6种一年生草本物种响应于发芽季节的生物量分配机制
2023-05-22尹任娅
尹任娅,王 姝
(贵州大学 林学院,贵州 贵阳 550025)
种子发芽作为生活史的起点,是植物生命周期中最重要的阶段之一,决定着植物幼苗和成熟阶段的表现和种群延续[1].在水分适宜的环境条件下,某些一年生杂草的发芽过程能够持续整个生长季.不同的发芽季节使植物经历不同环境因子变化的综合影响,从而改变其在整个生长季的生长过程及最终表现,使同一物种形成表型差异巨大的一系列植物种群[2-3].研究植物应对不同发芽季节的生长策略对于理解其在自然环境条件下的种群生存与延续不仅有重要的生态学意义,还可能涉及很多进化学问题.在多变的环境中,植物进化出了感应季节性环境信号的机制,以使发芽、生长和繁殖的时间与有利条件相吻合[4].例如,有研究表明较高的土壤温度刺激了AbutilontheophrastiL.,sidaspirzosaL.和sesbaniaexaltataL.的发芽[5],对于这些物种来说,在4月底或5月初的幼苗发芽与土壤温度的上升有关[6],但随后的幼苗发芽则与降雨模式有关[7].
通常情况下,在季节早期发芽的幼苗比晚期发芽的幼苗表现得更好,因为早发芽的植株可能有更高的存活率、更好的生长状况和适应性[8-9].由于生长期缩短,延迟发芽的幼苗可能根系较浅、植株矮小、叶片少,以及碳水化合物储备不足,无法在后续的干旱环境中生存,所以幼苗死亡率偏高[10].并且,发芽时间的短暂延迟可能导致植物的生物量和繁殖特征发生巨大变化[11-12].例如,延迟萌发5 d会导致地三叶(TrifoliumsubterraneumL.)总生物量减少约50%[13];随着发芽时间的延迟,狗尾草(Setariaviridis)的种子产量显著降低[14].另外,虽然小万寿菊(Tagetesmicrantha)早期发芽的幼苗比后期发芽的幼苗存活率要低,但早期存活下来的个体比后期发芽的个体有更高的繁殖力[15].这可能由于早发芽与发芽推迟的植物采用不同的生长和繁殖策略:早发芽的植株繁殖延迟,营养生长和生命周期延长,表现出竞争策略(C-strategy);而晚发芽的植株更倾向于杂草策略(R-strategy),即由于生命周期缩短,其具有更短的营养生长时间并提前开始繁殖[16].
即便如此,与发芽推迟的植物相比,在季节早期发芽的植物并不总能保持优势,甚至可能有劣势[17].因为环境压力往往在季节早期出现,例如干旱、洪涝、风暴和早期霜冻等环境灾害事件[18-19],可能会使植物面临更高的死亡风险[11].当季节早期环境条件较差的时候,延迟发芽或许对一年生草本植物更有利.所以,发芽时间在决定幼苗存活和生长方面的相对重要性可能在很大程度上取决于发芽时间不同的植物所经历的后期环境因素相对有利还是不利[20-21].因此,在不同时间发芽对于植物来说可能各有不同的利弊,而延迟发芽的生态学意义可能被低估了[19].有研究表明,一种自然常见杂草西部苋(Amaranthusrudis)延迟发芽的群体相比早发芽的群体开花较快、花期相对较短、资源分配效率更高且种子生产潜力很大[22].但也有相关研究表明,植物有最佳发芽时间段.例如,苘麻(Abutilontheophrasti)的最佳发芽季节在晚春(6月末),在该时期发芽的植物可以在相对有利的条件下最大限度地发挥其生长潜力[23].因此,在生长季内发芽的不同植物可能都存在最佳发芽时间段,也可能不同物种的最佳发芽时间段不一致.然而,由于相关研究十分欠缺,植物在季节时间这一问题上的研究仍存在很多争议.
植物可以通过形态上或生理上的调整响应于不同的环境条件,即表型可塑性[24-25].生物量分配是生活史特征的核心概念[26],研究生物量分配的可塑性可能对于理解植物适应不同发芽时间的生长策略有重要意义.已有研究大多集中于发芽季节对生活史特征或繁殖特征的影响[10-11,27-29],缺少对植物在营养生长阶段的可塑性的相关研究.有限研究表明,发芽季节推迟会导致苘麻(Abutilontheophrasti)叶片生物量分配和繁殖生物量分配增大,茎生物量分配降低,根生物量分配可能降低或不变[23].说明晚发芽的植物营养期的缩短主要源自茎部生长的减弱,叶片生长反而增加,以支持繁殖生长[23],与苘麻通过茎的额外伸长而响应于密度增大的策略相反[30].但由于相关研究比较少见,不同物种的生物量分配对发芽季节的可塑性响应是否表现出相似的趋势并不清楚.
为更好理解不同物种如何通过生物量分配的可塑性响应于发芽季节的推迟,笔者选择了6种一年生草本植物(金狗尾、地肤、冬葵、野黍、狗尾、苋菜),在田间实验条件下,使它们在同一生长季节的不同时期(包括春季、夏季、秋季)发芽,并测量不同器官生物量及其分配,探讨以下问题:(1)植物如何通过不同器官生物量及其分配的可塑性响应于发芽季节的推迟;(2)不同植物的可塑性及响应策略是否有差异.
1 材料与方法
1.1 研究物种
6种一年生草本植物,包括金狗尾(Setariaglauca(L.) Beauv.)、地肤(Kochiascoparia(L.) Schrad.)、冬葵(Malvacrispavar.crispa)、野黍(Eriochloavillosa(Thunb.) Kunth)、狗尾(Setariaviridis(L.) Beauv.)和苋菜(AmaranthustricolorL.),物种信息见表1.金狗尾和狗尾在全球温带地区广泛分布,全株高可达20~90 cm,花果期一般在6—10月,开花后产生的种子可以在早春至初秋几个月内发芽,其生活周期短、种子萌发快和可塑性高等特点使其可以在短时间内成为地表优势种,常形成斑块状的单优种群生长在林缘和弃耕农田中[14,31-32].地肤分布于欧洲及亚洲,花期一般在6—9月,于7—10月结果,全株可高达50~100 cm,地肤等蔾科(Chenopodiaceae)植物是干旱、荒漠和盐碱地等较差环境中分布最广的植物之一,由于在长期适应较差生境的过程中导致藜科植物群落形成特殊的生殖物候和繁殖特征,因此具有重要的研究意义[33].冬葵源自中国南方地区,在全国各地都有分布,花果期一般在6—9月,其耐旱、耐贫瘠、繁殖力强、生命周期短等特点使其具有极强的适应环境的能力[34].野黍广泛分布于中国的华北、东北和西北地区,一般在9月结果,可高达30~100 cm,因其种子粒大,传播距离短,分蘖强易形成地表及地下的种群优势,常为弃耕地的先锋植物.苋菜花期一般在5—8月,果期在7—9月,可生长至80~150 cm,苋菜在中国的分布很广,品种资源丰富,生长快且生物量大,因此被作为重要的生物修复资源加以研究[35].在中国东北地区,这些一年生草本植物常见于干旱田地、路旁和荒芜的空地,分布范围较广,发芽季节范围较大,可能具有较强的可塑性,以适应自然生境中的各种环境变化,便于它们对发芽季节的可塑响应.另外,这些物种种子产量丰富且易于采集,种子萌发迅速且对土壤条件要求不高,幼苗生长极快且生活周期短[14],是理想的实验材料.

表1 6种植物的属性及名称的缩写
1.2 实验设计
研究地点在吉林省长岭县东北师范大学草地生态研究实验站(北纬44°45′;东经123°45′).该地区位于松嫩平原南部,属于半干旱半湿润温带季风气候,具有典型的大陆性季风气候特点:夏季炎热多雨,冬季寒冷干燥.植物主要生长季在6—8月,平均降水量占全年的70%~80%,7月平均气温最高(>20 ℃)[36].该实验在研究站的实验地进行,实验地土壤为pH=8.3的沙壤土,其养分含量为有机碳3.1 mg·kg-1,有效氮素21.0 mg·kg-1,有效磷1.1 mg·kg-1[37].所有植物物种的种子于2006年秋季,从研究站附近的野生种群中采集,并于—4 ℃的干燥条件下保存.
实验开始于2007年6月7日,采用裂区实验设计,根据3个不同的发芽季节(germination season,简称GS)共划分3个主区,分别进行3种发芽处理,即春季6月7日(GS1)、夏季7月7日(GS2)和秋季8月7日(GS3),在每个主区内,每种植物随机分布(表2).这些一年生草本植物物种不同发芽季节的设置符合中国东北吉林西部地区自然生境中的发芽季节范围.在吉林西部地区,春季一般开始于5月初,夏季开始于6月末,秋季开始于8月中旬,冬季开始于10月初.4—5月期间气温仍较低(4月0~15 ℃,5月10~20 ℃),降雨量不稳定.因此,将春季发芽处理设置于6月初,避免过高的幼苗死亡率.所有处理中,所有物种的植株播种时保持30 cm的间距,大多数种子在播种4~5 d后发芽,必要时除草,并定期浇水.

表2 6种植物的发芽季节、种植及取样时间、生长时长
1.3 数据收集和分析
6种植物中,由于在夏季发芽(GS2)的野黍、狗尾和苋菜在取样时可用植株过少,为确保数据分析的准确性和实验结果的代表性,剔除了这3种植物在夏季发芽(GS2)的数据,故这3种植物只包含2个发芽季节(GS1和GS3,表2).所有处理在植物繁殖中后期一次性取样.收获时,每种植物从每个处理中随机选取10株植物,因此实验的最大取样数量是3个处理×3种植物×10个重复+2个处理×3种植物×10个重复=150株.每株植物分为根、茎、叶、繁殖和侧枝部分(如果可用),并在75 ℃下烘干2 d,称重.繁殖部分包括沿主茎和枝条生长的花朵和果实,侧枝部分包括侧枝上的茎和叶.计算所有植株的总生物量(total mass,简称TM)以及根生物量(root mass,简称RM)、茎生物量(stem mass,简称SM)、叶生物量(leaf mass,简称LM)、繁殖生物量(reproductive mass,简称REM)、侧枝生物量(branch mass,简称BM)、根生物量比(root mass ratio,简称RMR)、茎生物量比(stem mass ratio,简称SMR)、叶生物量比(leaf mass ratio,简称LMR)、繁殖生物量比(reproductive mass ratio,简称REMR)和侧枝生物量比(branch mass ratio,简称BMR).
使用SPSS.20统计软件进行数据分析,用Origin.18软件作图.对于植物总生物量,采用双因素方差分析法(two-way ANOVA)分析物种、发芽季节及其二者交互作用的影响,并采用单因素方差分析法(one-way ANOVA)分析发芽季节或物种在每个或所有其他处理中的影响.植株大小(例如总生物量)可能会对其他特征产生非常显著的影响,导致表观可塑,与发芽季节处理造成的特征之间差异(真正的可塑)相混淆[38-39].因此,为去除个体大小的影响,将总生物量作为协变量(covariate),使用双因素协方差分析法(two-way ANCOVA)分析发芽季节、物种及其二者交互作用对所有生物量及分配特征的总体影响,并使用单因素协方差分析法(one-way ANCOVA)分析发芽季节对所有生物量及分配特征的影响.多重比较用一般线性模型(general linear model,简称GLM)程序中的最小显著差法(least significance difference,简称LSD),并在协方差分析(ANCOVA)中产生各个特征的调整后的平均值和标准误差[23].
2 结 果
2.1 总生物量
发芽季节、物种及其交互作用对总生物量(TM)都有极显著的影响,见表3所列.除冬葵(M)外,对于其他5种植物来说,总生物量随发芽季节的延迟而降低(图1).春季发芽的植物(GS1)平均总生物量为31.031 g,远高于秋季发芽的植物(GS3)平均值(0.82 g; ANOVA,p<0.001).夏季发芽(GS2)的金狗尾(Sg)和地肤(K)总生物量分别为1.30 g和6.01 g,与春季发芽(GS1)的植物有显著差异(p<0.001),夏季发芽(GS2)的冬葵(M)总生物量为18.58 g,与GS1差异不显著.
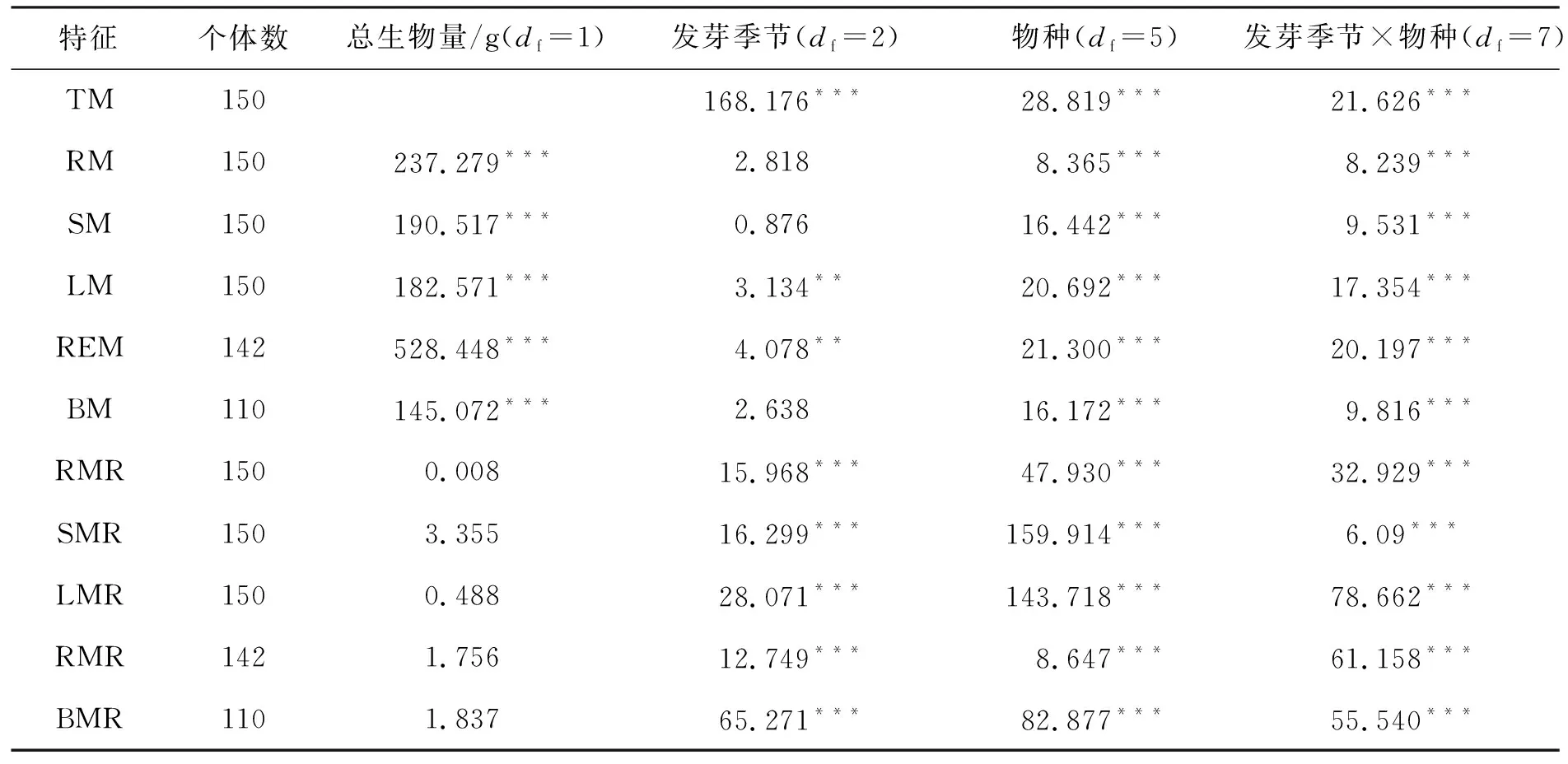
表3 发芽季节、物种及其交互作用对6种植物生物量和分配特征影响的双因素协方差分析

不同小写字母(a,b)表示同一物种在不同发芽季节存在显著性差异(ANOVA, LSD, p<0.05);GS1, GS2, GS3分别代表春季、夏季和秋季发芽季节.图1 6种植物在不同发芽季节的总生物量平均值
2.2 营养器官生物量及分配
总生物量对所选植物的各器官生物量和分配特征都有显著影响,当去除个体大小之后,发芽季节、物种及其交互作用对大多数特征的影响仍然显著(表3~4).在春季发芽(GS1)的金狗尾(Sg)和地肤(K)的根生物量显著高于GS2(ANCOVA,LSD,p<0.05),但GS3与GS1,GS2之间无显著差异;而冬葵(M)根生物量在夏季(GS2)时最高(p<0.001;图2).根生物量分配方面,金狗尾(Sg)和冬葵(M)随发芽季节(GS)的延迟而增加(p<0.001);而地肤(K)刚好相反,在GS1处理中最高,GS3处理最低,且差异明显(p<0.001;图3).
在茎生物量方面,GS2处理的物种冬葵(M)的茎生物量显著高于另外2个GS处理的(ANCOVA,LSD,p<0.001;图2).对于茎生物量分配,金狗尾(Sg)、苋菜(A)随发芽季节推迟而降低,地肤(K)随发芽季节推迟而增加;冬葵(M)在GS2时最高,GS3时最低(p<0.001;图3).
在叶生物量方面,就地肤(K)而言,GS1和GS3处理显著高于GS2的(ANCOVA,LSD,p<0.01;图2);冬葵(M)在GS2时叶生物量最高(p<0.001;图2).对于叶生物量分配,地肤(K)和苋菜(A)随发芽季节延迟而降低,但金狗尾(Sg)和冬葵(M)分别在GS3和GS2时最高(p<0.05;图3).
2.3 繁殖、侧枝生物量及分配
在繁殖生物量方面,随着发芽季节的延迟,GS3处理的野黍(E)的繁殖生物量显著高于GS1的(ANCOVA,LSD,p<0.001),GS2处理的地肤(K)最高(p<0.05),GS1的冬葵(M)显著高于GS2的(p<0.01;图2).对于繁殖生物量分配,地肤(K)和狗尾(Sv)都随着发芽季节的延迟而增加,在GS3时达到最大(p<0.001),而冬葵(M)和苋菜(A)随发芽季节的延迟而降低(p<0.001;图3).
冬葵(M)的侧枝生物量及其分配在GS3时显著高于GS1,GS2时的(p<0.01;图2);对于苋菜(A)来说,其侧枝生物量及其分配随着发芽季节的延迟而表现出增加的趋势(p<0.001;图3).

表4 发芽季节对6种植物生物量及分配特征影响的单因素协方差分析
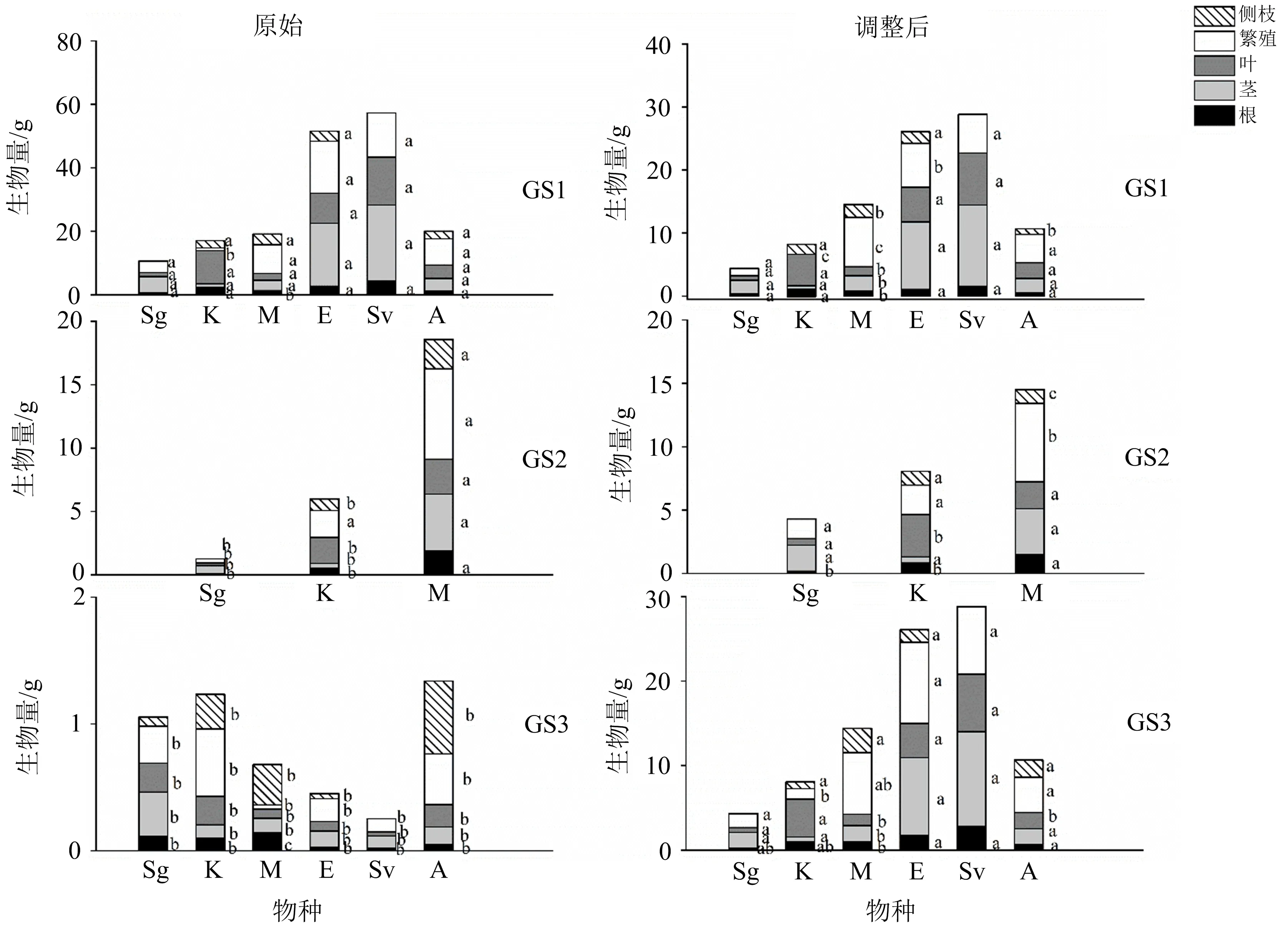
不同小写字母(a,b,c)表示同一物种的根、茎、叶、繁殖和侧枝生物量在不同发芽季节存在显著性差异(ANCOVA, LSD, p <0.05);Sg,K,M,E,Sv,A分别代表金狗尾、地肤、冬葵、野黍、狗尾和苋菜;GS1,GS2,GS3分别代表春季、夏季和秋季发芽季节.图2 6种植物各器官生物量在不同发芽季节的原始(左图)和调整后(右图)平均值
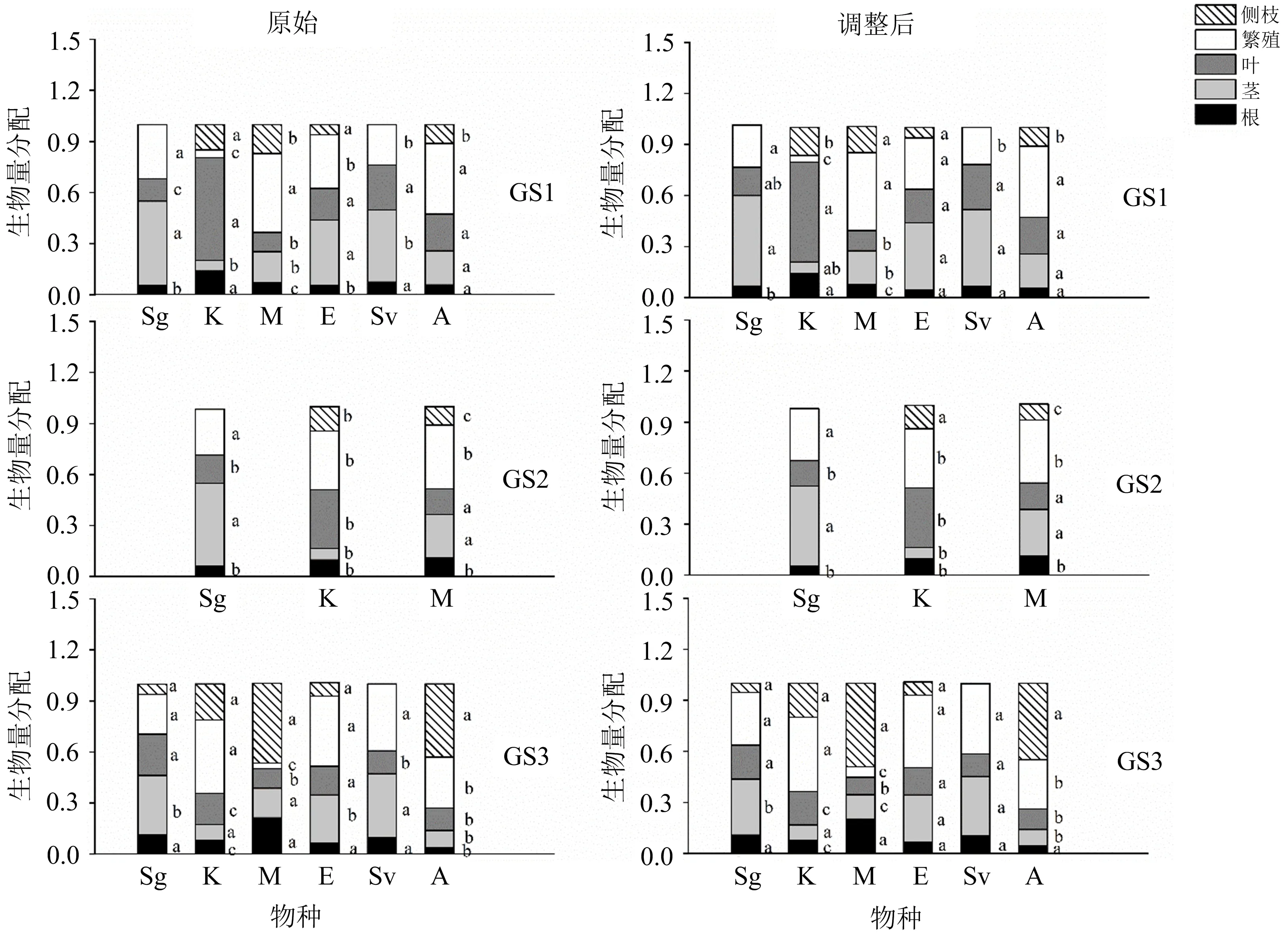
不同小写字母(a,b,c)表示同一物种的根、茎、叶、繁殖和侧枝生物量分配在不同发芽季节存在显著性差异(ANCOVA, LSD, p<0.05);Sg,K,M,E,Sv,A分别代表金狗尾、地肤、冬葵、野黍、狗尾和苋菜;GS1,GS2,GS3分别代表春季、夏季和秋季发芽季节.图3 6种植物各器官生物量分配在不同发芽季节的原始(左图)和调整后(右图)平均值
3 讨 论
许多一年生草本植物广泛分布于多变且易受干扰的环境中,可以在生长季的不同时期发芽.在季节早期发芽的幼苗通常比晚发芽的植株更有利,因为早发芽意味着有更多的时间去获取有限的资源以供生长繁殖[18].然而,延迟发芽也可以通过提高种子生产力使植物受益[17,22].同时,早发芽和晚发芽也都有各自的弊端,例如,早期发芽在提高资源获取时间的同时,也可能增加在季节早期遇到不利条件的风险[18-19,40],而延迟发芽的植物生活史周期变得有限.植物对不同发芽季节的选择还受到各种生物和非生物因素的综合影响[20].可能存在一个最佳发芽时间段,使植物在不可预测的环境中提高适合度从而使植物的整体性能得到优化[41].生物量分配是决定植物获取资源、竞争以及繁殖的重要因素[42],研究植物体内的资源分配到各器官的模式是研究植物生长格局变化的主要内容[43].在不同季节发芽的一年生草本植物往往通过改变和调整各器官的生物量分配以响应发芽季节的变化.这种可塑性响应对于解释一种植物的生活史策略有重要意义.该研究中,除冬葵在春季和夏季发芽总生物量表现没有显著差异外,其他5种植物总生物量都随发芽季节推迟而降低,但不同物种生物量分配对发芽季节的可塑性有显著差异,揭示了它们应对发芽时间变化的多样化的生活史策略.
3.1 资源获取优先策略
与春季(GS1)和夏季(GS2)发芽相比,秋季发芽(GS3)的金狗尾显著降低了茎生物量分配,增加了根和叶生物量分配,繁殖和侧枝生物量分配不受影响,基本符合晚发芽的植物缩短营养生长、提前繁殖的生活史策略[44].有研究表明,晚发芽的植物营养生长缩短主要通过降低茎生物量分配,增加根和叶生物量分配来实现[16,23].这是因为晚发芽的植物需要在更有限的时间内获取更多的地上和地下资源,叶片和根部分别是获取光照和养分最重要的器官,因此增加叶和根生物量分配有利于晚发芽的植物快速获取资源,积累生物量[3,45].不仅如此,在不存在其他个体或物种竞争光照的情况下,茎部的生长不仅无助于更多光照的获取,反而需要消耗更多资源成本.降低茎生物量分配,有利于植物快速获取资源,并节省用于营养生长的能量.因此,叶和根生物量分配增加,茎生物量分配降低,是植物采取的高效获取资源以快速生长的策略,有利于维持繁殖分配的稳定.金狗尾晚发芽的个体繁殖生物量分配没有显著高于早发芽的个体,一方面因为早发芽的个体可以利用更长的时间积累繁殖生物量,另一方面因为生命周期显著缩短导致晚发芽的个体生长潜力降低,不足以继续积累更高的繁殖生物量.
野黍与狗尾这2种植物对发芽季节推迟的响应不显著,倾向于提高季节晚期发芽的个体繁殖生物量分配.说明2种植物也能采取降低营养生长,加快繁殖生长的策略,以适应生命周期缩短的挑战.并且,由于它们的繁殖能力更强,并不需要过多减少用于营养生长的能量,也能维持稳定的繁殖水平.
3.2 垂直生长优先策略
与早发芽的个体(GS1和GS2)相比,秋季发芽(GS3)的地肤个体根和叶生物量分配显著降低,茎和繁殖生物量分配显著增加,与金狗尾营养器官对发芽季节的可塑性响应截然相反.这可能与地肤的物种特性有关.地肤属于常见的藜科一年生草本植物,是典型的荒漠植物,其生长的生境具有干旱少雨、土壤盐碱度高、降水时间变化大以及空间分布不均匀等特点[46].水分是影响地肤等荒漠植物生长的重要环境因子之一.所以,在湿度较高的春季(GS1)发芽并增加根和叶生物量分配有利于地肤吸收水和养分并进行旺盛的营养生长,以抵御随后夏季高温干旱的恶劣环境[46-47].这是地肤等荒漠植物长期适应干旱高盐碱等不利生境的结果.因此,早发芽的地肤通过增加根和叶等代谢活性器官的生物量分配有利于提高其总生物量表现.与早发芽(GS1和GS2)相比,秋季发芽的地肤茎和繁殖生物量分配显著增加.说明对于地肤来说,茎部可能是相对于根和叶片更重要的支持繁殖生长的器官,因此促进茎部生长是地肤植物在短生长季开始繁殖时进行资源储存的结果[48].有研究表明植物可以通过垂直生长策略使植物获得优势,尤其是在竞争条件下[49-50],在笔者的研究中地肤主要采取垂直生长优先策略来获取或储存用于繁殖生长所需的能量和资源,以减弱晚发芽带来的不利影响.在反枝苋(AmaranthusretroflexusL.)和绿珠藜(Chenopodiumacuminatum)2种植物的研究中发现,晚发芽植物会在植株较小时就可以开始繁殖生长,且繁殖生长开始的相对时间要早于先发芽的植物[44],因此繁殖生物量分配显著提高.这表明在季节晚期发芽的地肤也可以提前开始繁殖以便将更多的能量投入繁殖生长,有利于在更有限的生命周期内完成整个生活史.
3.3 侧向生长优先策略
随着发芽季节的推迟,冬葵根和侧枝生物量分配显著提高,繁殖生物量分配降低,茎和叶生物量分配先增大后减小,最终春季和夏季发芽的个体(GS1和GS2)总生物量没有显著差异,但都显著高于秋季发芽的个体(GS3).与冬葵相似,晚发芽(GS3)的苋菜茎、叶和繁殖生物量分配都低于早发芽的个体(GS1),但侧枝生物量分配显著提高.说明冬葵和苋菜早发芽的个体能够通过茎、叶和繁殖等器官获取更多的资源用于营养和繁殖生长,因此提高了植物的总体表现.对于这2种植物而言,最佳发芽时间可能在季节早期,这与Ferrise等[51]对硬质小麦(TriticumturgidumL. subsp.durum(Desf.) Husn)的研究结果一致.在秋季发芽(GS3)的苋菜和冬葵与春季发芽(GS1)相比,茎、叶和繁殖生物量分配都显著降低,但侧枝生物量分配都增加,说明苋菜和冬葵并未通过叶片生物量分配的提高,而是侧枝生长的增加,以响应发芽季节的推迟.这可能是由于晚发芽的苋菜和冬葵生长的侧枝主要用于繁殖,它们可以依靠侧枝上的繁殖器官的生长维持适合度水平的稳定,这与在低密度种群中通过增加侧枝等侧向生长结构而提高苘麻的繁殖生长的研究结果一致[50].有研究表明草本植物的分枝和侧枝等形态结构性状对植物的有性繁殖具有重要意义[52-53],是影响植物繁殖输出的重要因子[44].因此,晚发芽的冬葵和苋菜可以通过增加侧枝的生物量分配而提高未来的繁殖输出.尽管由于生长时间不足,这2种植物未能实现最后繁殖生物量分配的稳定或提高,但只要生长季稍微延长,侧枝的额外投入就可以得到收益.
4 结束语
笔者的研究表明,植物可以通过生物量分配的可塑性响应于发芽季节的推迟,揭示了不同物种采取的3种生长策略:
(1) 资源获取优先策略:晚发芽的金狗尾可以通过提高资源获取器官根和叶的生物量分配,降低茎生物量分配,以维持繁殖生长的稳定,野黍和狗尾由于具有更强的繁殖能力,可以不通过降低营养生长而维持稳定的繁殖分配;
(2) 垂直生长优先策略:地肤通过降低根和叶生物量分配,提高茎生物量分配以促进繁殖生长;
(3) 侧向生长优先策略:冬葵和苋菜通过侧枝生物量分配的提高而争取繁殖生长的稳定.
在对发芽季节的响应过程中,尽管不同物种调整营养器官生物量分配的方式不同,最终目标都是达到稳定甚至更高水平的繁殖分配,以争取在有限的生长时间内完成整个生活史,保证种群后代的延续.该研究只观测了不同物种在不同季节发芽的最终表现,未来可以有更多研究关注不同类型植物响应于发芽季节的可塑性,包含更多方面和水平的特征的动态变化,揭示不同物种应对发芽季节变化的生长策略,探讨植物在真实的自然环境下适应复杂环境变化的机制.
