保持较高发芽率。随着温度的增加,蛀食组和非蛀食组的发芽势、发芽指数均呈先升高后降低的趋势(表1)。蛀食组卷果涩荠种子和非蛀食组卷果涩荠种子在5 ℃的低温和30 ℃的高温条件下,虽然最终发芽率较高,但种子的初期发芽比较缓慢,低温和高温都不利于种子的发芽,抑制了种子初期发芽速率。在15~20 ℃培养温度下,蛀食组和非蛀食组种子的发芽势显著高于其余温度处理,在该温度范围内两种类型卷果涩荠种子的发芽能力最高,发芽速率最快。蛀食组和非蛀食组卷果涩荠种子发芽指数均在30 ℃下最低,15 ℃(非蛀食组)和20 ℃(蛀食组)下最高,高温对两种类型卷果涩荠种子发芽活力的影响略大于低温影响,但两者之间无显著差异。

表1 不同温度条件下两种类型卷果涩荠种子发芽率、发芽势和发芽指数Table 1 Germination rate, germination potential and germination index of seeds of Malcolmia africana (Bunge)Boiss. of two types under different temperature conditions
对卷果涩芥种子的发芽速率与发芽温度进行线性回归,结果表明,种子发芽温度阈值在蛀食组卷果涩芥和非蛀食组卷果涩芥中存在差异(图1、表2)。其中,蛀食组的Tb和To都大于非蛀食组,而非蛀食组的Tc大于蛀食组(表2),筒喙象甲幼虫的蛀食行为降低卷果涩荠种子发芽的温度阈值。说明非蛀食组种子在较低温度条件下即开始积累积温值,但同时非蛀食组卷果涩荠种子发芽所需要的积温也大于蛀食组卷果涩荠种子,需要更长的发芽时间积累积温以达到发芽。两种类型卷果涩荠种子的这种发芽Tb较低的植物所需积温较高,Tb较高的植物所需积温较低的情况,避免了种子在多变的温度环境下较早发芽所遇到的风险。

表2 卷果涩荠种子发芽的温度阈值和回归方程Table 2 Temperature thresholds and regression equations for seed germination of Malcolmia africana (Bunge)Boiss.
蛀食组卷果涩芥和非蛀食组卷果涩芥种子发芽的最低、最高和最适温度的变幅范围均较小,如蛀食组种子发芽的Tb、Tc、To变幅范围分别为1.19~1.21 ℃,30.78~31 ℃和19.69~19.75 ℃(图1左);非蛀食组发芽的Tb、Tc、To变幅范围分别为0.7~0.71 ℃,34.96~35.65 ℃和15.42~15.55 ℃(图1右)。由图1可知,蛀食组种子和非蛀食组种子发芽速率分别在20 ℃和15 ℃最高,由此确定两种类型卷果涩荠种子发芽的亚适温与超适温温度范围。0~20 ℃为蛀食组卷果涩荠种子发芽的亚适温范围,0~15 ℃为非蛀食组卷果涩荠种子发芽的亚适温范围,在这一温度范围内种子发芽速率随着环境温度的升高而加快,与发芽温度呈显著正相关;一旦超过种子发芽的最适温度,如蛀食组在25~30 ℃超适温条件下,非蛀食组在20~30 ℃超适温条件下种子发芽速率开始急剧降低,发芽速率与发芽温度呈显著负相关,由此可见环境温度进一步升高将不利于种子的萌发。
2.2 干旱胁迫对两种类型卷果涩荠种子发芽的影响
经过PEG模拟的干旱胁迫后,两组卷果涩荠种子发芽率对干旱胁迫的响应具有显著差异(p<0.05)(表3)。相比对照组,遭受干旱胁迫的非蛀食组卷果涩荠种子发芽率均表现为显著降低,降低幅度为16.4%~33.1%,其中在发芽温度为10 ℃、15 ℃、20 ℃时降低幅度最大,均超过25%。表明非蛀食组卷果涩荠种子发芽率更易受到干旱胁迫的影响,在低温或高温条件影响下,干旱胁迫对种子发芽率的影响较低。而蛀食组卷果涩荠种子的表现与非蛀食组卷果涩荠则完全相反,仅在5 ℃和30 ℃下发芽率显著降低,降低幅度分别为15.8%和13.5%;在其余发芽温度下,蛀食组卷果涩荠种子发芽率减低幅度均小于等于7.7%。表明蛀食组卷果涩荠对干旱胁迫响应并不明显,在干旱胁迫下仍能够保持较高发芽率,但在低温与高温条件影响下,发芽率会出现大幅下降,极端温度对蛀食组卷果涩荠种子发芽率影响较大。
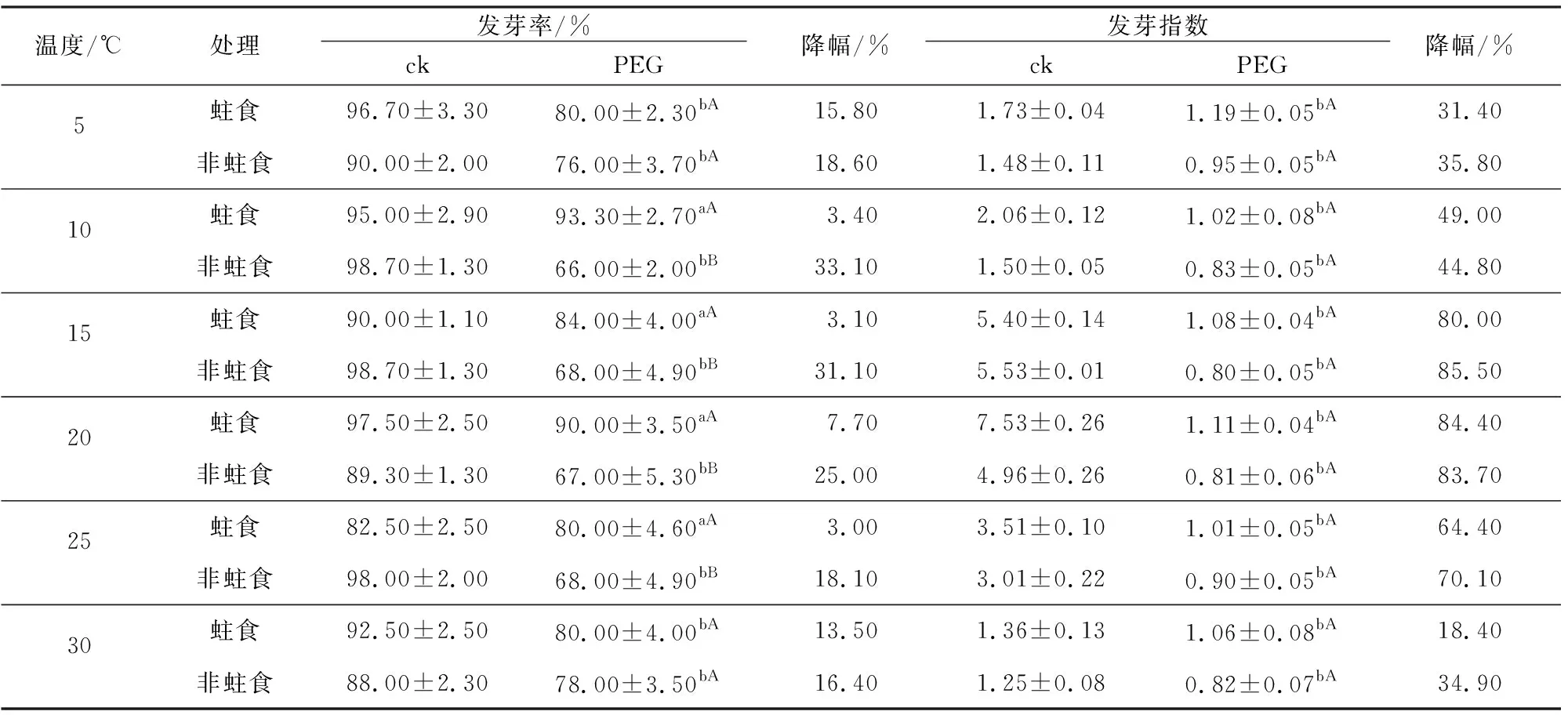
表3 PEG干旱胁迫下两种类型卷果涩荠种子发芽率和发芽指数Table 3 Seed germination rate and germination index of Malcolmia africana (Bunge)Boiss.of two types under PEG drought stress
在同温度条件下,蛀食组/非蛀食组卷果涩荠种子发芽率之间存在差异。除5 ℃和30 ℃的发芽温度下,表现为蛀食组卷果涩荠种子发芽率高于非蛀食组,差异不显著外;其余4个发芽温度下,均表现为蛀食组卷果涩荠种子发芽率显著高于非蛀食组。表明蛀食组卷果涩荠种子对干旱胁迫的抗性要强于非蛀食组。进一步对两种类型卷果涩荠种子在干旱胁迫下的发芽指数进行比较,发现干旱胁迫对两种类型卷果涩荠种子的发芽指数影响趋势相同,相比对照组发芽指数均显著降低,降低幅度在18.4%~85.5%之间,在蛀食组/非蛀食组卷果涩荠种子组内6个温度下的发芽指数均无显著差异,表明干旱胁迫显著降低了两种类型卷果涩荠的种子活力,此时发芽指数受到干旱胁迫的显著影响,而温度对活力造成的影响相对较小,差异不显著。
蛀食组卷果涩荠种子和非蛀食组卷果涩荠种子组间的发芽指数在6个温度处理下均无显著差异,但蛀食组卷果涩荠种子的发芽指数均大于同温度下的非蛀食组卷果涩荠种子发芽指数,其原因是蛀食组卷果涩荠种子发芽指数本身大于非蛀食组卷果涩荠种子发芽指数,因此出现干旱胁迫后两种类型卷果涩荠种子发芽指数差异不显著。进一步说明在水分充足的情况下,适宜的温度会提高种子发芽指数,增强种子活力;但在水分不足干旱胁迫下,水分是限制卷果涩荠种子发芽指数的首要因素。
3 讨 论
3.1 温度对卷果涩荠种子发芽特性的影响
温度对种子发芽率的影响表现为低温和高温会抑制种子发芽,不同植物受到抑制的温度也存在差异。本研究结果显示,温度对两种类型卷果涩荠种子发芽率无显著影响,各温度下种子最终发芽率均在85%以上。古尔班通古特沙漠属于冷荒漠,气温变化幅度极大,有研究指出古尔班通古特沙漠3月至7月的温度变化幅度为-22.7 ℃~30.3 ℃[31]。在古尔班通古特沙漠中调查显示,包括卷果涩荠在内的多种短命植物种子都集中在3—4月大量发芽,但在后期雨水充足的条件下,在4至6月期间也存在部分种子发芽情况,甚至在高温的夏季出现发芽[32]。因此,在水分充足的情况下,两种类型卷果涩荠种子在多个温度条件下仍然保持较高发芽率是与古尔班通古特沙漠特有的环境相匹配,同时也表明卷果涩荠种子存在相对较广的发芽温度阈值。
发芽温度是植物种子完成后续生长发育的重要环境条件,植物种子在特定的温度范围内发芽,表明植物所处的温度生态位[33]。因此,发芽温度是影响植物分布与丰度的重要筛选条件。在本实验中,蛀食组卷果涩荠产生的种子发芽温度阈值相对较小,所处温度生态位更狭窄,更容易受到环境温度变化的影响[34];而非蛀食组卷果涩荠拥有更宽的发芽温度阈值,但也需要更高的发芽积温值,这代表非蛀食组卷果涩荠可以在更广泛的温度条件下发芽,但它需要比蛀食组卷果涩荠更多的发芽时间。发芽速率的线性回归结果显示,0~20 ℃为蛀食组卷果涩荠种子发芽的亚适温范围,而非蛀食组卷果涩荠种子亚适温范围则是0~15 ℃。这个结果则恰好解释了蛀食组卷果涩荠种子和非蛀食组卷果涩荠种子在20 ℃时发芽率、发芽势、发芽指数出现显著差异的原因,即20 ℃温度已经处于非蛀食组卷果涩荠种子发芽的超适温范围,此时种子发芽与温度呈负相关关系。因此,非蛀食组卷果涩荠发芽受到抑制,各项发芽特征均显著低于蛀食组;蛀食组则受到温度促进能够更快地发芽,更多占据生态位,拥有更强的竞争能力与同时期更多短命植物进行竞争。
种子发芽实验的结果表明,卷果涩荠种子发芽势和发芽指数整体变化相同,随着发芽温度升高,发芽势与发芽指数呈先增加后降低的趋势,蛀食组卷果涩荠在20 ℃下种子的发芽势和发芽指数最高,此时种子活力和质量等级达到最佳,非蛀食组卷果涩荠则是在15 ℃下最佳,超过15 ℃后,种子发芽势与发芽指数开始逐步下降。结果与表2预测结果高度吻合(蛀食组最适温度19.72 ℃,非蛀食组最适温度15.43 ℃)。两种类型卷果涩荠种子的最适发芽温度出现差异,一方面是因为蛀食组卷果涩荠种子由于本身发芽温度阈值较小,选择在晚春时期(20 ℃)大量发芽;非蛀食组卷果涩荠广泛的发芽温度阈值使其于早春时节(15 ℃)大量发芽,在之后降低自身发芽速率从而达成与蛀食组卷果涩荠种子错峰发芽的结果。这种结果避免了种子同期发芽造成同生境内生态位过度拥挤导致的种内竞争。Quarles and Roach[35]的研究中也提到,在生境资源有限的条件下,植物会通过降低个体植株生长速率或者增加单株植物的结实量来降低种内竞争的压力,提高植物获取资源量,增加植物的适合度。另一方面,蛀食组卷果涩荠种子错峰推迟发芽可能与自身防御机制有关。油菜筒喙象在3月初就开始交配产卵行为[36]。蛀食组卷果涩荠推迟发芽的行为降低被早期油菜筒喙象产卵蛀食的风险。类似的现象也发生在同为十字花科的黑芥菜(Brassicanigra)上,在遭受油菜粉虱(Pierisbrassicae)产卵侵扰后,黑芥菜加速繁殖投资,使得种子提前成熟,避免了油菜粉虱幼虫对其花的采食[37]。这种推迟发芽时间躲避植食性动物捕食的逃逸方式也是植物防止自身适合度降低的一种手段。
3.2 干旱胁迫对卷果涩荠种子发芽特性的影响
水分是限制种子发芽和幼苗生长的重要环境因素。本研究结果显示,在25%的PEG溶液胁迫影响下,蛀食组和非蛀食组两种类型卷果涩荠种子的发芽均受到不同程度负面影响,但两种类型卷果涩荠种子的发芽率仍然高于60%,表现出较强的耐旱性。卷果涩荠种子具有较强耐旱性与其母体生长环境长期受到干旱胁迫影响有关。植物母体在环境胁迫下会通过表观遗传的途径将自身应对机制传递给后代,使后代在遭受与母体相同胁迫时可以更快速地进行响应,具有更强的抵抗力[38]。同时植物母体受到干扰时间越久会更容易将这种表达传递下去,母体效应表达会更明显[39]。非蛀食组卷果涩荠种子发芽率受到干旱胁迫的影响程度要显著大于蛀食组卷果涩荠,在6个温度处理下,仅在5 ℃和30 ℃下发芽率与蛀食组卷果涩荠无显著差异。本研究认为,油菜筒喙象对卷果涩荠母体植株的蛀食产卵行为增加了后代卷果涩荠种子对环境的抵抗能力。植物遭受植食性昆虫采食后会改变物理性状,通过物理防御的方式增强对植食性昆虫的抵抗力,如表皮硬化会增加植食性动物采食难度,叶面积减小会降低植食性昆虫的产卵选择性,使昆虫繁殖受到影响[40]。同时叶面积减小、韧皮增厚硬化等物理特性的改变,也减少植物体内水分丧失,提高植物的抗旱能力[41]。推测卷果涩荠母体遭受油菜筒喙象的蛀食胁迫后,改变了自身物理性状,增强对昆虫防御性能同时,也促使自身抗旱性增强。因此,通过母体效应产生了具有更强抗旱性的种子。
干旱胁迫改变了两种类型卷果涩荠种子的发芽方式,使卷果涩荠种子陷入休眠,发芽时间大幅推迟,在16 d后才开始陆续发芽。造成两种类型卷果涩荠种子的发芽指数大幅度降低,种子发芽速率和活力也降低,但干旱胁迫下不同温度间的发芽指数无显著差异。说明水分条件是限制卷果涩荠种子发芽的首要因素。
4 结 论
通过对遭受油菜筒喙象蛀食和非蛀食的卷果涩荠产生的种子在不同温度梯度和干旱胁迫下的发芽特性进行研究,得出以下结论:
1) 发芽温度对两种类型卷果涩荠种子发芽率无显著影响,但发芽温度过低和过高均会抑制卷果涩荠的发芽速率。油菜筒喙象蛀食缩小了卷果涩荠发芽温度阈值,但使卷果涩荠拥有更高的亚适温范围且加快了种子发芽速率。
2) 干旱胁迫使两种类型卷果涩荠种子均进入休眠,推迟种子发芽时间,在温度胁迫和水分胁迫同时干扰卷果涩荠种子发芽时,水分胁迫是影响卷果涩荠种子发芽的首要因素。油菜筒喙象蛀食增强了卷果涩荠种子抗旱性,蛀食组卷果涩荠种子发芽率显著高于非蛀食组卷果涩荠。
