长江支流大宁河巫溪段鱼类群落结构的年际变化及保护对策
2023-04-29唐会元朱其广金瑶董纯赵娜陈小娟李德旺
唐会元 朱其广 金瑶 董纯 赵娜 陈小娟 李德旺

摘要:研究大宁河巫溪自然流水河段鱼类群落结构的年际变动特征,为大宁河鱼类资源保护提供依据。2011-2022年的6-7月和11-12月,在大宁河下游、巫溪县城以上的流水河段7个区域,使用定置刺网和虾笼捕捞鱼类,研究种类组成、优势种、物种多样性、群落结构及其变动特征,分析三峡水库175 m正常蓄水后该河段鱼类群落结构的年际变化特征及其影响因素。结果显示:调查河段有鱼类6目13科52属72种,其中长江上游特有鱼类9种、国家二级重点保护动物3种,优势种7种;鱼类种类组成在各年间的Jaccard指数值在0.85~1.00,差异不显著;鱼类的Margalef 丰富度指数和Shannon-Wiener多样性指数在年际间呈现先上升后下降再上升的趋势,而Pielou均匀度指数则呈现先下降后上升的趋势;鱼类群落结构在2016年和2018年时发生了明显的变化,渔获物中喜流水性的长江上游特有鱼类或国家重点保护鱼类的相对丰度下降,广布型的小型鱼类相对丰度明显上升。建议通过实施就地保护、生态景观修复、非土著鱼类控制等方式,加强对巫溪江段特有及重要鱼类种类的保护。
关键词: 物种多样性;群落结构;年际变动;保护对策;大宁河
中图分类号:S932.4 文献标志码:A 文章编号:1674-3075(2023)06-0063-09
鱼类群落的主要特征,包括物种多样性、优势种及群落结构等,是鱼类生态学中的主要调查与研究内容之一(殷名称, 1995)。基于时空梯度调查,辨识鱼类群落在时间和空间尺度上的分布特征,进而发现其时空差异性,一直以来是国内外鱼类群落结构研究重点之一(Gao et al, 2010; Lin et al, 2019; 刘燕山等, 2021; Yang et al, 2021)。通过辨识鱼类群落结构的时空格局变动,明晰其发生变化的时间节点或空间位置,可以在整合人类活动胁迫信息下,确定影响其变动的原因,并进而获得针对性的鱼类资源保护措施(刘燕山等, 2021; Yang et al, 2021; 邱龙辉等, 2023)。
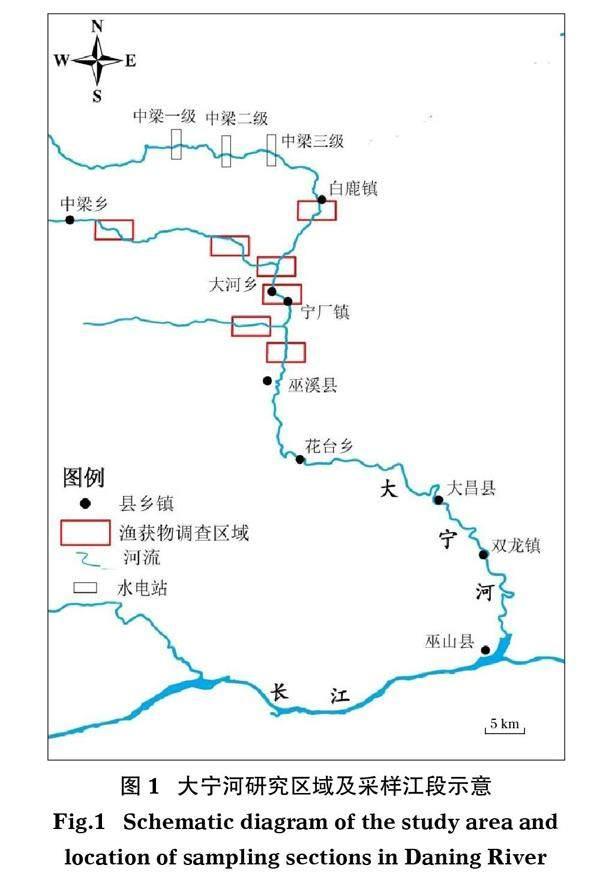
大宁河古称巫溪,地处三峡库区左岸,主流西溪河发源于陕西省平利县的中南山麓,其源头汇集东溪、西溪、后溪、马连溪等小溪而成。西溪河与东溪河汇合后称大宁河,干流穿行于巴山和巫山的云岩峡谷之中,横贯重庆所属巫溪、巫山两县,在巫山县巫峡口注入长江。大宁河全长250 km,其中受三峡水库调度运行影响的河段长约143 km(张佳磊等, 2012)。175 m水位下,三峡水库回水区末端在巫溪县花台乡上游;巫溪县城及以上河段为流水河段。流水河段为山地峡谷溪流生境河段,底质以卵石、砾石和基岩为主;河道海拔830 m以上,多年平均气温18℃左右,不仅为许多冷水性鱼类如裂腹鱼属和高原鳅属鱼类的分布区域(冯家兄等, 2014;张雷等, 2017),而且也为国家二级重点保护动物多鳞白甲鱼的重要分布区域(杨峰等, 2013; 唐容, 2020)。迄今,已有三峡水库蓄水初期(2008-2009年以及2011-2012年)大宁河巫溪流水河段的鱼类资源状况研究(耿相昌, 2010; 杨峰等, 2013),但三峡水库175 m蓄水运行后(2010年10月26日,最高水位达到175 m),有关大宁河巫溪流水河段鱼类群落结构时间变动的研究暂无报道。随着三峡水库蓄水运行年份的延长,大宁河下游回水区河段中适应静缓流生境的鱼类是否会进入上游自然流水河段,若进入对流水河段鱼类的群落结构会造成什么影响,需要进行研究。本文于长江流域十年禁捕前在大宁河巫溪流水河段进行渔获物调查,探讨大宁河巫溪自然流水河段鱼类群落结构的时间变动特征,为大宁河鱼类资源保护提供依据。
1 材料与方法
1.1 调查区域及样本采集
调查区域位于大宁河下游、巫溪县城以上的流水河段(图1),7个采样区域包括干流白鹿镇-巫溪县城间的5个采样区域、支流中梁乡下游的2个采样区域。调查时间为2011-2020年,每年6-7月、11-12月在上述7个采样区域各开展1次渔获物调查,每次调查5d。
聘请当地专业渔民使用2种网具(定置刺网和虾笼)在指定调查区域捕捞。定置刺网网目大小为20 ~250 mm(包括20、40、80、100、120、200和250 mm),网长100~200 m;虾笼网目大小为8 mm。定置刺网放置在大型岩石的间隙以及水流缓慢的沿岸带或湾沱中(杨志等, 2015);虾笼放置在岩石密布或河道覆盖物复杂的区域。定置刺网和虾笼被放置的时间为17:00-19:00至翌日04:00-06:00,该时段鱼类活动最频繁(Blabolil et al, 2017)。为保证数据的可比性,各年定置刺网和虾笼的规格、放置地点及作业时间均基本一致。现场对采集到的所有样本进行种类鉴定和计数,并逐尾测量全长、体长(精确到1 mm)和称体重(精确到0.1 g)(杨志等, 2015)。
1.2 数据统计与分析
对采集到的鱼类种类的拉丁名(Fricke & Eschmeyer, 2023)进行核对,剔除同物异名或错误命名的种类。在此基础上,基于各年鱼类种类组成的有无数据,构建种类组成的Jaccard相似性矩阵,并采用组平均连接的聚类分析(Cluster analysis,CA)方法,分析鱼类种类组成在年际间的分组情况(Clarke & Gorley, 2006;郭朋军等,2020)。针对CA结果,采用One-way ANOSIM方法,检验鱼类种类组成在各组之间是否具有显著性的差异。同时,采用以下生物多样性指数计算公式,计算研究区域各年各多样性指数的值,并分析其年际变动特征(殷名称, 1995):
上述各式中,S为总种类数,N为所有种类的个体数,Pi为第i种的个体数。同时,参考文献(Pinkas & Iverson,1971)计算不同种类在不同年份上的相对优势度(Index of Relative Importance,IRI),IRI大于2%的为优势种。
计算不同鱼类种类在各年渔获物中的数量百分比,并将该数量百分比作为各种鱼在各年采样期间的相对丰度。基于上述相对丰度数据,构建Bray-Curtis相似性矩阵,并采用CA方法,分析鱼类群落结构在年际间的分组情况(Clarke & Gorley, 2006)。构建Bray-Curtis相似矩阵前,对相对丰度数据进行lg(x+1)转换;在进行CA分析之前,为避免稀有物种对分析结果的不确定性影响,将每年渔获物中相对丰度均小于0.5%的鱼排除(郑梦婷等, 2023)。针对CA结果,采用One-way ANOSIM方法,检验鱼类群落结构在各组之间是否具有显著性的差异。在此基础上,采用百分比相似性分析方法(Similarity Percentage,SIMPER)获得引起鱼类群落结构在不同组间差异的主要种类(Clarke & Gorley, 2006);使用公式④计算主要种类的相对丰度在不同聚类组间的变化情况(郑梦婷等, 2023),参考文献(杨少荣, 2012; 梁秩燊等, 2019)确定主要种类对流速的偏好情况。
式中:Rav 为主要鱼类种类的相对丰度在不同聚类组之间的变化(%);G1ra、G2ra分别为主要物种在聚类组1和聚类组2中的相对丰度(%)。
使用软件PRIMER-E (Version 6) 和 PERMANOVA+ (Anderson et al, 2008)进行CA、ANOSIM 和SIMPER分析。本文的显著性水平设置为P<0.05。
2 结果与分析
2.1 鱼类种类组成及年际变动
2011-2022年,在调查区域统计渔获物1 245.21 kg、2 4765尾,鉴定出鱼类种类72种,隶属于6目13科52属(表1)。鲤科鱼类种类数最多,31属39种,占总种类数的54.17%;鲿科和鳅科分别为4属9种和6属8种,各占12.50%和11.11%;平鳍鳅科和鰕虎鱼科各采集到3种,各占4.17%;钝头鮠科和鮨科各采集到2种,各占2.78%。极危等级1种(胭脂鱼)、易危等级6种(厚颌鲂Megalobrama pellegrini、多鳞白甲鱼、齐口裂腹鱼、岩原鲤、细体拟鲿Pseudobagrus pratti和白缘?Liobagrus marginatus),长江上游特有鱼类9种,国家二级重点保护动物3种(胭脂鱼、多鳞白甲鱼和岩原鲤)。
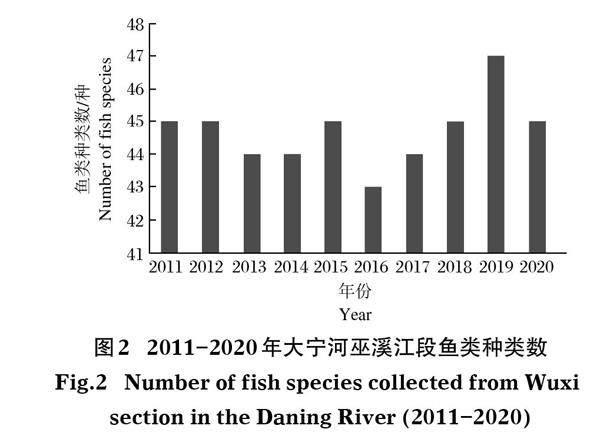
各年采集到的鱼类种类数在43~47种,2016年种类数最少,2019年种类数最多(图2)。聚类分析结果显示:鱼类种类组成在各年间的Jaccard指数值在0.85-1.00之间,平均值为0.93,可以分为2个聚类组,其中组A包括2016-2017年的鱼类种类组成,组B包括其它8个年份的鱼类种类组成(图3)。One-way ANOSIM检验显示种类组成在两组间差异不显著(r=0.556,P=5.9%,迭代次数136次)。
2.2 物种多样性及年际变动
调查期间,大宁河巫溪江段鱼类生物多样性指数值的年际变动情况如图4。巫溪江段鱼类的R 和H在年际间呈现先上升后下降再上升的趋势,J则呈现先下降后上升的趋势。R和H在各年间的变动范围分别为8.30~9.64、2.83~3.05,平均值分别为8.89、2.95,最大值和最小值出现的年份均分别为2015年、2017年;J各年间的变动范围为0.84~0.87,平均值为0.86,最大值和最小值出现的年份分别为2011年、2017年。
2.3 优势种及其年际变动
2011-2020年巫溪江段有优势种鱼类9种(图5)。云南盘鮈Discogobio yunnanensi的年均相对重要性指示值最高,为20.32%;切尾拟鲿Pseudobagrus truncatus最低,为4.17%。
优势种类中,有长江上游特有鱼类2种(齐口裂腹鱼和宽口光唇鱼)、国家二级重点保护动物1种(多鳞白甲鱼)。相对优势度在各年间的变化(图6),齐口裂腹鱼呈现先下降后上升再下降的趋势,多鳞白甲鱼呈现先下降后上升且总体上呈下降的趋势,宽口光唇鱼呈现先上升后波动下降的趋势(2020年比2011年减少了73.16%)。
2.4 群落结构及其年际变动
调查期大宁河巫溪江段鱼类群落结构的聚类分析结果见图7。在83.23%的Bray-Curtis相似性水平上可将该江段10年的鱼类群落结构分为3大组:组1包括2013-2015年的鱼类群落结构,组2包括2016-2017年的鱼类群落结构,组3包括2018-2020年的鱼类群落结构。One-way ANOSIM检验显示各大组间及任意2大组间的鱼类群落结构在统计学上差异显著(各组间:r=0.995,P=0.1%,迭代次数999次;组1 vs组2:r=0.982,P=4.8%,迭代次数21次;组1 vs 组3:r=1.000,迭代次数56次;组2 vs 组3:r=1.000,迭代次数10次)。
聚类分析的百分比相似性指数(SIMPER)分析结果见表2。引起鱼类群落结构在组1和组2间差异的主要种类有11种,其中大鳞副泥鳅丰度变动引起的差异贡献率最高,为9.25%,其次为宽口光唇鱼(7.82%)和切尾拟鲿(6.96%);引起鱼类群落结构在组2和组3间差异的主要种类有11种,其中切尾拟鲿丰度变动的贡献率最高,为8.93%,其次为贝氏?Hemiculter bleekeri(8.48%)和大鳞副泥鳅 (6.01%);引起鱼类群落结构在组1和组3间差异的主要种类有13种,其中宽口光唇鱼的贡献率最高,为11.47%,其次为贝氏?(7.89%)和齐口裂腹鱼(6.94%)。宽口光唇鱼、多鳞白甲鱼等喜流水性的长江上游特有鱼类或国家重点保护鱼类在渔获物中相对丰度的减少,以及广布型的小型鱼类如贝氏?和麦穗鱼Pseudorasbora parva等在渔获物中相对丰度的明显增加,是导致巫溪江段鱼类群落结构在年际间发生分离的主要原因。
3 讨论
3.1 鱼类种类组成及物种多样性变动特征
10年间共在巫溪河段采集到鱼类72种,占历史上大宁河流域鱼类99种(耿相昌, 2010)的72.73%,表明巫溪河段是大宁河鱼类资源非常丰富的河段,保护该河段的鱼类物种多样性对于大宁河鱼类资源的保护具有重要的意义。在本河段中,分布有较多的适应流水生境的长江上游特有鱼类或其他鱼类,这与耿相昌2008-2009年(耿相昌,2010)和杨峰2011-2012年(杨峰,2013)在巫溪河段调查的结果一致。本次未调查到泸溪直口鲈和云南光唇鱼这2种鱼,可能与采样误差有关。此外,结果显示各年的种类组成尽管可以分为2组,但是在两组间差异不显著,表明巫溪河段的鱼类种类组成在2011-2020年间无明显的变化。
总体上,巫溪河段的鱼类物种丰富度及Shannon-Wiener多样性指数值呈现略微上升的趋势,与董纯等(2019)在库区4个干流江段(秭归、巫山、云阳和涪陵)的研究结果相似。原因可能包括以下2个方面:(1)三峡水库形成后,库区许多江段的水文生境发生了明显改变,适应静缓流生境的鱼类种类数增多,如原分布在长江中下游的太湖新银鱼Neosalanx taihuensis、短颌鲚Coilia brachygnathus、团头鲂Megalobrama amblycephala等以及外来鱼类种类在库区逐渐出现(董纯等, 2023; 郑梦婷等, 2023; 张伟等, 2023),部分种类进入了干支流上游的流水江段或河段;(2)巫溪河段仍保持自然或近自然的流水生境,仍是喜流水性鱼类的分布区域,喜流水性鱼类的种类数并没有明显减少,在一定程度上维持了研究河段物种多样性的稳定。
3.2 鱼类群落结构变动特征
三峡水库175 m正常蓄水后,巫溪流水河段的鱼类群落结构已发生了明显的变化,与Yang Z等(2021)在具有一定长度流水生境的三峡库区干流回水区江段研究结果相似。目前,大宁河干流仅在上游西溪河上修建了中梁水库及其一、二、三级电站(2011年底时均已完工);巫溪以上的较长河段均为流水生境河段(于子铖等, 2023)。因此,巫溪河段的鱼类群落结构变动受到上游梯级水库蓄水运行的影响应该较小。同时,调查发现许多广布型的鱼类如贝氏?、子陵吻鰕虎鱼等的资源丰度在2015年以后明显增加且均为三峡水库蓄水后数量才明显增加的种类(杨志等, 2015)。在三峡水库蓄水前,这些鱼并不是大宁河砾石激流生境中分布的常见或主要鱼类种类(丁瑞华, 1994; 曹文宣等, 2007),因此可以推知这些种类的多数个体应该来自于大宁河下游的回水区江段,说明巫溪自然流水江段鱼类群落结构的年际变化也受到了三峡水库蓄水运行的明显影响。已有研究表明,水库内鱼类物种的扩散,会导致水库上游河段内鱼类群落结构同质性的变化(Franchi et al,2014;Pasquaud et al 2015);生物间关系(如低阶或高阶组合的种间关系、生态演替过程、物种散布及占据等)对群落结构的组成及变动具有重要的意义(Bairey et al, 2016; Paller et al, 2016)。
宽口光唇鱼、多鳞白甲鱼等喜流水性的鱼类在渔获物中相对丰度的下降,是导致巫溪江段鱼类群落结构在年际间发生明显变动的重要原因之一,表明近年来巫溪河段鱼类资源保护面临的突出问题是长江上游特有鱼类或国家重点保护鱼类的资源衰减。这类鱼多为以底栖无脊椎动物或/和着生藻类为主要食物、在砂砾或岩石间隙产沉性或粘沉性卵的鱼类种类(曹文宣等, 2007);其种群维系与河道基质的分布特征密切相关:较大粒径的砾石或石质基质为鱼类饵料提供附着或庇护基质,而较小粒径的砂砾则是这些鱼类适宜产卵场生境的重要组成部分。现场调查发现,大宁河沿岸城镇及村落集中分布,近年来,河岸带固化、河道采砂以及沿岸公路、路桥等的施工建设很常见。已有调查也发现,相较之2010年,2018年大宁河的林地、城镇用地、农村居民点及其他城镇用地呈明显的增加趋势(勾蒙蒙等, 2023)。上述情况均很可能导致大宁河底质特征的变化,而底质特征的变化又会通过影响鱼类的饵料来源及产卵基质特征来影响鱼类的种群动态(杨晓鸽等, 2021)。随着社会经济的发展,人们对野生鱼类的需求量增加,在捕捞技术发展的背景下,渔业捕捞效率提高及强度增加(谢平, 2017),很可能也是研究区域鱼类群落结构变动的原因。调查发现,从2015年起,齐口裂腹鱼在巫溪河段渔获物中的相对优势度呈明显的上升趋势,这很可能与其大量地被人工放流有关。由于齐口裂腹鱼与宽口光唇鱼、多鳞白甲鱼在食物组成上的相似性,因此食物竞争很可能是导致宽口光唇鱼、多鳞白甲鱼等鱼类资源量减少的重要原因之一。
3.3 大宁河巫溪江段鱼类资源保护措施
考虑到宽口光唇鱼、多鳞白甲鱼等长江上游特有或国家重点保护鱼类的栖息、觅食和产卵的适宜生境条件(流水、砾石浅滩),以及区域岸坡固化、路政建设及河道采砂等对河床基质的影响,建议从栖息地的就地保护及生境修复两个方面开展工作。大宁河干流白鹿镇上游修建了中梁水库及其一、二、三级电站,中上游的支流中也修建了较多的水电站(陶涛等,2017),巫溪花台乡及其下游河段为三峡水库蓄水倒灌的河段,因此,目前大宁河的流水河段主要集中在白鹿镇至花台乡河段。该河段是大宁河喜流水性的特有或保护鱼类的主要分布河段,应予以优先保护。建议在通过调查,确定特有或保护鱼类“3场1通道”的具体分布位置的前提下,实施就地保护措施,建立面向鱼类关键生境保护的自然保护区,避免人类活动(特别是河道内工程建设以及商业性的垂钓)对这些关键区域的严重干扰。对于已受到破坏的河漫滩以及非生态固化的岸坡,应采用生态景观修复技术进行生态修复。
广布型鱼类逐渐扩散到巫溪流水河段,并改变了该河段的鱼类群落结构。考虑到这类鱼通常为杂食性的小型鱼类,且食谱宽、摄食能力较强,应通过持续的监测,分析了解这些鱼类摄食及栖息生境占据对巫溪河段土著鱼类的影响。当这些鱼类的种群数量过大,极有可能影响土著鱼类的栖息觅食时,应采取定向捕捞或修建隔离措施的方式对其种群规模进行控制。另外,巫溪县城下游的回水区江段已出现了大量的短颌鲚、罗非鱼等外来鱼类(董纯等, 2023),这些外来鱼类未来是否会散布到巫溪流水河段并大量增殖,需要持续地关注。
参考文献
曹文宣, 常剑波, 乔晔, 等, 2007. 长江鱼类早期资源[M]. 北京: 中国水利水电出版社.
董纯, 杨志, 朱其广, 等, 2023. 三峡水库外来鱼类资源状况初步研究[J].长江流域资源与环境, 32(5): 928-938.
丁瑞华, 1994. 四川鱼类志[M]. 成都:四川科学技术出版社.
冯家兄,陈沐希,闫雪, 2014. 论巫溪县大宁河流域气候地域特征[J]. 环球人文地理,(16):33.
耿相昌, 2010. 大宁河鱼类资源现状及云南盘鮈生物学初步研究[D]. 重庆: 西南大学.
郭朋军, 江新琴, 俞存根, 等, 2020. 舟山沿岸渔场春秋季鱼类群落结构特征分析[J]. 渔业科学进展, 41(4): 1-11.
勾蒙蒙, 刘常富, 李乐, 等, 2023.三峡库区典型流域生境质量时空演变特征与情景模拟[J]. 生态学杂志, 42(1): 180-189.
刘燕山, 谷先坤, 唐晟凯, 等, 2021. 东太湖鱼类群落结构特征及其与环境因子的关系[J]. 生态学报, 41(2): 769-780.
梁秩燊, 易伯鲁, 余志堂, 2019. 江湖鱼类早期发育图志[M]. 广州: 广东科技出版社.
唐容, 2020. 任河和大宁河多鳞白甲鱼生物学及遗传学特征的比较研究[D]. 重庆:西南大学.
陶涛, 李玉桥, 2017. 重庆市大宁河干流水电开发方案研究—与南水北调中线三峡水库补水工程无缝对接[J]. 水利规划与设计, (8): 24-26,140.
谢平, 2017. 长江的生物多样性危机——水利工程是祸首, 酷渔乱捕是帮凶[J]. 湖泊科学, 29(6): 1279-1299.
杨少荣, 2012. 长江流域鱼类群落生态学研究[D]. 北京: 中国科学院大学.
殷名称, 1995. 鱼类生态学[M]. 北京: 中国农业出版社.
杨 志, 唐会元, 朱迪, 等, 2015. 三峡水库175 m 试验性蓄水期库区及其上游江段鱼类群落结构时空分布格局 [J]. 生态学报, 35(15): 5064-5075.
杨峰, 姚维志, 邓华堂, 等, 2013. 三峡库区蓄水后大宁河鱼类资源现状研究[J]. 淡水渔业, 43(4): 51-57.
杨晓鸽, 范传文, 鲍宇轩, 等, 2021. 阊江上游河道整治底质粒径变化对鱼类群落多样性的影响[J]. 长江流域资源与环境, 30(10):2430-2437.
于子铖, 赵进勇, 彭文启, 等, 2023, 小水电河流水文地貌-生态响应关系研究[J]. 水利水电技术(中英文), 54(6): 137-146.
张佳磊, 郑丙辉, 刘录三, 等, 2012. 三峡水库试验性蓄水前后大宁河富营养化状态比较[J]. 环境科学, 33(10): 3382-3389.
张雷,时瑶,张佳磊,等, 2017. 大宁河水生态系统健康评价[J]. 环境科学研究,30(7):1041-1049.
张伟, 翟东东, 熊飞, 等, 2023. 三峡库区鱼类群落结构和功能多样性[J]. 生物多样性,31(1): 22136.
郑梦婷, 杨志, 胡莲, 等, 2023. 小江回水区江段鱼类群落结构的时空变动及驱动因素分析[J]. 水生态学杂志, 44(3): 42-53.
Bairey E, Kelsic E D, Kishony R, 2016. High-order species interactions shape ecosystem diversity[J]. Nature Communications, 7: 12285.
Blabolil P, ?íha M, Ricard D, et al, 2017. A simple fish-based approach to assess the ecological quality of freshwater reservoirs in Central Europe[J]. Knowledge & Management of Aquatic Ecosystems, 418:53.
Clarke K R, Gorley R N, 2006. PRIMER V6: User manual/tutorial PRIMER-E[M]. Plymouth,UK: PRIMER-E.
Franchi E, Carosi A, Ghetti L, et al, 2014. Changes in the fish community of the upper Tiber River after construction of a hydro-dam[J]. Journal of Limnology, 73(2): 203-210.
Fricke R, Eschmeyer W N, Van der Laan R, 2020. Eschmeyer's catalog of fishes: Genera, species, references[P]. http://researcharchive.calacademy.org/research/ichthyology/catalog/fishcatmain.asp. Electronic version accessed 12 July 2023.
Gao X, Zeng Y, Wang J W, et al, 2010. Immediate impacts of the second impoundment on fish communities in the Three Gorges[J]. Environmental Biology of Fishes, 87(2): 163-173.
Lin P C, Gao X, Liu F, et al, 2019. Long-term monitoring revealed fish assemblage zonation in the Three Gorges Reservoir[J]. Journal of Oceanology and Limnology, 37: 1258-1267.
Paller MH, Prusha BA, Fletcher DE, et al. Factors influencing stream fish species composition and functional properties at multiple spatial scales in the sand hills of the southeastern United States[J]. Transactions of the American Fisheries Society, 2016, 145: 545-562.
Pinkas L, Oliphant MS, Iverson K, 1971. Food habits of albacore bluefin tuna and bonito in Californian waters [J]. California Department of Fish and Game: Fish Bulletin, 152: 1-105.
Pasquaud S, Vasconcelos RP, Franca S, et al, 2015. Worldwide patterns of fish biodiversity in estuaries: effect of global vs. local factors[J]. Estuarine, Coastal and Shelf Science, 154:122-128.
Yang Z, Pan X J, Hu L, et al, 2021. Effects of upstream cascade dams and longitudinal environmental gradients on variations in fish assemblages of the Three Gorges Reservoir[J]. Ecology of Freshwater Fish, 30(4):503-518.
(责任编辑 张俊友)
Interannual Variations of Fish Community Structure and Measures to
Preserve Fishery Resources in the Wuxi Section of Daning River
TANG Hui‐yuan, ZHU Qi‐guang, JIN Yao, DONG Chun, ZHAO Na, CHEN Xiao‐juan, LI De‐wang
(Key Laboratory of Ecological Impacts of Hydraulic-projects and Restoration of Aquatic Ecosystem
of Ministry of Water Resources, Institute of Hydroecology, Ministry of Water Resources
and Chinese Academy of Sciences, Wuhan 430079, P.R. China)
Abstract:Daning River is located on the left bank of Three Gorges Reservoir and is affected by its regulation and operation. In this study, we analyzed the interannual variation and influencing factors of fish community structure in the Wuxi section of Daning River at the 175 m impoundment level of Three Gorges Reservoir, focusing on species composition, dominant species, and community diversity. Meanwhile, measures for protecting fish resources in this section were suggested. Our aim was to provide evidence for preserving fish resources in Daning River. The river area investigated was the section with natural running water in the lower Daning River above Wuxi County. In June-July and November-December during the period 2011-2022, an annual fishery survey was carried out at seven sampling sites using gill nets and shrimp cages. The fish collected were counted and identified in situ, and total length, body length and weight of all samples were measured. A total of 72 fish species from 52 genera, 13 families and 6 orders were collected during the 10 years, including 9 species endemic to the upper Yangtze River and 3 species were national second-class protected animals (Myxocyprinus asiaticus, Onychostoma macrolepis, and Procypris rabaudi). The number of fish species collected each year was in the range 43-47, with the highest richness of fish species in 2019 and the lowest in 2016. Jaccard coefficient values, indicating fish species composition, were in the range 0.85-1.00 during the study period, and there were no significant differences in species composition among the years. Margalef richness (R) and Shannon-Wiener diversity indices (H) both increased initially, then decreased, and increased again. The Pielou evenness index (J) decreased initially and then increased. There were 7 dominant species in the study area, including 2 fish species endemic to the upper Yangtze River (Schizothorax prenanti and Leptospira latifolia) and 1 fish species of the national key protected animal (Onychostoma macrolepis). The fish community structure changed obviously in 2016 and 2018. The relative abundance of species that prefer running water, and are endemic to the upper reaches of the Yangtze River and species under special state protection decreased, while small, highly adaptable fish clearly increased, resulting in obvious interannual changes in the fish community structure of the Wuxi section. The operation of Three Gorges Reservoir played an important role in the change of fish community structure in the Wuxi section. It is suggested that the protection of endemic and important fish species in the Wuxi section of Daning River should be strengthened by implementing in-situ protection, ecological landscape restoration and non-indigenous fish control.
Key words:species diversity; community structure; inter-annual variation; protection measures; Daning River
收稿日期:2023-07-06
基金项目:“十四五”国家重点研发计划课题(2021YFC3200304);水利部重大科技项目(SKR-2022015);金沙江下游流域水生生态监测(JG/19018B)。
作者简介:唐会元,1964年生,研究员,主要从事鱼类生态学研究。E-mail:751148270@qq.com
通信作者:李德旺,主要从事水生态与水环境研究。E-mail:64644208@qq.com
