利用分子标记辅助选择培育优良食味、低谷蛋白香粳稻新品系
2023-01-18陈涛赵庆勇朱镇赵凌姚姝周丽慧赵春芳张亚东王才林
陈涛 赵庆勇 朱镇 赵凌 姚姝 周丽慧 赵春芳 张亚东* 王才林*
(1江苏省农业科学院 粮食作物研究所/江苏省优质水稻工程技术研究中心/国家水稻改良中心 南京分中心,南京 210014;2扬州大学 江苏省粮食作物现代产业技术协同创新中心, 江苏 扬州 225009; *通信联系人, email: zhangyd@jaas.ac.cn; clwang@jaas.ac.cn)
我国是世界最大的稻米生产国,年种植面积约3000万hm2,占全球水稻总种植面积的近20%,年产稻谷超过2.1亿t,约占全球稻谷总产量的30%[1]。同时,水稻也是我国是第一大主粮作物,约占居民口粮消费总量60%[2]。稻米在为人们提供碳水化合物的同时,也是植物性蛋白质的重要来源。研究表明,稻米蛋白质含量约占糙米干质量的8%~10%,是仅次于淀粉的第二大类贮藏物质,其含量与稻米营养价值密切相关[3]。根据溶解特性的不同,稻米贮藏蛋白分为溶于稀酸或稀碱溶液的谷蛋白、溶于稀盐溶液的球蛋白、溶于水的清蛋白以及少量醇溶蛋白。其中,醇溶蛋白含量占20%~25%,贮存在致密的Ⅰ型蛋白体中,不能被人体消化,而由37~39 kD α酸性亚基和22~23 kD β碱性亚基组成,以Ⅱ型蛋白体形式存在的谷蛋白含量较高,约占种子贮藏蛋白的60%~75%,是稻米蛋白中可供人体吸收的主要成份[4]。因此,稻米营养价值的改善可以通过提高谷蛋白含量得以实现。
随着我国城乡居民生活方式和饮食结构的改变,肾脏病的发病率逐年升高,已成为威胁公共健康安全的重要疾病。据统计,我国慢性肾脏病人1.32亿,患病率高达10.8%,其中住院患者88.8万,占住院总人数的4.8%[5]。由于肾脏病患者常伴有肾机能不全、蛋白代谢紊乱等症状,要减轻肾脏负担,除进行必要的药物治疗外,还必须在饮食中严格限制病人对可溶性蛋白的摄入。目前市场上可供肾脏病专用的低蛋白食品不仅种类稀少而且价格昂贵,这无疑增加了患者的经济和精神负担,降低了治疗效果[6]。因此,为满足肾脏病人对该类食品的特定需求,培育综合性状优良的低谷蛋白水稻新品种已成为当前功能性水稻育种的一个重要方向。
低谷蛋白水稻的研究始于日本。Iida等[7-8]利用化学诱变剂乙烯亚胺处理水稻Nihonmasari的种子,筛选到低谷蛋白-高醇溶蛋白种质NM67,并在与原始亲本杂交的F2代中得到农艺性状较优的育种新材料“LGC-1”。遗传分析表明,LGC-1的低谷蛋白特性受单个显性基因Lgc1控制,该基因位于水稻第2染色体标记XNpb243和G365之间[9]。进一步研究发现,在LGC-1中核苷酸序列相似性达99.8%的B亚族谷蛋白基因GluB4和GluB5之间存在一个3.5 kb的片段缺失,从而形成一个尾对尾的反向重复序列,该序列产生双链RNA发夹结构,进而诱导RNA沉默,导致谷蛋白含量降低[10]。
在育种利用方面,日本育种家以LGC-1为亲本先后选育出LGC-soft、Saikai 231、LGC-Katsu和LGC-Jun等品质优良的低谷蛋白品种,并受到各大医院及肾病患者的普遍欢迎[11-13]。我国开展相关品种选育的时间较晚,但发展迅速。2002年,南京农业大学水稻研究所利用低谷蛋白品种LGC-1与日本优质粳稻越光杂交并以越光为轮回亲本进行回交,结合分子标记辅助选择育成了国内首个低谷蛋白新品种W3660[14]。此后,以LGC-1为直接或间接亲本,与籼、粳稻主栽品种五山丝苗、武育粳3号、秀水128、圣稻735等进行杂交和选育,创制了一批优质、高产的低谷蛋白新品系[15-18]。然而,这些材料在江苏、上海等长江中下游适宜地区进行种植时,在食味品质上还难以满足喜吃半糯香粳米人群的要求[19]。因此,迫切需要在育种中加强对功能性和食味品质方面的协同改良。
本研究以具有低直链淀粉含量基因Wxmp和香味基因fgr的优良食味粳稻品种南粳46为母本,与含低谷蛋白基因Lgc1的日本粳稻品种LGC-1杂交、回交,利用与目标基因共分离的分子标记进行跟踪检测。在此基础上,结合田间农艺、产量和品质性状的选择,选育具有优良食味和香味特征的低谷蛋白高产粳稻新品系。
1 材料与方法
1.1 供试材料
供试水稻材料包括含Lgc1基因的低谷蛋白粳稻品种LGC-1,含低直链淀粉含量基因Wxmp和香味基因fgr的晚粳稻品种南粳46,南粳46和LGC-1杂交、回交后代以及具有Lgc1Lgc1 WxmpWxmpfgrfgr纯合基因型的5个BC2F6新品系。所有材料在南京试验基地按常规栽培方式进行田间管理。
1.2 DNA提取和分子标记检测
水稻移栽后15 d,按单株剪取水稻叶片,用CTAB法提取基因组DNA[20]。低谷蛋白基因Lgc1检测所用插入/缺失标记为InDel-Lgc1-1(正向序列5'-TTCTACAATGAAGGCGATGC-3',反向序列5'-CTGGGCTTTAACGGGACT-3')和InDel-Lgc1-2(正向序列5'-ACCGTGTTATGGCAGTTT-3',反向序列5'-ATTCAAGGGCTATCGTCT-3')[21];低直链淀粉含量基因Wxmp检测所用标记序列为四引物扩增受阻突变体系PCR标记:正向外引物序列Wxmp-O-F为5'-ATGTTGTGTTCTTGTGTTCTTTG CAGGC-3', 反向外引物序列Wxmp-O-R为5'-GTA GATCTTCTCACCGGTCTTTCCCCAA-3',正向内引物序列Wxmp-I-F为5'-GGGTGAGGTTTTTCCAT TGCTACAATCG-3',反向内引物序列Wxmp-I-R为5'-GTCGATGAACACACGGTCGAC TCAAT-3'[22];香味基因fgr检测所用的插入/缺失标记为InDel-E2:正向引物序列5'-GGGAGGCGCTGAAGAGGA-3',反向引物序列5'-GGGTAGTCACCACCCTACC TTG-3'[23]。20 μL PCR体系包括DNA(10 ng/μL)2.0 μL,引物(4 pmol/μL)2.0 μL,10×缓冲液(含MgCl2)2.0 μL,dNTP(2.5 mmol/L)0.4 μL,Taq(5 U/ μL)0.5 μL,ddH2O 13.1 μL。反应程序如下:95℃下预变性5 min;95℃下变性30 s,(Lgc1基因标记56℃,Wxmp基因标记65℃,fgr基因标记55℃)复性30 s,72℃下延伸1 min,循环35次;72℃下延伸7 min,10℃冷却10 min。其中,Lgc1和Wxmp基因扩增产物加入DuRed核酸染料后在1.5%的琼脂糖凝胶电泳分离,而fgr基因扩增产物在8.0%的聚丙烯酰胺凝胶电泳分离和银染显色。
1.3 种子总蛋白提取、SDS-PAGE
种子总蛋白提取参照江绍玫等[24]的方法,略有改动。具体步骤:将单粒精米放入碾钵磨成极细的粉末,全部转入1.5 mL 离心管。加入15 μL β-巯基乙醇浸润20 min,再加入285 μL 65℃温浴的蛋白抽提液(5 mol/L脲,4% SDS,20% 甘油,100 mmol/L Tris-HCl,pH=6.8),充分涡旋数秒钟,室温静置过夜。电泳上样前10 000 r/min下离心10 min,取10 μL上清进行SDS-PAGE电泳(分离胶15%,浓缩胶7.5 %),电泳后用考马斯亮蓝G-250染色。
1.4 稻米总蛋白和各组分含量测定
成熟稻谷放在通风处2个月后,经垄谷机(SY88-TH,韩国双龙)、小型精米机(BLH-3120,台州伯利恒)去壳出精,用旋风式磨粉机(CT193,FOSS,瑞典)研磨成米粉,过100目筛去除大颗粒,获得用于测定的米粉。总蛋白含量测定采用全自动凯氏定氮仪(Kjeltec 8400,FOSS,丹麦)测定米粉中的全氮含量,再乘以换算系数5.95。蛋白质组分的提取和测定参照相关方法进行,略有改动[25-26]。取精米粉0.5 g,按顺序依次提取4种蛋白质,即清蛋白(10 mmol/L Tris-HCl,pH 7.5),球蛋白(1 mol/L NaCl,10 mmol/L Tris-HCl,pH 7.5),醇溶蛋白(体积分数为55%的正丙醇,10 mmol/L Tris-HCl,pH 7.5)和谷蛋白(质量分数为0.24% CuSO4·5H2O),1.68% KOH,0.5%酒石酸钾钠和体积分数为50%的异丙醇)。每次提取液用量为25 mL,室温下振荡2 h,其后于4000 ×g下离心10 min。清蛋白、球蛋白和醇溶蛋白的测定采用考马斯亮蓝法,谷蛋白的测定采用双缩脲法,分别计算出4种蛋白所占含量和比例,用凯氏定氮仪测定的总蛋白含量对各组分含量进行换算。每个样品测定3次,取平均值。
1.5 农艺、产量和品质性状测定
以亲本LGC-1和南粳46作对照,对新品系进行比较试验。每个品系(品种)重复3次,每重复小区栽种10行,每行28苗,株行距为13.2 cm×26.6 cm。各小区均分别记载播种期、抽穗期和收获日期。成熟后,在每个小区的第5或第6行中间连续取样10株,分别调查株高、单株穗数、单株实粒数、单株颖花数、每穗实粒数、每穗颖花数、结实率、千粒重和单株产量等性状。同时,测定每个小区实收产量和稻谷含水量,并按粳稻14.5%的标准水分折算小区产量。
糙米率、精米率、整精米率、粒长、粒宽、垩白粒率、垩白度、糊化温度、直链淀粉含量、胶稠度等性状的测定参照优质稻谷国家标准(GB/T 17891-1999),并利用米饭食味仪(STA-1A,日本佐竹)对食味值进行测定,每个样品测定3次,取平均值。
1.6 统计分析
按照莫惠栋介绍的方法[27],采用Excel 2016进行方差分析和多重比较,多重比较采用Duncan新复极差法。
2 结果与分析
2.1 分子标记辅助选择和新品系选育
2009年夏,在南京以南粳46为母本与LGC-1进行杂交,同年冬季在海南种植F1,选择真杂种作父本与南粳46进行回交,产生BC1F1种子。2010年夏,对21个BC1F1单株进行Lgc1、Wxmp和fgr基因进行检测,其中2个单株同时具有Lgc1lgc1WxmpWxmpfgrfgr基因型,并以其作父本分别与南粳46进行回交,产生BC2F1(图1)。2010年冬在海南,对2个BC2F1共25个单株进行检测,其中11株为Lgc1lgc1WxmpWxmpfgrfgr基因型,按单株收获种子。2011年夏,种植BC2F2株系,每个株系种植40株,在这些株系中选择农艺性状优、丰产性好的85个单株进行检测,筛选出19个Lgc1Lgc1WxmpWxmpfgrfgr基因型的单株。2012-2014年在南京分别种植BC2F3到BC2F5株系,并在农艺性状、丰产性好的株系中选择优良单株,根据外观品质性状进行淘汰,最终在BC2F6中筛选出目标基因型明确、综合性状优良、农艺性状稳定的5个新品系(图2~3)。
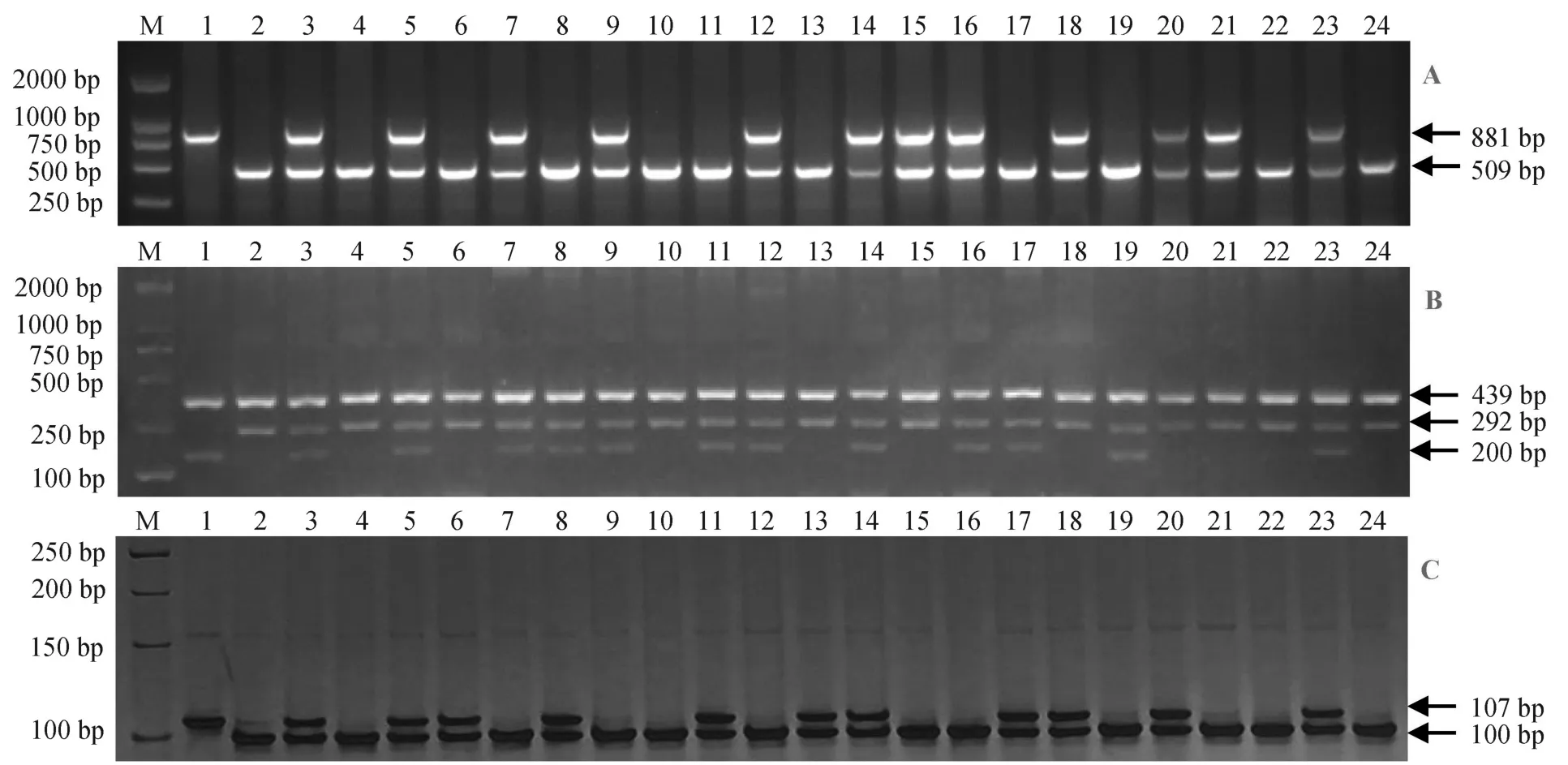
图1 BC1F1世代各单株Lgc1(A)、Wxmp(B)和fgr(C)基因型的电泳检测Fig. 1. Electrophoresis detection of Lgc1(A), Wxmp (B) and fgr (C) genotype for B1C1 plants.

图2 新品系Lgc1(A)、Wxmp(B)和fgr(C)基因型的电泳检测Fig. 2. Electrophoresis detection of Lgc1(A), Wxmp (B) and fgr (C) genotype for new lines.
2.2 种子总蛋白的SDS-PAGE分析
从水稻种子总蛋白的电泳谱带可以看出,与谷蛋白含量正常的品种南粳46相比,低谷蛋白品种LGC-1和5个新品系在成熟的谷蛋白α酸性亚基(37~39 kDa)和β碱性亚基(22~23 kDa)的条带变弱、含量较低,而醇溶蛋白(13 kDa)的条带增强、含量较高(图4)。同时,以南粳46为母本与LGC-1杂交获得的F1种子的蛋白条带与LGC-1完全相同,表明由Lgc1基因控制的低谷蛋白性状确实呈显性遗传,且不存在胚乳的剂量效应,这与前人的研究结论一致[9-10]。
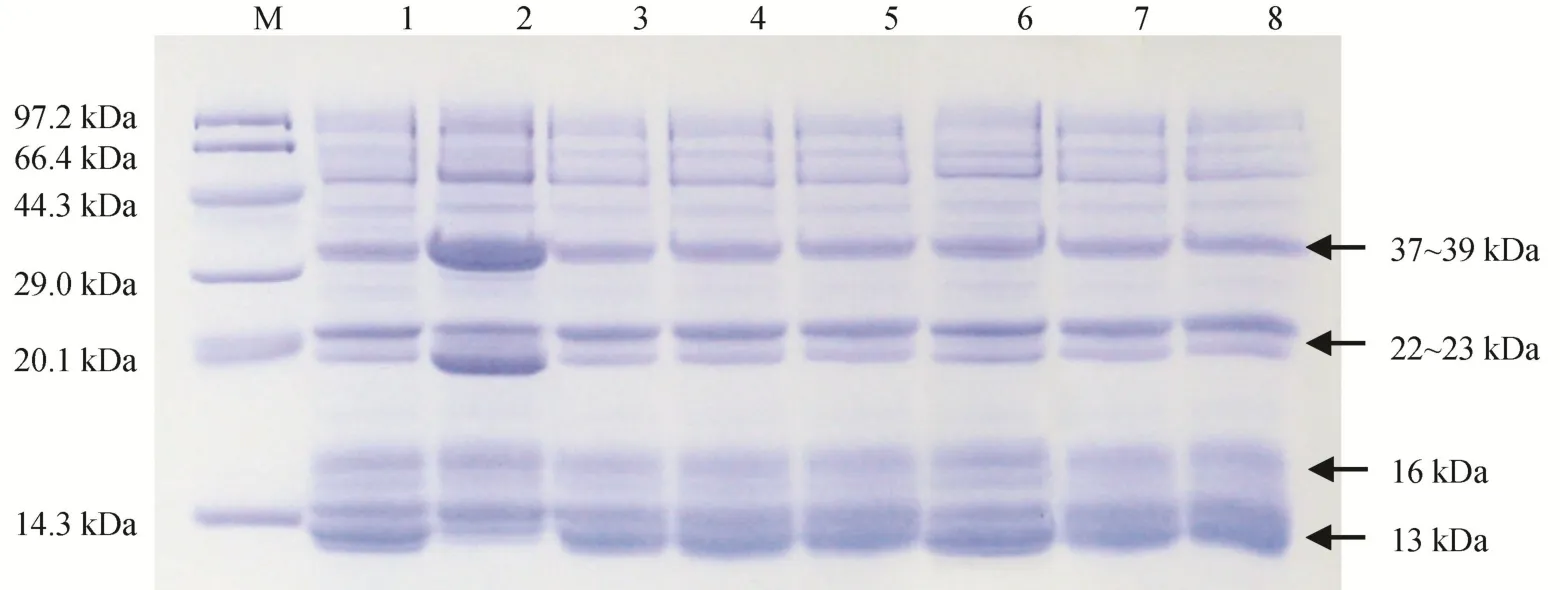
图4 低谷蛋白新品系种子总蛋白的SDS-PAGE分析Fig. 4. SDS-PAGE analysis of total protein in rice seeds for new lines of low glutelin content.
2.3 新品系稻米总蛋白及各组分含量的分析

图3 优良食味、低谷蛋白香粳稻新品系的选育过程Fig. 3. Process of development of new low glutelin content japonica rice lines with good eating quality and fragrance.
对5个新品系及其亲本的稻米总蛋白及各组分含量测定值的统计分析(表1)表明,L03、L07、L16、L19和L20的总蛋白含量为7.91%~8.15%,略高于南粳46,但品系间、亲本间均无显著性差异。新品系的谷蛋白含量为2.38~2.53%,醇溶蛋白的含量为4.30~4.47%,分别占总蛋白的比例为29.38~31.10%和54.11~55.37%,与LGC-1相近,而显著低于和高于亲本南粳46。在清蛋白、球蛋白的含量和比例上,新品系间和亲本间均无显著性差异。进一步对清蛋白、球蛋白、谷蛋白三种可吸收蛋白的总含量进行分析,5个新品系的可吸收蛋白含量为3.61%~3.74%,均显著低于南粳46,而与LGC-1无明显差异,符合低谷蛋白水稻%可吸收蛋白低于4%的指标要求。
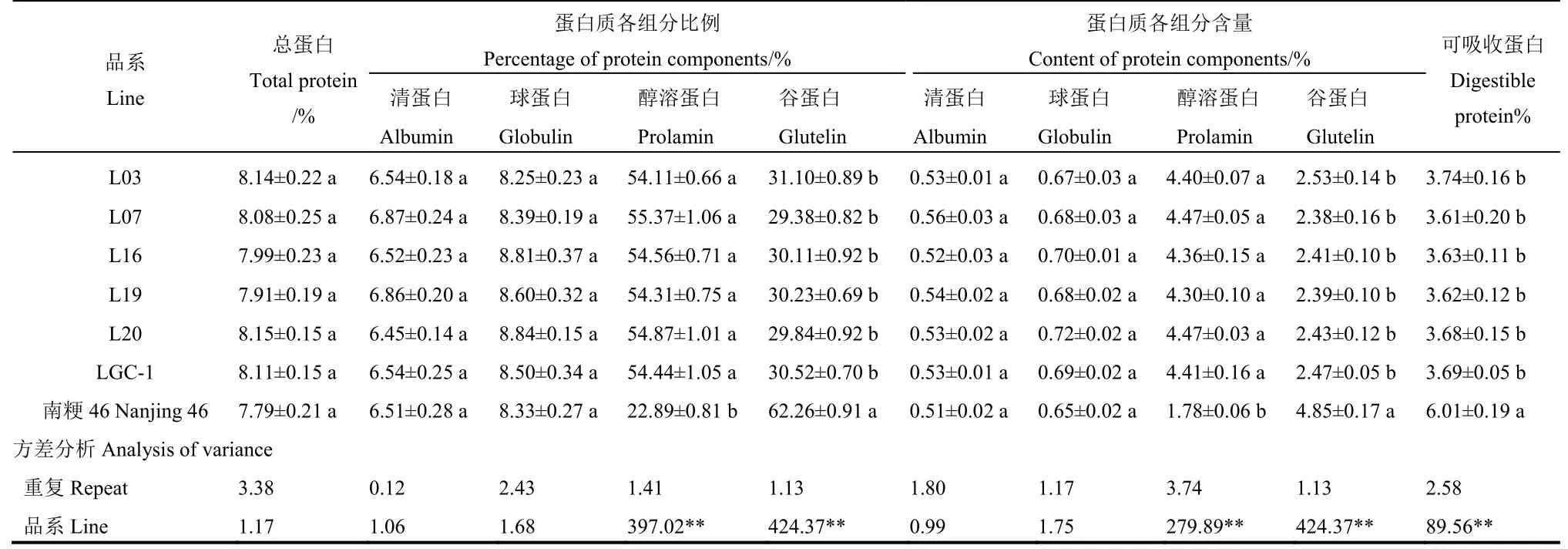
表1 新品系与亲本稻米总蛋白及各组分含量的比较Table 1. Comparison of total protein and its component contents in rice grains for new lines and parents.
2.4 新品系主要农艺、产量性状的分析
分析新品系及其亲本主要农艺和产量性状的差异发现,除结实率外,10个相关性状在不同品种(系)间的差异均达1%的显著水平(表2)。5个新品系的生育期比LGC-1长20.4~40.0 d,其中L03、L07和L16比南粳46早6.7~20.0 d,而L19和L20与南粳46相近。株高为94.2~107.8 cm,介于亲本LGC-1和南粳46之间。在产量性状方面,与LGC-1多穗、弯小穗的田间表现不同,5个新品系的单株穗数明显减少,每穗实粒数、每穗颖花数、单株实粒数、单株颖花数、千粒重、单株产量均显著提高,表现出亲本南粳46粗秆大穗、丰产性好的特征。与南粳46相比,L03和L07除单株穗数显著增加,结实率、千粒重无明显差异外,其他产量性状均显著降低;L16的每穗实粒数、每穗颖花数降低但穗数增加,因而单株实粒数、单株颖花数没有减少,同时千粒重的增加反而使其单株产量显著高于南粳46;L19和L20的产量性状则与南粳46相似。实际测产与产量性状的考种分析结果基本一致,其中L16的产量最高,L19和L20的产量与南粳46相近,L03和L07的产量虽低于南粳46,但均显著高于低谷蛋白品种LGC-1(表2,图5-A~F)。
2.5 新品系主要品质性状的分析
进一步对其主要品质性状进行方差分析,所测定的品质性状在品种(系)间的差异均达5%或1%的显著水平(表3)。多重比较的结果(表3)表明:亲本和新品系的糙米率、精米率和整精米率等碾磨品质性状较优,其中L03、L16、L19的3项指标值与高值亲本南粳46相近, L07、L20与低值亲本LGC-1差异不显著。外观品质性状中,新品系L16的粒长、粒宽均显著高于其他品系和双亲,这可能是其千粒重较大的主要原因,其他品系与亲本南粳46无明显差异,但粒宽都显著高于LGC-1;新品系的垩白粒率和垩白度均显著高于亲本LGC-1,个别品系如L03、L07、L16等的垩白粒率或垩白度高于高值亲本南粳46,但总体较优,都达到国标三级优质稻谷的标准(图5-G~L)。食味品质性状中,由于新品系具有与优良食味亲本南粳46相同的WxmpWxmp纯合基因型,其直链淀粉含量均在9.3%~9.8%范围,显著低于含WxbWxb纯合基因型的亲本LGC-1。同时,新品系直链淀粉含量的降低也导致其胶稠度升高。同时,食味值的测定结果也表明新品系的食味值均显著高于低谷蛋白亲本LGC-1,部分品系与南粳46接近。

图5 低谷蛋白新品系和LGC-1的田间表现及稻米外观Fig. 5. Field performance and appearance quality for new lines of low glutelin content and LGC-1.
3 讨论
作为我国的主要粮食作物,水稻对保障国家粮食安全和促进国民经济发展具有重要意义。长期以来,水稻改良的目标主要集中在高产、优质、抗病虫等重要农艺性状,并育成了一批具有重大应用价值的优良品种。近年来,随着社会经济的不断发展和生活水平的持续提升,饮食健康逐渐引起人们的关注。因此,加快功能稻米的相关研究和产业化,才能满足人们由“吃得饱”、“吃得安全”,向“吃得健康”转变,从而推动健康中国战略的深入实施。

功能稻米是一类区别于普通稻米的水稻功能性产品。它除具备能维持人体正常能量所需的基本营养成分外,同时其稻米皮层、胚和胚乳等含有某些对人体健康有益的生理活性物质或特殊成分较普通稻米高或低,通过食用可以平衡体内营养、预防疾病发生和促进疾病康复,且符合安全标准的一类稻米产品[28-29]。根据其功能性成分的类型,可分为功能性蛋白质型、活性多糖型、功能性油脂型、功能性维生素型、功能性黄酮类化合物型、必需微量元素型等[30],其中低谷蛋白稻米属于功能性蛋白质型。与普通稻米相比,其谷蛋白含量明显降低,可吸收蛋白含量低于4.0%。临床试验表明,利用低谷蛋白大米对慢性肾脏病患者进行饮食辅助治疗,能够减少蛋白摄入量,明显降低血清肌酐含量[31]。进一步研究发现,它能明显降低健康小鼠的尿蛋白水平,对肾脏具有一定的保护作用,且对小鼠的营养状况没有影响[32]。同时,通过在仿生大肠反应器中对健康人群的粪便微生物的发酵试验也证实低谷蛋白大米对肠道菌群的结构和代谢均具有正向影响[33]。因此,作为肾脏病患者一种经济、有效的食疗辅助品,低谷蛋白大米具有广阔的应用前景,而培育低谷蛋白新品种也成为功能性水稻育种的热点。
传统低谷蛋白品种的选育主要是根据种子总蛋白的SDS-PAGE电泳来推测其基因型并进行选择。由于控制低谷蛋白性状的Lgc1基因为显性遗传,要明确杂合或纯合基因型就必须对同一单株的多个籽粒进行检测,其操作过程繁琐、工作量大。同时,在实际育种中还需考虑对其他重要基因和综合性状的选择,仅通过传统育种方法很难育成具有多个目标性状基因的优良品种。分子标记辅助选择利用与目标性状基因紧密连锁的分子标记对基因型进行选择,且不受环境和等位基因显隐性、互作的影响,是实现水稻多基因聚合的一种重要手段,并在功能稻育种方面取得明显成效。张光恒等[34]将转葡萄糖焦磷酸酶(AGP)基因的水稻中超123的T5代纯系与富含γ-氨基丁酸的水稻巨胚1号杂交,通过分子标记辅助选择创建了高千粒重且具有降血压作用的功能稻新品系;Morita等[35]利用开发的分子标记对低谷蛋白品种LGC-1与来自越光的α-球蛋白缺失突变体的杂交后代进行辅助选择,创制出可吸收蛋白含量进一步降低的水稻材料;王萌等[36]以红米正常胚品系上师大6号和白米巨胚品种上师大5号进行杂交、回交,并结合红米基因Rc、Rd、巨胚基因gec和条纹叶枯病抗性基因qSTV11KAS的分子标记辅助育种,选育出兼具抗病性的红米巨胚功能稻新品系上师大10号。本研究中,为培育符合江苏、上海等长江中下游消费需求的优良食味、低谷蛋白香粳稻品种,我们选择被誉为“江苏最好吃大米”的江苏高产粳稻品种南粳46与日本低谷蛋白品种LGC-1进行配组,从而聚合关键的目标性状基因。为提高选择效率,在回交和自交的早期世代利用与低直链淀粉含量基因Wxmp、香味基因fgr和低谷蛋白基因Lgc1的功能性分子标记进行筛选,并结合常规育种的方法,选育出同时含有多个目标基因的粳稻新品系。与LGC-1相比,新品系的谷蛋白含量和可吸收蛋白含量相仿、食味品质得到显著提升。此外,在分子标记辅助选择时,我们也注重对其他重要性状的协同改良。分析结果表明,这些新品系不仅农艺性状良好,具有较高的增产潜力,而且其碾磨品质也得到明显改善。同时,新品系的生育期比亲本LGC-1长20.4~40.0 d,适宜在江苏中粳中熟、迟熟中粳和晚粳稻区进行选择性种植。
同时,研究中我们也发现杂交方式对提高品种选育的效率也非常重要。亲本LGC-1和南粳46虽同为粳稻,但地理来源不同,双亲遗传关系较远,F2世代的农艺性状会出现严重分离。若直接从该世代开始进行系谱法选择,不仅基因型检测的工作量大、成本高,而且很难筛选到农艺性状理想的单株。因此,本研究中我们将丰产性好的品种南粳46作轮回亲本回交两次再进行自交选择,既保证后代群体具有一定的遗传多样性和优良的农艺性状,而且也降低了田间种植和标记检测数量,显著提高了育种效率,这与目前利用日本品种LGC-1作亲本选育低谷蛋白水稻品种(系)的多数育种家的思路基本一致[14-15,18]。
由此可见,作为一种快速、准确、有效的育种手段,分子标记辅助选择目前仍不成熟、不完善,特别是在微效基因控制的数量性状选择上还不能完全替代常规育种的作用,两者只有在实践中紧密结合,才能高效、同步的改良产量、品质、抗性等综合性状,才能为水稻育种注入科技创新的活力。
