刺梨种子的顽拗性和休眠特性
2023-05-30樊卫国官纪元周禹佳
樊卫国 官纪元 周禹佳

摘 要:探究刺梨(Rosa roxburghii Tratt.)种子的顽拗性和休眠特性,为刺梨种子的保存和解析刺梨在自然条件下的种群繁衍、扩散提供科学依据。采用在3 ℃和-3 ℃低温下保湿贮藏后定时播种的方法,以不进行低温保湿贮藏为对照,分别测定不同失水率的和未失水的刺梨种子在不同低温下保湿贮藏80 d后的萌芽率,并对在3 ℃下保湿贮藏不同时间的未失水刺梨种子的发芽率、发芽势进行测定,同时测定未失水种子在3 ℃下保湿贮藏过程中的赤霉素(GA1+3)、玉米素核苷(ZR)、生长素(IAA)和脱落酸(ABA)的含量变化。结果表明:刺梨种子不耐失水,也不耐0 ℃以下的低温,对失水和0 ℃以下的低温反应敏感。失水或在-3 ℃下贮藏会丧失刺梨种子的生命力。在3 ℃条件下对未失水的刺梨种子进行低温保湿贮藏,能够保持种子的生命力并解除种子的休眠,在此过程中,刺梨种子中的ABA含量不断降低,GA1+3、ZR和IAA含量不断升高,GA1+3、ZR和IAA与ABA的含量比值不断增大。未失水的刺梨种子发芽率与种子中的GA1+3、ZR和IAA含量呈极显著正相关,与ABA的含量呈极显著负相关,与GA1+3、ZR、IAA与ABA的含量比值呈极显著的正相关。刺梨种子具有顽拗性和生理休眠特性。未失水的刺梨种子在3 ℃下保湿贮藏35 d,生理休眠才开始逐渐解除,完全解除休眠至少需要低温保湿贮藏80 d以上时间,即3 ℃的低温累积量至少要达到1 920 h。种子的顽拗性限制了野生刺梨种群的繁衍和扩散。
关键词:刺梨;种子;顽拗性;休眠特性;低温保湿贮藏;发芽率;发芽势
中图分类号:S661.9
文献标志码:A
种子是显花植物世代繁衍、种群扩散和更新的主要载体[1]。多数植物主要是依靠种子进行种质的传播,并提高和维持植物种群的遗传多样性,扩大种群的分布[2]。刺梨(Rosa roxburghii Tratt.)是原产我国西南地区的特有树种,在贵州中、高海拔地区的野生分布最为集中,周边省、市(区)分布的量较少。有研究者认为,鸟、兽的觅食是野生刺梨远距离传播基因交流的主要方式[3],但这一觀点的科学依据不足,迄今,究竟是什么因素影响着野生刺梨种质的扩散及资源分布尚不清楚。研究刺梨种子的繁殖生物学特性,对于解析上述问题和刺梨种子的保存、繁殖都有重要的科学意义。
植物种子可分为正常型、顽拗型和中间型三类[4-5]。正常型种子在成熟过程中水分逐渐减少,其代谢活动也随之降低,干燥或低温贮藏后这类种子仍能发芽[6]。顽拗型种子对脱水的反应极为敏感,既不耐失水也不耐低温,这类种子在成熟过程中始终保持很高的含水量,失水和低温都会损伤细胞膜或原生质,导致种子的生命力丧失[7-9]。中间型种子对脱水和低温的敏感性介于正常型和顽拗型种子之间,但含水量降至一定程度后或贮存于0 ℃以下也会丧失生命力[10-11]。定期测定脱水后的种子含水量和萌发率,了解种子脱水与萌发的关系,成为很多研究者确定种子是否具有顽拗性的重要方法[12]。
很多植物的种子具有休眠特性,这是植物应对不利环境的一种重要适应策略。多数正常型种子没有休眠特性,如玉米、水稻、小麦等农作物种子,但有的仍然会休眠,如苹果、梨、桃、李等的种子,顽拗型种子也是如此。对于有休眠特性的顽拗型种子,导致其休眠的原因各异,有的在脱离母体时,种胚组织尚未发育完全,需要经历胚的后熟过程才能萌发,有的因致密的种皮难以透水透气而抑制萌发,有的则是处于一种生理抑制状态,需要解除生理休眠后才能萌发[1]。内源激素对种子的生理休眠和萌发具有重要的调控作用,其中,脱落酸(ABA)能够使种子保持生理休眠,而赤霉素(GA)则能促进种子的萌发[13-14]。
在野生刺梨调查中发现,即便有大量的成熟刺梨果实落于地面,也很难发现地面有刺梨苗的生长,在自然条件下是何因素影响刺梨的自然繁衍有待进一步探究。刺梨种子不易保存,采用常规方法保存后发芽率极低[15]。为了探究刺梨种子的生物学特性,以便为解析刺梨的自然繁衍及种质传播和种子保存提供依据,本研究采用不同低温下保湿贮藏后定时播种的方法,研究了不同失水率、不同低温保湿贮藏时长对刺梨种子萌发的影响,分析了未失水刺梨种子在3 ℃下保湿贮藏过程中不同内源激素含量的变化,探究了刺梨种子成活及萌发与种子含水量及内源激素变化的关系,旨在确定刺梨种子的顽拗性和休眠特性,为解析野生刺梨自然繁衍、种质扩散方式和种子的贮存提供科学依据。
1 材料与方法
1.1 材料
试验于2018—2020年在贵州大学//国家林草局刺梨工程技术研究中心进行。供试材料为 ‘贵农5号刺梨成熟果实中的种子。种子贮藏所用的细砂为粒径2~5 mm的洗净河砂,播种基质为洗净、消毒后的2 mm粒径的石英砂和蛭石,混合比例1∶1。测定内源激素含量的酶联免疫试剂盒由中国农业大学提供。
1.2 试验设计
1.2.1 试验设计1
为了解种子不同失水率对其萌发的影响,以未失水种子为对照(0CK),设置种子失水率为10%、20%、30%、40%、50%、60%、70%、80%、90%、100%的10个处理。试验分常温对照组、3 ℃低温处理组和-3 ℃低温处理组,常温对照组是在种子的失水率达到设定值时,立即在适宜种子萌发的温度下播种。两个低温处理组是在种子的失水率达到预定值时,将不同失水率的种子分别在3 ℃和-3 ℃下保湿贮藏80 d后播种。不同处理播种的种子数为100粒,重复3次。
1.2.2 试验设计2
为了探究未失水种子低温保湿贮藏时长对其萌发影响,将未失水种子在3 ℃下保湿贮藏至114 d,在此期间,每隔7 d取种子播种测定萌发率,即以低温保湿贮藏每持续7 d为1个处理,每次播种量100粒,重复3次。
1.2.3 试验设计3
为了解未失水种子低温保湿贮藏时长对其內源激素含量的影响,将未失水种子在3 ℃下保湿贮藏至114 d,其间,每隔7 d取种子测定其中的不同内源激素含量,即以低温保湿贮藏每持续7 d为1个处理,每个间隔期均进行3次重复取样测定。
1.3 试验及测定内容与方法
1.3.1 种子的采集
在‘贵农5号刺梨果实成熟期采收果实取出种子,清洗,用浮选法去掉坏种和瘪种,然后滤干种子的表面水后备用。
1.3.2 种子的含水量及失水率的测定
新鲜种子的含水量用烘干称重法测定。用自然失重法测定种子的失水率,即以新鲜未失水种子的含水量视为100%,将未失水种子分成若干份分别称重,然后分别摊放在通风的室内使其自然失水,用减重法测定种子的失水率,将达到试验设计失水率的种子进行下一步试验处理。
1.3.3 种子的低温保湿贮藏处理
将试验种子与细河砂按1∶1的比例混合,河砂的湿度为潮润程度,然后分别置于3 ℃和-3 ℃恒温冰箱中进行不同低温保湿贮藏处理,对贮藏在3 ℃下的种子每隔3 d翻动1次。
1.3.4 播种及种子萌发观察和发芽率、发芽势的统计
按照试验设计的时间要求在智能人工气候室内进行播种。种子播于黑色塑料槽基质中。智能人工气候室的空气湿度控制在85%±1.5%,白天气温控制在28 ℃±0.5 ℃,夜间气温为20 ℃±0.5 ℃,白天7:00至19:00的光照强度为1 100 μmol·m-2·s-1。对播种基质适时喷雾保持湿润。种子播种后每隔7 d观察记录1次发芽的种子数,直至种子不再发芽时结束观察记录,按以下公式计算种子的发芽率和发芽势:
发芽率=(发芽种子粒数/处理的种子粒数)×100%
发芽势=(规定观察时间内发芽种子粒数/供试种子粒数)×100%
1.3.5 种子中内源激素含量的测定
对在3 ℃下保湿贮藏的未失水种子,每隔7 d称取3份,每份0.500 0 g,立即用液氮冷冻后,加2 mL 80%的冷甲醇在冰浴下研磨成匀浆,置于-79 ℃超低温冰箱中保存,待所有测定样品全部取样处理完毕后,用酶联免疫法[14]测定种子中的赤霉素(GA1+3)、玉米素核苷(ZR)、生长素(IAA)和脱落酸(ABA)的含量。
1.4 数据处理分析
用Excel进行数据处理并作图,用DPS v7.05软件进行方差分析,多重比较采用新复极差法。
2 结果与分析
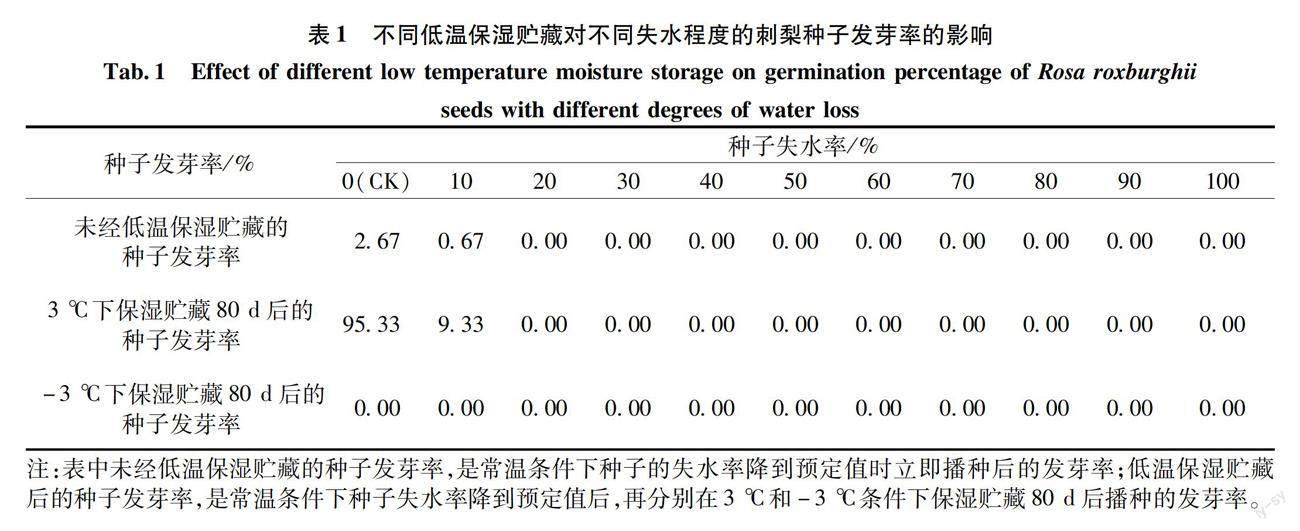
2.1 刺梨种子的含水量和不同低温保湿贮藏对不同失水率的刺梨种子萌发的影响
测定结果表明:新鲜刺梨种子的含水量为24.65%。在未经低温保湿贮藏的条件下,未失水的刺梨种子发芽率仅有2.67%,当失水率增加到种子含水量的10%时,发芽率仅有0.67%,失水率超过种子含水量的10%后,刺梨种子的发芽率为0(表1)。未失水的刺梨种子在3 ℃下保湿贮藏80 d后播种,发芽率高达95.33%,而失水率达10%的种子,发芽率降低至9.33%,失水率大于10%的种子的发芽率都为0。将不同失水率的种子在-3 ℃下贮藏80 d后播种,所有处理的发芽率都为0。
以上试验结果说明,刺梨种子既不耐失水,也不耐-3 ℃的低温,种子稍微失水后就会丧失生命力。在3 ℃下低温保湿贮藏后,能够促进未失水刺梨种子的萌发,但若失水率超过种子含水量的10%,即便通过3 ℃的低温保湿贮藏,种子也无生命力。由此可见,刺梨种子对脱水和0 ℃以下的低温反应敏感,具有明显的顽拗性特征。未失水刺梨种子在3 ℃下保湿贮藏80 d后能够大量萌发,与种子的休眠被解除有关。
2.2 低温保湿贮藏的时间长短对未失水刺梨种子萌发的影响
在3 ℃下对未失水刺梨种子进行保湿贮藏,能够促进种子萌发,但必须达到足够的低温保湿贮藏时间,否则种子不能萌发。表2显示,低温保湿贮藏不足35 d的刺梨种子发芽率为0,贮藏时长到35 d时,刺梨种子的发芽率只有9.33%,此后,随低温保湿贮藏时间的延长,种子发芽率不断提高,低温保湿贮藏84 d的种子发芽率达到96.01%,继续延长贮藏时间,发芽率略有提高。这一试验结果说明:未失水的刺梨种子需要低温保湿贮藏达到35 d后,种子的休眠才开始逐渐解除,要90%以上的种子解除休眠,至少需要近80 d的低温保湿贮藏时间,即3 ℃的低温累积量至少要达到1 920 h。
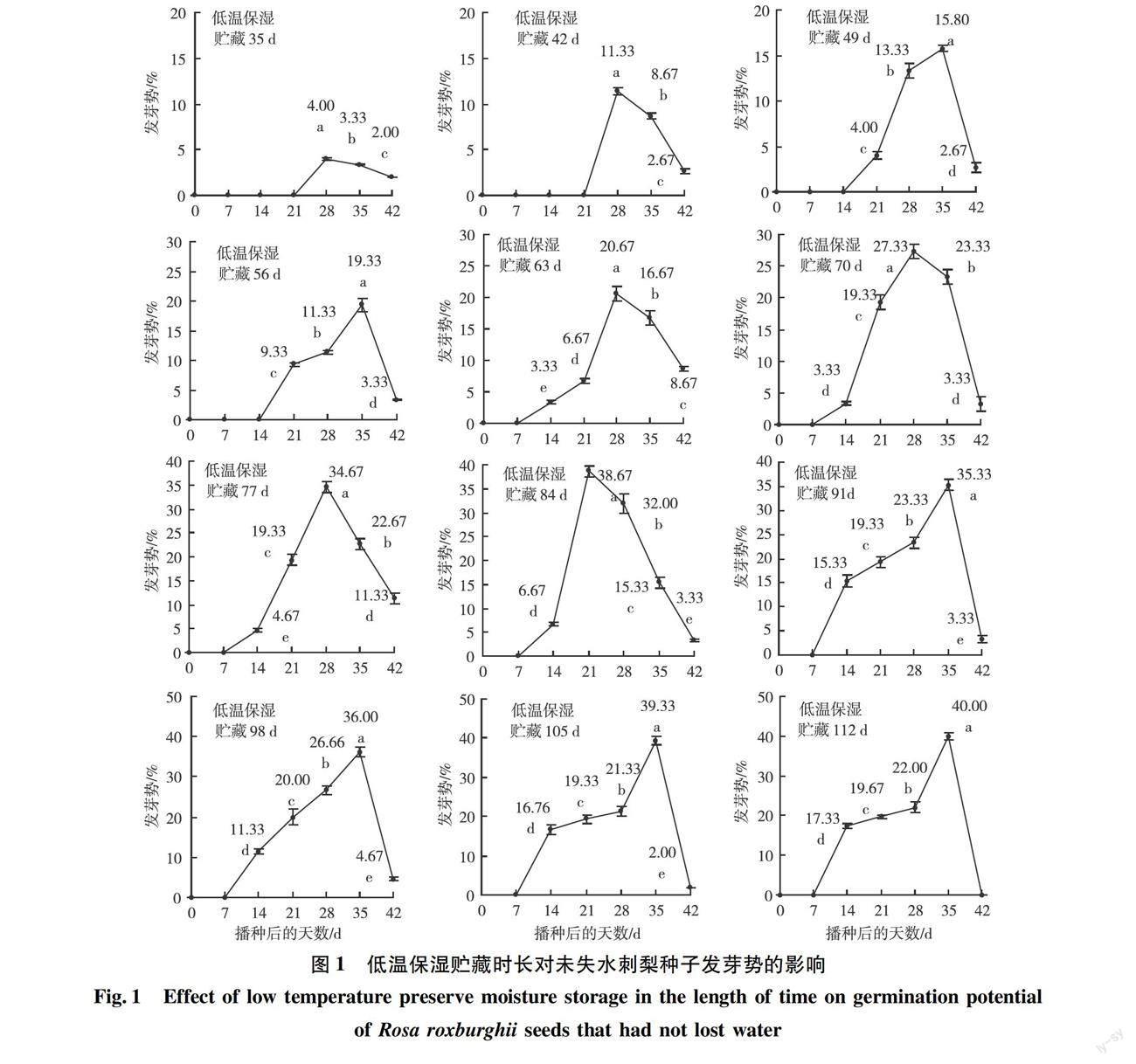
2.3 低温保湿贮藏的时长对未失水刺梨种子发芽势的影响
种子的发芽势是指种子在某个时间段内的发芽率,是衡量种子质量和发芽能力的重要指标,种子的发芽势高,说明种子在某段时间内萌发的数量多,幼苗的整齐度好。试验结果表明,低温保湿贮藏35 d和42 d的种子,播种后4周才发芽,初始发芽势分别仅为4.0%和11.33%(图1),低温保湿贮藏49 d和56 d的种子,播种后3周发芽,初始发芽势分别为4.0%和9.33%,第4周的发芽势分别增强至11.33%和13.33%。低温保湿贮藏63 d以上的种子,播种7 d后就开始发芽,并随低温保湿贮藏的时间延长,初始发芽势呈明显增强的趋势,从3.33%增加至17.33%。从图1还看出,低温保湿贮藏时间少于56 d的种子,发芽的时间明显推迟,发芽势明显减弱,发芽率也明显降低,而低温保湿贮藏时间达到63 d以上的种子,播种后的发芽时间能够提早1周,播种后第4周的发芽势都能增强至20%以上。低温保湿贮藏84 d的种子,播种后第3、第4周的发芽势分别从第2周的6.67%增强至38.67%和32.0%,这两周的发芽势合计达70.67%,说明大量的种子是在第3~4周萌发的,而低温保湿贮藏91 d的种子,播种后第5周发芽势为35.33%。低温保湿贮藏98 d、105 d和112 d的种子,第5周发芽势分别为36.0%、39.33%、40.0%。以上结果说明,刺梨种子发芽需要足够的低温时间打破休眠。在试验中还观察到,无论低温保湿贮藏时间多长的种子,播种42 d后均不再发芽。
2.4 未失水刺梨种子在低温保湿贮藏过程中的内源激素含量变化
2.4.1 不同内源激素的含量变化
在低温保湿贮藏过程中,未失水的刺梨种子中赤霉素(GA1+3)、玉米素核苷(ZR)、生长素(IAA)和脱落酸(ABA)的含量变化均有规律性。测定结果表明:在刚脱离母株的刺梨种子中,抑制发芽的ABA的含量高达2 330.79 ng·g-1FW,而在此时,种子中与萌发相关的GA1+3、ZR、IAA的含量都很低,分别仅为20.61 ng·g-1FW、0.15 ng·g-1FW和7.87 ng·g-1FW。但随低温保湿贮藏时间的延长,种子中的ABA含量迅速降低,而GA1+3、ZR和IAA的含量持续增加(图2),低温保湿贮藏84 d后,种子中ABA的含量降低开始维持在低水平和变化不大的状态,而在同期之后,种子中的GA1+3、ZR和IAA的含量仍然在明显增加,一直要到98 d后,其含量才趋于相对稳定。未失水的刺梨种子在低温保湿贮藏过程中的上述内源激素含量变化规律,与种子休眠的解除有关。
2.4.2 GA1+3、ZR、IAA与ABA的含量比值变化
GA和ABA是具有生理拮抗功能的两类内源激素,其中,种子中的GA对其萌发生理有正调作用,ABA可将GA的作用抵消[14,16]。通常,在GA与ABA的含量比值增大到一定程度后,种子的萌发生理开始得以启动[13]。表3显示,随低温保湿贮藏时间的延长,未失水刺梨种子中GA1+3与ABA的含量比值在不断地增大,而且从贮藏开始到91 d期间,每隔7 d的大多数比值间均有显著差异(P<0.05),贮藏到84 d后,种子中GA1+3与ABA的含量比值已经大于1,第91 d达到了1.063 0,到112 d增加到1.231 0,在贮藏到第98 d后的3周内GA1+3与ABA的含量比值的差异已经不显著。种子中ZR、IAA与ABA的含量比值变化也有相似的规律,但有所不同的是,在整个低温保湿贮藏过程中ZR、IAA与ABA的含量比值始终小于1,其最大值仅分别为0.260 1和0.331 3。
2.5 刺梨种子发芽率与内源激素含量的相关性
2.5.1 与GA1+3、ZR、IAA和ABA含量的相关性
相关性测定结果表明(图3):刺梨种子的发芽率与种子中GA1+3、ZR和IAA的含量呈极显著正相关,相关系数分别为0.91**、0.96**和0.93**,而种子发芽率与种子中的ABA呈极显著负相关,相关系数为0.93**。这种相关性反映出刺梨种子中的不同内源激素含量水平对种子的萌发与休眠具有重要调控作用,随GA1+3、ZR、IAA含量的增加和ABA含量的降低,梨种子的萌发率随之提高,反之,种子萌发率降低。
2.5.2 与GA1+3、ZR、IAA与ABA含量比值的相关性
刺梨种子的发芽率与种子中GA1+3、ZR、IAA与ABA的含量比值间也表现出极显著的正相关,其相关系数分别为0.88**、0.87**和0.86**,说明刺梨种子中GA1+3、ZR、IAA与ABA的含量比例对萌发也有至关重要的影响,其含量比值越大对促进种子萌发的调控作用越明显。
3 讨论
3.1 刺梨种子对失水和低温的敏感性
种子对脱水的敏感性是判定种子有无顽拗性的重要依据[1,12]。正常型种子耐失水、耐低温,顽拗型种子极不耐失水且对低温敏感[1,7,17-18]。在本研究中发现,刺梨种子对脱水和0 ℃以下的低温反应敏感,既不耐失水,也不耐0 ℃以下的低温,失水率达到新鲜种子含水量的10%以上就不再萌发,在-3 ℃持续低温下即便是短期的贮藏,未失水或失水的种子都会失去生命力,而在3 ℃低温下保湿贮藏后,能促进未失水种子的萌发,但即便是同样贮藏条件,种子一旦失水,生命力也会丧失。由此可见,刺梨种子具有顽拗性,属于顽拗型种子,不能干燥保存或在0 ℃以下保存。
种子顽拗性是植物长期适应特定生态环境的结果[1]。分布于气候环境潮湿、季相变化较小的植物,种子的水分不易散失,在长期进化过程中逐渐形成了种子不耐脱水的生态习性[1,12,19]。野生刺梨大多集中分布在雨量充沛、气候湿润的地区,具有喜湿、耐涝性极强、抗旱性极弱的生态特性[20-22],其种子的顽拗性与刺梨长期适应上述特殊的生态环境有关。
3.2 刺梨种子的休眠特性
虽然大多数顽拗性种子不会休眠,但也有一些顽拗性种子具有休眠特性,这类有休眠特性的种子需要在低温下打破休眠才能萌发[12],而且不能失水。脱落酸(ABA)在种子休眠中具有重要的调控作用,是种子休眠诱导和维持的正调控因子,而赤霉素(GA)具有促进种子萌发和拮抗 ABA 的作用[23]。未失水的刺梨种子在3 ℃的条件下保湿贮藏后,种子中的ABA含量随贮藏时间的延长而不断降低,GA1+3的含量不断增加,种子的萌发率也随之提高,这是刺梨种子具有休眠特性的重要生理学证据。低温能抑制休眠种子中ABA合成的基因表达,促进ABA的降解,而GA有类似低温的作用[13,24-25]。很多研究发现,决定种子究竟是休眠还是萌发,不仅取决于种子中的ABA含量的多少,更重要的還取决于GA与ABA的相互平衡,在高水平的GA和低水平的ABA状况下,种子的生理行为趋于萌发,反之,种子保持休眠[13,16,24,26]。在本研究中,随低温保湿贮藏时间的延长,刺梨种子中GA1+3、ZR和IAA含量不断增加,ABA含量不断降低,GA1+3、ZR、IAA与ABA的含量比值也不断增大,这是种子休眠逐渐被解除的重要生理表征。虽然ZR和IAA对种子休眠的解除没有直接联系,但其含量增加对促进种胚的细胞分裂和分化肯定有重要作用。
一些研究者将种子的休眠分为物理休眠、形态休眠、形态生理休眠、生理休眠和复合休眠(物理休眠+生理休眠)五大类型[1,27],其中,物理休眠是由坚硬、致密的种皮结构对种子萌发形成的障碍,形态休眠是种胚未充分发育而使种子不能萌发,生理休眠是因种子中存在抑制萌发生理过程的物质所致,这种抑制物质主要是ABA[16,24,26]。刺梨种子虽然有坚硬、光滑的木质化种皮,但并不是种子萌发的物理障碍。将未进行解除生理休眠处理的刺梨种子用物理和化学的方法破损种皮后,发芽率仍然很低[15]。本研究结果证明,刺梨种子的休眠应属于生理休眠。
3.3 刺梨種子的顽拗性和休眠特性对种群繁衍、扩散的影响
种子繁殖是种子植物种群繁衍、扩散和自然更新的主要方式之一,这种方式对提高并维持种群的遗传多样性和增强植物对异质环境的适应性都具有重要作用。种子的顽拗性对植物生态位的拓展造成了一定限制[1],在自然状态下,很多正常型种子容易在土壤中形成永久的种子库,而对短命的顽拗型种子则不能[19]。刺梨种子的顽拗性和休眠特性对其种群的繁衍、扩散具有不利的影响。刺梨的果实一般在8—9月成熟,脱离母株的刺梨种子还未等到冬季低温来临前,处于休眠的种子会因短期干旱失水而丧失生命力。据长期的观察,鸟、兽不觅食野生刺梨的果实,这可能与刺梨果实上有刺,果肉味酸涩,粗纤维多,不会软化有关。因此,认为野生刺梨种群扩散和基因交流主要是通过鸟、兽的觅食将种子远距离传播实现的,这一观点不符情理,即便如此,刺梨种子不耐失水的顽拗性也不利于种子远距离传播后萌发。在对野生刺梨的调查中发现,虽然刺梨果实中种子很多,但树下难见小苗,这种现象与刺梨种子的顽拗性、休眠特性和8—9月后常出现的短期干旱有关。由此可见,刺梨种子的顽拗性限制了种群的繁衍与扩散。野生刺梨的种群繁衍与扩散可能与土壤芽库有关,刺梨的根容易产生不定芽,地质变化、泥石流等外力作用会将带有刺梨根段的土壤芽库“搬运”出原地,进而在异地形成植株,但这种方式难以实现远距离的种质扩散和传播,这有可能是野生刺梨大多集中分布在某些区域的重要原因。
4 结论
刺梨种子具有顽拗性和休眠特性,种子不耐失水,也不耐0 ℃以下的低温,失水或在0 ℃以下贮藏会导致种子的生命力丧失。未失水刺梨种子在3 ℃下保湿贮藏,能够保持种子的生命力并逐渐解除种子的休眠,在此过程中,刺梨种子中的ABA含量不断降低,GA1+3、ZR和IAA含量不断升高,GA1+3、ZR和IAA与ABA的含量比值不断增大。低温保湿贮藏35 d后,刺梨种子的休眠才开始逐渐解除,随贮藏时间的延长,种子发芽率不断增加,发芽势也不断增强。刺梨种子的休眠属于生理休眠,完全解除休眠至少需要80 d以上的低温保湿贮藏时间,即3 ℃的低温累积量至少要达到1 920 h。刺梨种子的顽拗性限制了种群的繁衍与扩散。
参考文献:
[1]张俊杰, 蒋学皎, 郭庭鸿,等.顽拗性种子脱水敏感性研究进展[J]. 西北林学院学报, 2021, 36(3):144-151.
[2] 王桔红, 陈文. 5种马鞭草科植物种子萌发对低温层积和干燥贮藏的响应及其更新对策[J].生态学杂志, 2015, 34( 12): 3313-3318.
[3] 张怀山, 鄢秀芹, 鲁敏, 等. 基于EST-SSR标记的贵州野生刺梨居群遗传多样性分析[J]. 中国农业科学, 2017, 50(6): 1098-1108.
[4] ROBERTS E H. Predicting the storage life of seeds[J]. Seed Science and Technology,1973,1: 499-514.
[5] NTULI T M, BERJAKP P, PAMMENTER N. Different assessments of the effect of drying rates on recalcitrant seed material[J]. American Journal Biollogy and Life Sciences,2015,3(3):75-79.
[6] MATILLA A J, MAMATILLA-VAZQUEZ M A. Involvement of ethylene in seed physiology[J]. Plant Science, 2008,175(1):87-97.
[7] FARRANT J M,WALTERS C. Ultrastructural and biophysical changes in developing embryos of Aesculus hippocastanum in relation to the acquisition of tolerance to drying[J]. Physiology Plant, 1998, 104(4): 513-524.
[8] BERJAK P, PAMMENTER N W. Seed recalcitrance-current perspectives[J]. South African Journal of Botany, 2001, 67(2): 79-89.
[9] MARQUES A, NIJVEEN H, SOMI C, et al. Induction of desiccation tolerance in desiccation sensitive Citrus limon seeds[J]. Journal of Integrative Plant Biology, 2019, 61(5): 624-638.
[10]DIAS D S, LOPES P S N, RIBEIRO L M, et al. Tolerance of desiccation and cryopreservation of Butia capitata palm seeds[J]. Seed Science and Technology, 2015, 43(1): 90-100.
[11]PAMMENTER N W, BERJAK P. Physiology of desiccation-sensitive(recalcitrant)seeds and implications for cryopreservation[J]. International Journal of Plant Sciences,2014,175(1):21-28.
[12]闫兴富, 杜茜, 周立彪. 顽拗性种子的脱水敏感性及萌发生态学研究进展[J]. 种子, 2007, 26(7): 44-48.
[13]樊卫国, 马文涛, 龙登楷, 等. 保湿冷藏对湖南山核桃种子萌发及生理的影响[J]. 果树学报, 2011, 28(5): 831-836.
[14]李宗霆, 周夑. 植物激素及其免疫检测技术[M]. 南京: 江苏科学技术出版社, 1996: 250-298.
[15]向显衡. 刺梨的繁殖技术研究[J]. 贵州农学院丛刊, 1984, 3(1): 23-34.
[16]江玲, 万建民. 植物激素ABA和GA调控种子休眠和萌发的研究进展[J]. 江苏农业学报, 2007, 23(4): 360-365.
[17]杨海平, 咸洋, 韩彪, 等. 林木顽拗性种子研究进展[J]. 山东林业科技, 2021(5): 88-92.
[18]潘丽梅, 付金娥, 万凌云, 等. 植物顽拗性种子保存及脱水、低温敏感性研究进展[J]. 江苏农业科学, 2021, 49(4): 1-7.
[19]杨期和, 尹小娟, 叶万辉, 等. 顽拗型种子的生物学特性及种子顽拗性的进化[J]. 生态学杂志, 2006, 25(1): 79-86.
[20]樊卫国, 刘国琴, 何嵩涛, 等. 刺梨对土壤干旱胁迫的生理响应[J]. 中国农业科学, 2002, 35(10): 1243-1248.
[21]樊卫国, 安华明, 刘国琴, 等. 刺梨的生物学特性与栽培技术[J]. 秣业科技开发, 2004, 18(4): 45-48.
[22]潘佳佳. 刺梨的耐涝性及其对水涝胁迫的响应[D]. 贵阳: 贵州大学, 2018.
[23]宋松泉, 刘军, 徐恒恒, 等. 脱落酸代谢与信号传递及其调控种子休眠与萌发的分子机制[J]. 中国农业科学, 2020, 53(5): 857-873.
[24]曾骧. 果树生理学[M]. 北京: 北京农业大学出版社, 1992: 1-39.
[25]于敏, 徐恒, 张华, 等. 植物激素在种子休眠与萌发中的调控机制[J]. 植物生理学报, 2016, 52(5): 599-606.
[26]王艳梅, 姚兵, 刘伟伟, 等. 休眠解除后山桐子种子萌发过程中内源激素的动态变化[J]. 林业科学, 2018, 54(6): 44-52.
[27]唐安军. 中国特有植物凹叶木兰种子的休眠与贮藏行为[J]. 植物生理学报, 2014, 50(1): 105-110.
(责任编辑:曾 晶)
Recalcitrance and Dormancy Characteristics
of Rosa roxburghii Seeds
FAN Weiguo*1, GUAN Jiyuan2, ZHOU Yujia1
(1.Engineering Technology Research Centre for Rosa roxburghii of National Forestry and Grassland Administration, Guizhou University, Guiyang 550025, China; 2.Guizhou Botanical Garden, Guiyang 550004, China)
Abstract:
Exploring the recalcitrance and dormancy characteristics of seeds of Rosa roxburghii Tratt., this research was aimed to provide a scientific basis for the preservation of Rosa roxburghii seeds and the analysis of the populations breeding and diffusion of Rosa roxburghii under natural conditions. The method of regular sowing seeds after moisture storage at the low temperature of 3 ℃ and -3 ℃ was used, while the seeds without low temperature moisture storage was used as control. The germination percentage of Rosa roxburghii seeds with different percentages of water loss and those that had not lost water were respectively determined after 80 days of moisture storage under different low temperatures. The germination percentage and germination potential of Rosa roxburghii seeds that had not lost water were determined after moisture storage under 3 ℃ for different time period. At the same time, the contents change of gibberellic acid (GA1+3), zeatin riboside (ZR), indole-3-acetic acid (IAA) and abscisic acid (ABA) in seeds that had not lost water were determined during moisture storage at 3 ℃. The results showed that Rosa roxburghii seeds were sensitive, and not tolerant to water loss and low temperature below 0 ℃, both of which would result in vitality loss of Rosa roxburghii seeds. The Rosa roxburghii seeds that had not lost water after moisture storage under the low temperature of 3 ℃ could maintain the vitality of seeds and break the dormancy of seeds. During the process of low temperature moisture storage, the ABA content in Rosa roxburghii seeds decreased continuously, the contents of GA1+3, ZR and IAA increased continuously, and the content ratio of GA1+3, ZR and IAA to ABA increased continuously. The germination percentage of Rosa roxburghii seeds that had not lost water were extremely positively correlated with the contents of GA1+3, ZR and IAA in seeds, as well as with the content ratio of GA1+3, ZR and IAA to ABA in seeds, while extremely negatively correlated with the contents of ABA in seeds. The seeds of Rosa roxburghii were proved to have the recalcitrance and physiological dormancy characteristics. The physiological dormancy of Rosa roxburghii seeds that had not lost water was broken gradually after 35 days of moisture storage under 3 ℃. The breaking dormancy needed at least 80 days of moisture storage under low temperature, or the low temperature moisture storage under 3 ℃ should reach at least 1 920 h. The recalcitrance of the seeds limited breeding and diffusion of wild Rosa roxburghii populations.
Key words:
Rosa roxburghii Tratt.; seeds; recalcitrance; dormancy characteristics; low temperature moisture storage; germination percentage; germination potential
樊卫国,1958年生,贵州安龙人,贵州大学首批二级教授,博导和学科(学术)带头人,国务院特殊津贴专家,贵州省第二批核心专家 。长期从事刺梨、柑橘等果树的育种及生理生态学研究和产业化技术研发。现任国家林草局刺梨工程技术研究中心(贵州大学)主任,贵州省刺梨学会理事长,贵州省园艺学会理事长,中国园艺学会理事,中国柑橘学会理事,贵州省林木良种审定委员会经济林专业委员会主任,国家十二五、十三五、十四五全国高等农林院校教材——《果树栽培学各论(南方本)》副主编。先后主持国家和省、部级科研项目50余项,育成刺梨优良新品种4个,脐橙品种新4个,核桃新品种1个,并率先创建了我国刺梨良种繁育及栽培生理生态学和产业技术体系、喀斯特石漠化地区柑橘生理生态学及优质高产技术体系,共制订刺梨、柑橘等果树的省级地方标准26个并获得批准发布实施,育成的刺梨、脐橙、核桃等新品种及配套技术累计在我国南方省、区、市推广300余万亩,产生了重大经济、生态和社会效益。先后共获省、部级科技成果奖11项,其中有9项为第一获奖人,发表研究论文160余篇,出版学术专著4部,国家级教材1部。
收稿日期:2022-08-21
基金项目:国家林草局刺梨工程技术研究中心运行项目( 2019133002)
作者簡介:樊卫国(1958—),男,教授,博士生导师,研究方向:果树育种及生理生态与栽培,E-mail:wgfan@gzu.edu.cn.
通讯作者:樊卫国,E-mail:wgfan@gzu.edu.cn.