应用CRISPR/Cas9技术与分子标记辅助选择创制广东丝苗米新种质
2023-01-18王石光陆展华刘维卢东柏王晓飞方志强巫浩翔何秀英
王石光 陆展华 刘维 卢东柏 王晓飞 方志强 巫浩翔 何秀英
(广东省农业科学院 水稻研究所/广东省水稻育种新技术重点实验室/广东省水稻工程实验室, 广州 510640; *通信联系人, email: xyhe@163.com)
优质稻选育是水稻产业供给侧改革,提高水稻种植效益的关键举措。广东作为水稻起源地之一,率先开启了水稻优质化育种,在常规稻和杂交稻优质化育种过程中取得了显著成效[1]。紧随时代发展需求,广东水稻优质化育种一直走在全国前列,形成了走细小粒型的广东丝苗米特色之路。2018年广东开始实施丝苗米振兴战略,创立了广东丝苗米产业联盟,并制定了广东丝苗米品种标准和产品标准,其中香味与细小粒型是广东丝苗米品种的主要评价指标[1],极大地促进了广东高档优质稻米的发展。然而,当前优质丝苗米品种特别是高产、稳产、抗逆等综合性状优良的优质香型丝苗米水稻品种仍较为欠缺,亟需优化优质丝苗米品种结构。
水稻功能基因组研究成果为践行水稻分子设计育种、创制“绿色超级稻”提供了具有重要利用价值的基因资源[2]。研究表明,编码甜菜碱醛脱氢酶的基因Badh2功能性缺失,导致2-乙酰基-1-吡咯啉(2-AP)特异积累而使稻米产生香味[3,4]。在水稻粒型与稻米外观品质方面,大量相关的基因被相继克隆。其中,从粒型细长的优质不育系泰丰A中分离克隆的粒型新基因GW7表达量上调可增加谷粒纵向细胞分裂并减少横向细胞分裂,导致谷粒变得细长[5];其等位基因GL7由于一个17.1 kb的串联重复片段而使得其存在拷贝数变异(copy number variant, CNV),进而引起GL7表达量上升,同时下调了邻近的负向调节因子的表达量,从而增加水稻粒长并改善稻米外观品质,序列分析显示GL7位点的CNV在育种中受到选择并得以应用[6]。
在优质稻育种方面,本研究团队选育出了以粤晶丝苗2号、粤农丝苗、粤王丝苗、粤禾丝苗为代表的优质稻品种[7-9]。其中,粤农丝苗是丰抗优综合性状优良的优质常规稻品种,具有高产稳产性好、稻米品质优、高抗稻瘟病和中抗白叶枯病、抗倒力强且适应性广等特点,自2013年以来已连续十年被推荐为广东省农业主导品种[10-13]。优质稻粤王丝苗携带有GW7的优良等位基因,并表现为粒型细长、外观品质好等特点[7]。近年来,以CRISPR/Cas9为代表的基因编辑技术为特异种质的创制提供了有力的工具,为下一代育种战略奠定了基础,有望成为解决全球粮食短缺问题的重要方法[14]。本研究利用CRISPR/Cas9技术编辑香味基因Badh2定向改良高产稳产主栽品种粤农丝苗与优质稻粤王丝苗,成功创制两个香稻新品系。在此基础上,利用MAS技术定向导入GW7/GL7优良等位基因,成功选育两个丰产性好、籽粒长宽比超过4.3的优质香型丝苗米水稻新品系。这将为优质高产丝苗型香稻新品种(系)的创制与选育提供新的借鉴,对广东丝苗米新品种选育与丝苗米产业的发展具有重要意义。
1 材料与方法
1.1 试验材料与种植条件
本研究以广东省农业主导品种粤农丝苗和优质稻粤王丝苗为试验材料,利用CRISPR/Cas9技术进行基因编辑,在此基础上利用分子标记辅助选择进行系谱选育创制新种质。所有研究材料均种植于广东省农业科学院水稻研究所试验基地,大田育秧、单株移栽,株行距为16.7 cm×16.7 cm,采用常规栽培措施进行田间管理。
1.2 敲除载体的构建与无T-DNA元件的敲除株系的获得
香味基因Badh2的CRISPR/Cas9敲除载体构建与遗传转化均在百格基因科技有限公司进行,靶标设计采用该公司开发的高通量CRISPR-Cas9靶点设计程序。构建好的敲除载体利用农杆菌介导法分别转化至粤农丝苗与粤王丝苗中。利用包含Badh2靶点的特异引物对所有转化的T0代植株进行扩增测序,通过序列比对并结合测序峰图鉴定各T0代植株的突变方式。靶点扩增引物正反向序列分别为5'-CCTCCTCTTCACCCTCTCCA-3'和5'-AGGAT ACGCACTTCCGATGC-3'。利用Cas9与gRNA特异扩增引物进行PCR筛选T1及其后代是否存在T-DNA,以此获得无T-DNA元件的敲除纯合株系。Cas9检测引物正反向序列分别为5'-CACCA TCTACCACCTGAGAA-3'和5'-CGAAGTTGCTCT TGAAGTTG-3'。gRNA检测引物正反向序列分别为5'-CTGGCGAAAGGGGGATGTGCTGCAA-3'和5'-ACGACCGGGTCACGCTGCACCT-3'。
1.3 农艺性状调查与糙米粒型比较分析
在水稻灌浆充实期,调查待测株系中间一行的5个不同单株的株高;成熟期时,将每个待测株系中间生长正常的3个单株收割并自然晒干,用于有效穗数、穗长、穗粒数、千粒重、结实率、产量以及谷粒粒形与糙米粒形等性状的考查。其中,谷粒粒型与千粒重性状使用智能考种分析仪(TPKZ-3,托普云农)进行考种测定;糙米粒型性状使用米质分析仪(TS-M,托普云农)进行考种测定。将各性状的平均值作为该性状的表型值,使用Excel 2013进行数据整理,使用SPSS 22.0进行数据统计分析,根据最小显著差异法进行多重比较,具体考种标准与数据分析参照文献[15]进行。
1.4 外标法测定香味物质2-AP相对含量
待测株系稻谷收获晒干后磨成糙米放-20℃冰箱保存备用。待测糙米样品使用LUKYM-1冷冻研磨仪磨粉,称取2 g样品加入10 mL色谱纯二氯甲烷进行萃取并用过量无水硫酸钠去除水分,立即用无菌注射器吸取1 mL注入顶空样品瓶进行气相色谱质谱测定。GC-MS采用岛津GCMS-QP2020NX型气相色谱质谱联用仪和2-AP标准样品外标法标定香气相对含量。香味物质2-AP含量的测定委托广东省农业科学院稻米品质研究室完成。
1.5 GW7基因型的鉴定与分子标记辅助选择育种
采用Zhang等[16]开发的功能性标记GL7_Dup、GL7_InDel1与GL7_InDel2对试验材料的GW7/GL7基因型进行分子鉴定。利用上述功能标记进行分子标记辅助选择育种。GW7/GL7位点的分子检测从F1代开始贯穿于整个系谱选育过程,用以选育外观粒形细长的优质香型丝苗米稻新种质。本研究采用的PCR程序与参数设置以及电泳检测参照此前文献[17]进行。
2 结果与分析
2.1 粤农丝苗与粤王丝苗外观粒型的比较分析
广东丝苗米的典型特征是细小粒型,其关键的评价指标是糙米粒长≥6.5 mm且糙米长宽比≥3.5,千粒重≤21 g,或长宽比≥4.0的品种千粒重≤23 g[1]。我们以优质丝苗米品种象牙香占为对照,对试验材料的外观粒型与千粒重进行了比较分析(图1)。结果表明,粤农丝苗的糙米粒长超过6.90 mm,但其粒型最短且宽,其糙米长宽比只有3.37±0.05,谷粒千粒重达到24.66±0.30 g,均未达到广东丝苗米品种标准。与之对应的是,粤王丝苗粒型最为细长,其糙米粒长达到8.40±0.19 mm,糙米长宽比则达到4.70±0.11,而千粒重为22.51±0.42 g,均符合广东丝苗米品种标准。
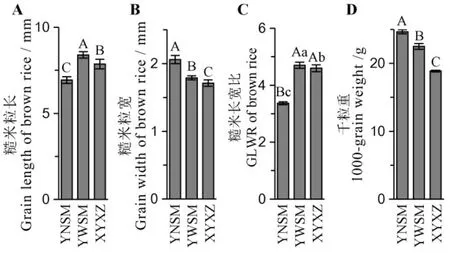
图1 亲本糙米粒型比较分析Fig. 1. Comparative analysis of grain size of parental brown rice.
2.2 CRISPR/Cas9定点敲除香味基因Badh2
为创制新的香稻种质,我们在香味基因Badh2的第2外显子处设计了一个靶点(图2-A),成功构建了其CRISPR/Cas9敲除载体。利用农杆菌介导法分别转化至粤农丝苗与粤王丝苗,分别获得26株与12株T0代转基因阳性植株。基于PCR扩增测序对上述阳性植株进行靶点变异分析,筛选粤农丝苗与粤王丝苗两个背景下香味基因badh2突变纯合单株。测序结果比对分析发现,粤农丝苗背景存在两种突变方式:一种为T碱基的插入,另一种为G碱基的插入;粤王丝苗背景存在一种突变方式:T碱基的插入(图2-B、C)。以上突变方式造成Badh2基因移码突变或形成终止密码子而导致翻译提前终止(图2-B)。随后,从T1代开始连续多代利用Cas9与gRNA特异检测引物对上述敲除株系进行检测,成功筛选到无T-DNA插入的纯合突变株系(图2-D),分别暂定名为yn-kobadh2与yw-kobadh2。

图2 Badh2基因敲除植株突变位点分析Fig. 2. Mutation types of Badh2 in gene knockout plants.
2.3 主要农艺性状比较与香味物质2-AP含量测定
为了检测Badh2基因敲除对主要农艺性状的影响,我们对yn-kobadh2与yw-kobadh2敲除株系的株高和产量等相关性状进行了比较与测定(表1)。与各自野生型对照相比,yn-kobadh2-1的株高显著降低,而穗长和谷粒粒长则显著或极显著增加;yw-kobadh2-1的株高显著降低,yw-kobadh2-2的谷粒长宽比显著增加。而在糙米外观粒型、有效穗数、每穗实粒数、千粒重与产量方面,yn-kobadh2与yw-kobadh2敲除株系与各自野生型对照均无显著差异(图3-B~F)。随后,我们对yn-kobadh2与yw-kobadh2敲除株系的稻米香味物质2-AP含量进行了测定(图3-G)。结果表明,相比于野生型亲本稻米中未检测到香味物质2-AP,yn-kobadh2与yw-kobadh2敲除纯合株系的2-AP含量均极显著提高,达到239.39~440.79 μg/kg。因此,可以利用CRISPR/Cas9基因编辑技术对Badh2进行定点敲除快速地创制香稻新种质。

图3 Badh2基因编辑香稻新种质的农艺性状比较分析Fig. 3. Agronomic traits of new fragrant rice germplasms through Badh2 gene editing.
2.4 利用MAS创制丝苗型香稻新品系
综合比较粤农丝苗与粤王丝苗发现,粤农丝苗的产量显著高于粤王丝苗,而粤王丝苗的外观粒型更为细长(表1和图4)。利用功能标记检测发现,粤王丝苗携带优良的GW7/GL7等位位点(图4-C)。为创制优质高产丝苗型香稻新种质,以yn-kobadh2-1与yw-kobadh2-1为亲本杂交,利用GW7/GL7位点的功能标记进行分子标记辅助选择。经过连续多代的分子检测与系谱选育,于F4代选育出两个优良株系,暂命名为NWbadh2GW7-1与NWbadh2GW7-2。农艺性状比较分析结果表明,NWbadh2GW7-1的产量与粤农丝苗相当,而NWbadh2GW7-2的产量达到7708.4 kg/hm2,极显著高于粤农丝苗,增加约23.2%;外观粒型方面,NWbadh2GW7-1和NWbadh2GW7-2与粤王丝苗相当,表现为细长粒型,其谷粒长宽比与糙米长宽比均超过4.3,极显著大于粤农丝苗(图4-D~G)。香味物质2-AP含量测定结果表明,NWbadh2GW7-1和NWbadh2GW7-2株系的2-AP含量均超过400 μg/kg(表1)。综上表明,NWbadh2GW7-1和NWbadh2GW7-2株系丰产性好且达到广东丝苗米品种关于香味与外观粒型的认定标准。
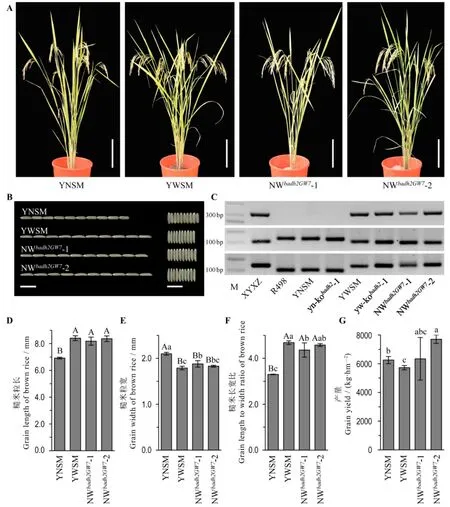
图4 丝苗型香稻新株系的农艺性状比较分析Fig. 4. Agronomic traits of new fragrant Simiao rice lines.
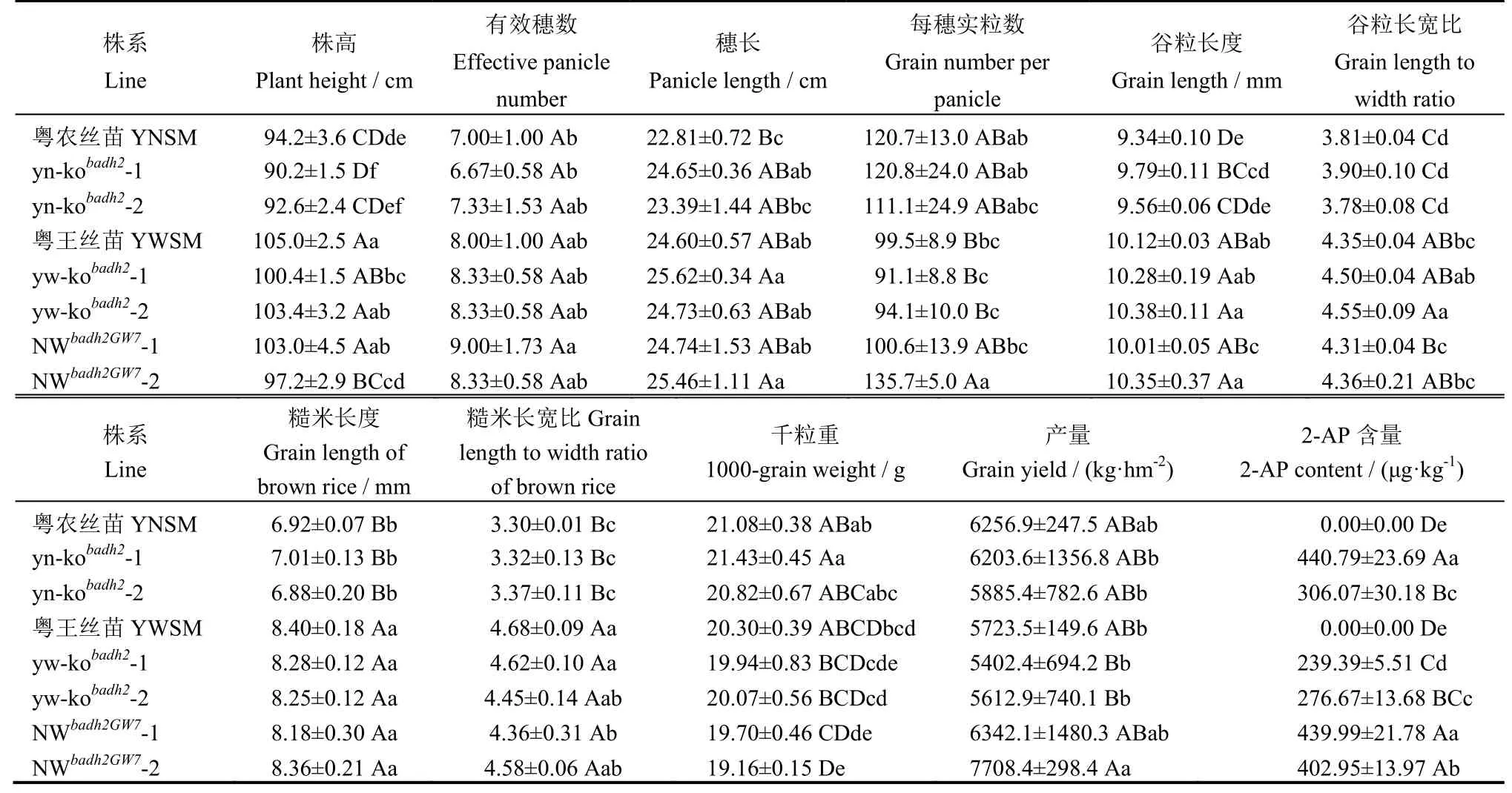
表1 野生型与创制株系的农艺性状Table 1. Agronomic traits of the wild type and generated lines.
3 讨论
水稻外观品质特别是粒型以及香味是影响稻米品质的重要性状。广东在水稻优质化育种进程中形成了走细小粒型的广东丝苗米特色之路,并制定了广东丝苗米品种标准和产品标准,其外观品质的主要评价指标就是细小粒型且要有香味[1]。其中,香味作为丝苗米必备的品质性状,赋予了丝苗米更高的稻米价格与种植效益。在水稻中,Badh2编码一个甜菜碱醛脱氢酶(betaine aldehyde dehydrogenase 2, Badh2),是控制稻米香味的主要基因,其功能缺失可导香味物质2-AP含量的大量积累,使稻米产生香味[3,4]。相比于传统的香稻育种存在的费时费力且效率低下的缺点,现已有大量的研究通过生物技术的手段对香味基因Badh2进行编辑以加速香稻育种进程,并创制出了大量新的Badh2突变等位基因[18-26]。本研究利用CRISPR/Cas9基因编辑技术定点编辑优质稻粤农丝苗与粤王丝苗的Badh2基因,成功创制两份香稻新品系yn-kobadh2与yw-kobadh2,两者2-AP含量均极显著提高,达到239.39~440.79 μg/kg(图3)。前人的研究结果表明,Badh2不同类型的突变方式对2-AP的生物合成与积累的影响可能不同[26-27]。本研究中,yn-kobadh2-1与yw-kobadh2及其两者衍生的NWbadh2GW7株系的Badh2基因均为同一种的突变方式,但yn-kobadh2-1与NWbadh2GW7株系的糙米中2-AP含量均超过400 μg/kg,极显著地高于yw-kobadh2的含量(表1)。以上结果表明,Badh2的突变可能会因不同品种之间遗传背景的差异而影响2-AP的合成与积累。此外,本研究中Badh2基因的敲除对有效穗数、每穗实粒数、糙米外观品质、千粒重与产量等主要农艺性状无显著影响(表1)。因此,定点编辑非香型优良水稻品种的Badh2基因可为香稻育种与改良提供新的优良种质资源。
前期,本研究团队利用Xa23、Pi2与Wx等作为主要抗源或目标基因,通过分子标记辅助选择(MAS)育成了粤恢88、粤新银占2号、粤特油占等优质抗病恢复系或常规稻新品种与台早1S、台S、沃S等籼型优质两系不育系[17,28,29]。本研究发现,粤王丝苗优异的外观粒型可能是由于其携带优良的GW7/GL7等位位点所致(表1和图4)。因此,在yn-kobadh2与yw-kobadh2香稻新种质创制的基础上,我们进一步通过MAS技术定向导入GW7/GL7位点,结合系谱选育的方法成功创制两个优质高产丝苗型香稻新株系NWbadh2GW7-1与NWbadh2GW7-2(表1和图4)。结果表明,NWbadh2GW7-1与NWbadh2GW7-2综合了粤农丝苗高产与粤王丝苗优良外观品质的优点,可为广东丝苗米品种选育提供重要的种质储备与遗传改良资源。因此,MAS选择技术作为重要的育种手段,可快速地聚合和转移有利基因,缩短育种年限,提高育种效率。然而,具有育种利用价值的基因特别是可用于MAS的有利基因有限,以及目标基因与不利性状的连锁等限制了MAS技术在水稻育种中的应用。相比而言,CRISPR/Cas9等基因编辑技术不仅可应用于定向敲除不良目标基因,也能用于整合优良目标基因或去驯化,可极大地提高水稻的遗传改良效率[30-32]。随着我国在水稻功能基因组学上的不断突破与生物信息学的快速发展,基因编辑技术、MAS选择技术与常规育种技术等相结合,将极大地推动分子设计育种与全基因组选择育种,以更精准、高效地选育高产、优质、抗逆、营养高效利用的“绿色超级稻”。
由于水稻香味基因Badh2为隐性基因,杂合型植株的香味性状发生分离,故只需杂交水稻的亲本之一携带有功能丧失的badh2等位基因,其配组的杂交组合的部分籽粒表现为香,如桃优香占等[26,33]。本研究中使用的粤农丝苗既是一个综合性状优异的常规优质稻品种,同时作为育种“芯片”被广泛应用于优质常规稻育种与杂交稻配组,以其为恢复系育成并通过国家、省级审定或授权的杂交稻组合已超过40个[12];优质稻粤王丝苗也可作为恢复系应用于杂交稻配组,如已通过国家审定的爽两优粤王丝苗。因此,本研究创制的香稻新品系yn-kobadh2与yw-kobadh2与丝苗型香稻新株系NWbadh2GW7可作为重要的遗传资源用于优质香型杂交稻育种。
总的说来,本研究利用CRISPR/Cas9基因编辑技术成功创制了两个香稻新品系yn-kobadh2与yw-kobadh2,在此基础上结合MAS技术定向选择GW7/GL7位点,选育出丰产性好且籽粒长宽比超过4.3的优质丝苗香型水稻新品系NWbadh2GW7,既为后续优质丝苗香型水稻品种的选育提供新的种质,也为后续水稻精准、高效育种改良提供新的思路。
