西伯利亚杏花芽分化及开花生物学特性
2023-01-12于庆福王蓬锴李家豪王新鑫董胜君
张 剑,于庆福,王蓬锴,李家豪,王新鑫,董胜君
(1. 沈阳农业大学 林学院,辽宁 沈阳 110866;2. 辽宁省喀左县林业种苗管理站,辽宁 喀左 122300)
西伯利亚杏Armeniaca sibirica(L.)Lam.是蔷薇科杏属植物,是亚洲特有的生态经济型树种[1]。西伯利亚杏具有较强的防风固沙和水土保持能力[2],对生态环境较为脆弱的干旱、半干旱地区具有重要作用[3],且其杏仁含有丰富的苦杏仁甙,是世界公认的能防癌、抗癌的天然活性物质[4]。
果树的花芽分化是其营养生长过渡为生殖生长的标志[5],花芽分化期主要包括两个时期:一是生理分化期,即多年生木本植物完成营养物质积累后的茎尖分生组织经诱导发育为花序分生组织并为形态分化提供物质基础的过程,此阶段其形态并无明显变化,因此也可称作未分化期;二是形态分化期,此阶段花器官原基陆续分化、生长为花萼、花瓣、雄蕊、雌蕊,最终发育为完整的花器官[6-7]。花芽分化期是植物生长发育过程中至关重要的阶段,花芽分化是植物生殖发育的基础[8]。目前关于杏属植物花芽分化的研究报道较多:刘少华等[9]对银香白花芽分化进程的研究结果表明,其花芽分化始于7月中旬,至9月下旬分化完成,历时约70 d;Distefan等[10]认为,花芽分化进程的快慢对植物开花时间的早晚具有重要影响;孙浩元等[11]的研究结果表明,开花较早可能会遭受晚霜危害,严重影响其产量。杏属植物开花受气候、地理条件的影响较大[12-13],其花期一般集中为3月下旬—4月上旬,且受生长素、赤霉素等多种激素的调控[14-15],故其不完全花比例较高且不同品种间的差异较大。为给西伯利亚杏属植物花期的合理调控提供技术指导,以不同种源的西伯利亚杏无性系为研究对象,采用石蜡切片技术对其花芽形态分化进行解剖学观察,分析花芽分化规律及不同无性系间的差异情况,监测花期物候进程,研究花器官开花生物学特性,测定其生长素含量,现将研究结果分析报道如下。
1 材料和方法
1.1 试验地概况
试验地为辽宁省喀左县,地处辽西半干旱地区,东经 119°51′26″,北纬 41°02′59″,海拔 300 ~400 m,该地呈低山丘陵地貌,土壤以棕壤为主。该地属温带大陆性季风气候区,年均气温8.7 ℃,植物生长期(4—9月)的降水量约为451.6 mm,占全年降水总量的92%。年均日照时数为2 808 h,无霜期144 d。该地区的主要木本植物有西伯利亚杏、酸枣、刺槐、山楂、油松等。
1.2 试验材料
供试材料来源于沈阳农业大学山杏国家林木种质资源保存库,分别选取种源与开花特点均不同的西伯利亚杏8号(辽宁省朝阳市)、28号(内蒙古赤峰市)、595号(吉林省吉林市)、622号(山西省阳泉市)、626号(山西省阳泉市)无性系,其树龄为6~7年,株行距均为2 m×3 m,长势良好。
1.3 试验方法
1.3.1 花芽采集
于2020年7月5日—10月26日,每隔7~10 d采样1次,共采样15批次,每次的采样时间均为8:00—10:00时,每批次在每个无性系树冠外围各随机采集饱满的花芽30个,用FAA固定液(50%乙醇∶冰醋酸∶福尔马林=90∶5∶5)固定,保存。
1.3.2 石蜡切片的制作
参考常规石蜡切片法并加以改进,将固定液中的花芽取出,放入盛有50%乙醇的培养皿中,用镊子剥去鳞片,之后按照脱水—透明—浸蜡—包埋—切片—贴片—展片—脱蜡—复水—染色—脱水—透明—封片—晾干的步骤制作切片,并在Olympus BX51电子体视显微镜下进行镜检、拍照。采用连续纵切法切片,选取花芽最中间的切片进行拍照保存,将每批次采集的每个无性系的花芽样品各制作成3张切片。
1.3.3 开花物候期的观测与划分
于2021年的3—4月对5个西伯利亚杏无性系的开花物候期分别进行观测,并参考辣椒杏[15]、山杏[16]物候期的划分标准,将其开花物候期划分为花芽萌动期(5%的花芽开始膨大)、露红期(5%的花萼露出)、露白期(5%的花瓣露出)、始花期(5%的花瓣完全开放)、盛花期(25%的花瓣完全开放)、末花期(75%的花瓣变色及开始脱落)共5个时期。
1.3.4 花期的调查
于2021年的盛花期,每个无性系各随机选择10朵完全盛开的花,观察其花瓣、花萼、花药、花丝颜色、花瓣褶皱、雌蕊的发育程度(仅需目测比对雌、雄蕊的长度即可),并使用数显卡尺测定花径(cm),同时计数雌蕊、雄蕊、花瓣的数量。
1.3.5 完全花比例的调查
于2021年的盛花期,每个无性系各选取3株样木,作为3次重复,分别调查每株样木的花束状果枝(<5 cm)、短果枝(5~15 cm)、中果枝(15~30 cm)、长果枝(>30 cm)这4类果枝上的完全花,每株样木每类果枝各调查25朵花,计算完全花(雌蕊长于或等于雄蕊的为完全花,低于雄蕊或者退化的为不完全花)的比例。
1.3.6 生长素的测定
于2021年的盛花期,分别采集28号、595号无性系完全盛开的花,充分研磨后加入内标混合液,经涡旋—离心—浓缩—复溶后,过0.22 μm的滤膜,置于进样瓶中,采用高效液相色谱-质谱联用仪(QTRAP 6500+,SCIEX)测定生长素类激素的含量,每个无性系的测定各设3次生物学重复。
1.4 数据处理
使用Excel 2021软件对观测到的花器官数据进行整理与统计,计算完全花的比例;采用SPSS 22.0软件对不同无性系样木不同类型果枝上的完全花比例进行多重比较。
2 结果与分析
2.1 西伯利亚杏花芽分化进程的解剖学观察
6个西伯利亚杏无性系的花芽分化进程均可分为未分化期、花原基分化期、萼片分化期、花瓣分化期、雄蕊分化期、雌蕊分化期共6个时期,对其花芽在此6个时期的石蜡切片进行观察,结果如图1所示,西伯利亚杏花芽在此6个时期的分化特征描述如下。
1)未分化期的花芽分化特征:芽内生长锥的宽大于高,锥尖呈拱形;生长锥旁边有大量逐次向外排列的、不同发育阶段的叶原基,最外层被鳞片紧紧包围(图1a)。
2)花原基分化期的花芽分化特征:前期,与未分化期的花芽相比,芽内生长锥体积增大且向上微微隆起,纵切面呈半圆形,生长锥旁边的叶原基较未分化期的少(图1b);后期,生长锥伸长,顶部变宽且趋于平缓,纵切面呈近似等腰梯形(图1c)。
3)萼片分化期的分化特征:生长锥顶部周围形成大小不一的明显突起物,纵切面上表现为生长锥两端突起,这个突起物就是萼片原基,同时,叶原基消失(图1d)。
4)花瓣分化期的花芽分化特征:花芽内萼片原基向上伸长并向内弯曲,在弯曲的内侧底部分化出多个小突起,即花瓣原基,表明花芽已经进入花瓣分化期(图1e)。
5)雄蕊分化期的花芽分化特征:前期,花芽内花萼原基继续向上生长并且向内弯曲,同时内部出现大量突起物,即雄蕊原基(图1f);中期,雄蕊原基向两侧延伸,顶端不断膨大,形成孢原细胞,纵切面呈椭圆形(图1g—h);到后期,雄蕊原基不断膨大,孢原细胞平周分裂形成造孢细胞和周缘细胞(图1i—j)。
6)雌蕊分化期的花芽分化特征:前期,花芽在不断分化生长,内卷的花萼原基张开,在生长锥底部平坦的中心产生很小的突起,即组成雌蕊的心皮原基(图1g);中期,雌蕊心皮原基在底部继续由侧边向中间卷合,并向上伸长形成花柱(图1i);后期,花柱继续向上生长,同时花柱底部膨大,中心出现空室,即子房(图1j)。

图1 西伯利亚杏花芽分化的解剖学观察结果Fig. 1 Anatomical findings of flower bud differentiation process in A. sibirica
2.2 西伯利亚杏的花芽分化进程
5个西伯利亚杏无性系花芽在其6个分化时期的分化进程见表1。由表1可知,5个西伯利亚杏无性系花芽分化时间为7月中旬—10月中旬,各个无性系花芽的分化由快至慢依次为:622号<28号=626号<8号<595号。其中,8号、28号无性系花芽开始分化的时间均最早(7月12日);595号无性系的次之(7月21日);622号、626号无性系的最晚,分别为8月11日、8月18日,较8号、28号的晚30 d左右。28号于7月21日最早进入萼片分化期,8号、595号均于7月28日进入萼片分化期,而622号、626号均于8月中下旬进入萼片分化期。进入花瓣分化期的时间,28号无性系依旧最早,其余各个无性系也均于8月进入花瓣分化期;且各个无性系花芽进入萼片分化期与其进入花瓣分化期的时间约有7 d重叠。各个无性系进入雄蕊分化期的时间出现了较大差异,28号最早(8月4日),8号、595号均稍晚(8月18日),622号、626号均最晚(9月1日),按花芽进入雄蕊分化期的具体时间可将5个无性系的雄蕊分化期分为3个时间段,每个时间段之间相差14 d,而各个无性系雄蕊分化结束的时间却略有不同。各个无性系花芽在雌蕊分化期的表现与其在雄蕊分化期的表现类似,即28号最早(8月11日),8号、595号(8月25日)均稍晚,622号(9月1日)较晚、626号(9月9日)最晚;同时,这一时期与雄蕊分化期的时间重叠度较高,重叠时间为15~36 d,各无性系的重叠时间平均为25 d。此外,28号无性系花芽分化完成的时间最早(9月9日),而626号花芽分化完成的时间最晚(10月16日)。
5个西伯利亚杏无性系在不同花芽分化时期的持续时间如图2所示。由图2可知,8号、28号、595号的花原基分化期、萼片分化期、花瓣分化期均持续15~17 d,而622号、626号在此3个时期的持续时间均为8 d。各个无性系雄蕊分化期、雌蕊分化期的持续时间均长于其在前3个时期的持续时间,且其雌蕊分化期的持续时间最长。其中,8号雄蕊分化期、雌蕊分化期的持续时间均最短,均约20 d;28号、622号、626号雄蕊分化期、雌蕊分化期的持续时间均约30 d;而595号于此2个时期的持续时间均最长,均约40 d。
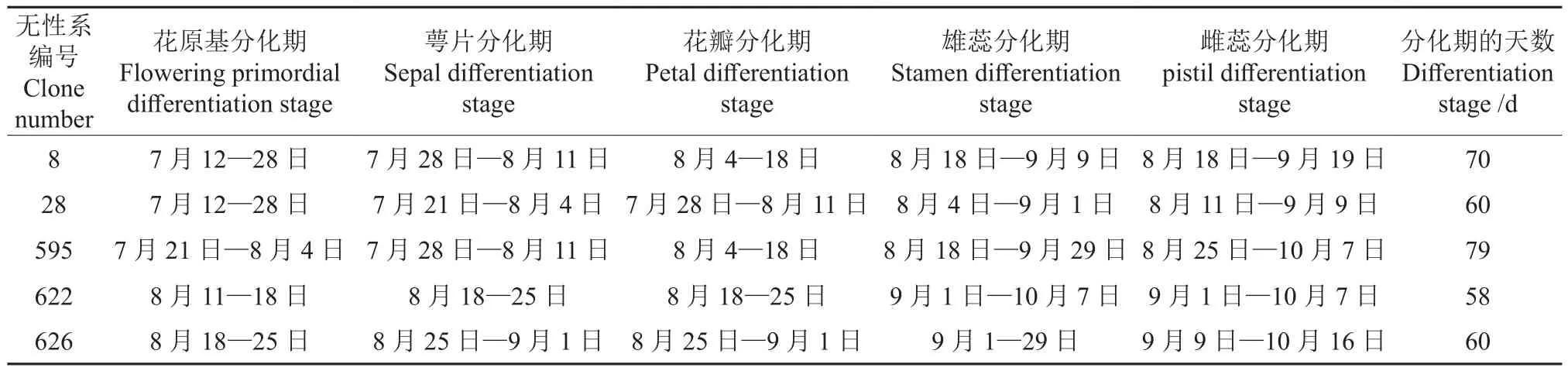
表1 西伯利亚杏花芽分化进程Table 1 Process of bud differentiation in A. sibirica

图2 5个西伯利亚杏无性系在不同花芽分化时期的持续时间Fig. 2 Duration of five A. sibirica clones at different periods of bud differentiation
由表1与图2可知,28号无性系的花芽分化较早,且其在各个分化时期的分化进程均较快,因此其分化期(59 d)较短;622号、626号无性系进入花原基分化期的时间均较晚,而其在各个分化时期的分化进程也都较快,因此其分化期与28号的均基本一致;但是,595号无性系的分化进程与28号、622号、626号的分化进程均相反,595号无性系进入花原基分化期的时间稍晚,而其雄蕊分化期、雌蕊分化期的持续时间均最长,因此595号花芽分化期的持续时间(79 d)最长。
2.3 西伯利亚杏的开花物候期
5个西伯利亚杏无性系的开花物候期见表2。由表2可知,5个西伯利亚杏无性系的花芽均在3月中旬发生萌动而进入花芽膨大期,3月下旬其鳞片裂开,花萼顶端露出,进入露红期,3月末萼片顶端开张,可见花瓣,到达露白期,4月初花朵开放,各个无性系由始花期进入盛花期的时间均较短,一般为2~4 d,而其盛花期的持续时间为4~5 d,4月中旬其花朵全部脱落,花期结束,此期的持续时间为9~11 d。各个无性系进入各个物候期的时间及其持续时间均仅相差1~3 d,其中,28号、595号的均较早,8号、622号、626号的均较晚。
2.4 西伯利亚杏花器官特征
西伯利亚杏各无性系花器官均表现为花瓣白色、不褶皱,花萼红褐色,花丝白色,花药黄色(表3),除28号无性系雌蕊高于雄蕊外,其余均为雌蕊低于雄蕊。数量性状方面,各无性系每朵花均具有1个雌蕊;除622号无性系花瓣出现变异(为六瓣花)外,其余无性系花瓣均为五瓣花;595号无性系的花径(2.19 cm)极显著小于其他无性系的,而其他无性系间其花径无显著性差异;各个无性系之间雄蕊数量的差异较小,其雄蕊数由多至少的顺序为:622号(32个)>8号(31个)>626号(30个)>595号(29个)>28号(28个)。
2.5 西伯利亚杏完全花的比例
5个西伯利亚杏无性系不同类型果枝的完全花比例的统计结果见表4。由表4可知,5个西伯利亚杏无性系各种类型果枝的完全花比例总均值为33.16%,不同无性系间完全花比例的差异较大。无性系的完全花比例,28号(99.42%)极显著高于其他无性系,是626号(28.59%)的3.44倍;626号(20.33%)极显著高于622号(20.33%)、8号(13.99%)、595号(3.50%),622号显著高于8号、极显著高于595号,而且8号极显著高于595号。

表2 5个西伯利亚杏无性系的开花物候期Table 2 Flowering phenology of five A. sibirica clones

表3 5个西伯利亚杏无性系的花器官特征†Table 3 Flower organ characteristics of five A. sibirica clones
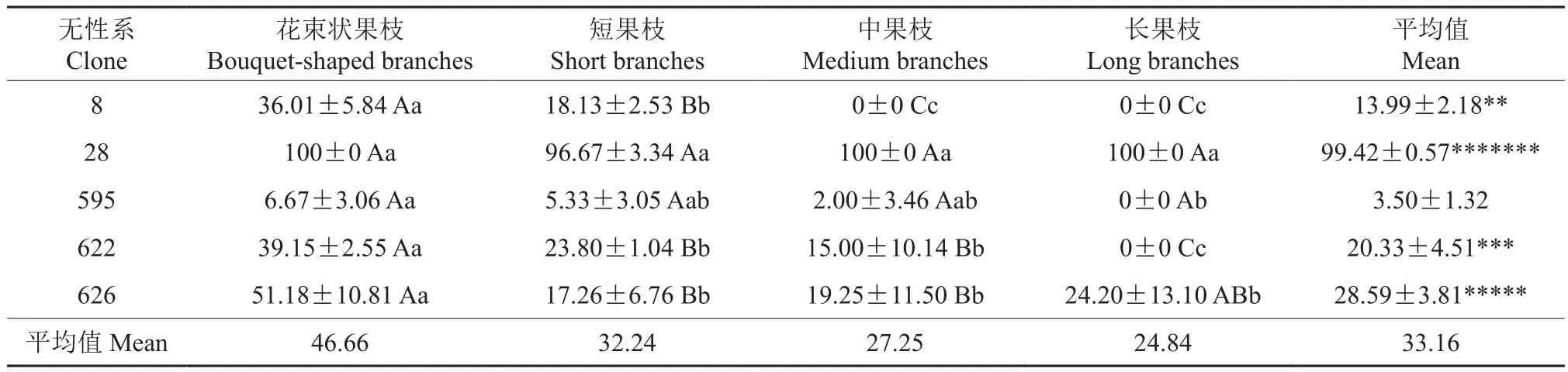
表4 5个西伯利亚杏无性系不同类型果枝的完全花比例†Table 4 The complete flower ratio in different fruiting branches of five A. sibirica clones %
由表4还可知,西伯利亚杏各类果枝的完全花比例由高到低依次为:花束状果枝(46.66%)、短果枝(32.24%)、中果枝(27.25%)、长果枝(24.84%)。各个无性系完全花比例最高的果枝均为花束状果枝,其中8号、622号、626号花束状果枝的完全花比例均极显著高于其自身其他果枝类型的,595号花束状果枝的完全花比例显著高于其自身长果枝的。8号、595号、622号中果枝、长果枝的完全花比例均低于28号、626号的,其中8号中果枝、长果枝的完全花比例均为0,595号、622号长果枝的完全花比例也均为0。28号各类果枝的完全花比例均最高,且其花束状果枝、短果枝、长果枝的完全花比例均达到100%,其各类果枝间完全花比例无显著性差异。
2.6 西伯利亚杏花内各种生长素的含量
28号与595号西伯利亚杏无性系盛花期花内的生长素含量见表5。由表5可知,在西伯利亚杏的盛花期,28号无性系花内吲哚-3-乙酸(IAA)、吲哚乙酸-天冬氨酸(IAA-Asp)和生长素(auxin)的总含量分别为215.97、5.36与226.16 ng/g,595号无性系花内IAA、IAA-Asp和生长素的总含量分别为77.59、3.67与87.45 ng/g,28号无性系的均极显著高于595号无性系的,而28号与595号无性系间其N-(3-吲哚乙酰基)-L-丙氨酸(IAA-Ala)、吲哚-3-乙酸甲酯(MEIAA)含量均无显著差异,28号无性系的吲哚乙酸-甘氨酸(IAA-Gly)含量显著低于595号无性系的,且28号无性系的IAA含量是595号IAA含量的2.78倍。

表5 28号与595号西伯利亚杏无性系盛花期花内的生长素含量†Table 5 The content of Auxin in A. sibirica flower at the full blooming stage ng/g
3 讨论与结论
3.1 讨 论
花芽分化是一个极为复杂的过程,生理分化完成后,植株受体内激素调控和外界条件刺激,逐渐分化出花芽和叶芽,进入形态分化期,各花器官逐步分化、生长,最终发育成为一个完整的花器官[8]。目前对植物花芽分化进程的划分并没有确切的标准,但是,前人一般按照花器官由外至内、按其形成时间由早及晚的顺序,将花芽的形态分化过程分为未分化期、花原基分化期、萼片分化期、花瓣分化期、雄蕊分化期、雌蕊分化期这6个时期[17],虽然有部分研究报道对某一分化阶段的称谓可能有所不同,但是对某一分化阶段的分化表现与特征的描述基本一致。研究结果表明,供试的5个西伯利亚杏无性系花芽形态分化过程也可分为上述的6个时期,同时参考孙建云等[18]对李花芽分化进程的划分标准,将西伯利亚杏花原基分化期又分为分化初前期、分化初后期两个时期。此外,为了详尽描述其花芽雄蕊、雌蕊的分化特征,本研究还根据雄蕊、雌蕊形态的分化特点将雄蕊分化期和雌蕊分化期均分为前期、中期、后期这3个时期。
研究结果表明,5个西伯利亚杏无性系花芽形态分化的时间为7月中旬—10月中旬,集中为7月下旬—9月中旬,其花芽分化进程与串枝红、银香白[9]的表现相似,但其进入分化期较辣椒杏[19]晚30 d左右。8号、28号、595号无性系的花瓣分化期均在8月中旬结束,而李光杏[8]、早熟黑叶杏[9]的花瓣分化期可持续至9月初。622号、626号无性系的花原基分化期、萼片分化期、花瓣分化期均为8 d,其持续时间均短于其他3个无性系的,而与龙王帽[20]的研究结果(10 d)相近,可能因为622号、626号无性系均为山西种源,与龙王帽的种源地(陕西)相近。Bartolini等[21]分别于夏、秋季对普通杏进行水分胁迫试验,结果表明,水分胁迫可导致其花芽分化进程滞后甚至停止。此外,Zhang等[22]的研究结果表明,含氮化合物有利于核桃花芽的分化;袁小军等[23]认为,施加氮磷钾可以促进油茶花芽的分化;Xiao等[24]发现,麻风树雌花原基在最初发育时需要细胞分裂素(CTK)的诱导,而发育后期只需较低浓度的CTK。同为西伯利亚杏的8号、28号、595号、622号、626号无性系其花芽分化各时期的起止时间、持续时间相差较大的原因可能是其自身抗性不同、对养分需求不同或自身激素含量不同,因此应根据不同无性系花芽发育状态制订适时、合理的栽培管理方案,如采取适时整形修剪、合理施肥灌溉等措施以促进西伯利亚杏花芽的形态分化。
研究中发现,辽宁喀左地区的西伯利亚杏花芽于3月中旬开始萌动发育,4月初进入花期,持续约10 d,其开花物候期与新疆伊犁的野生杏群体[12]的相近,比陕西周至的栽培杏[9]晚20 d左右,较内蒙古和林的西伯利亚杏[25]早15 d左右,这可能与各地经纬度、海拔等地理条件以及气温、降水等气候因子[13]或者自身遗传特性[26]不同有关。此外,供试的西伯利亚杏各无性系的开花物候期较为一致,其花期集中于4月上旬,有利于在花期统一进行避霜冻管理。
西伯利亚杏各无性系的花瓣、花丝、花药、花萼颜色一致,但8号、28号、595号、626号的花瓣均为典型的五瓣花,而622号的花瓣发生变异,为六瓣花。刘华堂等[27]根据花冠直径(D)将39个杏品种花的类型划分为大型花(D>3.0 cm)、中型花(2.5 cm≤D≤3.0 cm)、小型花(D<2.5 cm)这3类,据此,8号、28号、622号、626号的花瓣均为中型花,而595号的花瓣则为小型花。
杏属植物易分化形成花芽,但不同品种间其完全花比例往往不同[15],并且同一品种不同类型果枝间也有较大差异[28]。研究中发现,西伯利亚杏不同无性系间完全花比例的差异显著,这可能与各品种、无性系间遗传特性不同有关。西伯利亚杏不同类型果枝间完全花比例的差异较大,具体表现为:花束状果枝(46.66%)>短果枝(32.24%)>中果枝(27.25%)>长果枝(24.84%)。这一观测结果与刘立强等[29]对新疆杏的研究结果一致。但是,张鑫等[13]观察发现,山杏不同类型果枝的完全花比例的大小顺序为长果枝>花束状果枝>短果枝>中果枝,这可能与杏树自身树体结构、营养分配不均等因素的影响有关。因此,可根据花期物候进程及气候条件,针对杏树可采取适度的枝条修剪、合理施肥等措施以促花保花,从而提高其完全花比例,提高其坐果率和果实产量。
生长素在植物雌蕊形态形成与生长发育中具有重要作用。Shi等[30]在对果梅可育花与败育花差异基因的筛选中发现,59个基因参与了生长素信号通路,并且可育雌蕊中生长素的含量(50.03 ng/g)显著高于败育雌蕊的(28.29 ng/g),表明缺少生长素可能导致雌蕊败育。本研究在西伯利亚杏盛花期选取完全花比例最高的28号(99.42%)与最低的595号(3.50%),对其生长素类激素的含量进行测定,结果表明,28号的IAA、IAA-Asp、总含量均极显著高于595号的,且28号的IAA含量是595号的2.78倍,表明IAA可能是595号完全花比例低的影响因素,这一测定结果与庆军等[5]、Shi等[30]的研究结果均一致。
本研究仅从形态学方面观测西伯利亚杏花芽分化及开花物候进程,并未深入研究其内在机理,今后可进一步测定其花芽内碳水化合物等养分或其他种类激素的含量,以深刻剖析其原因,从而合理调控其花期。
3.2 结 论
西伯利亚杏花芽分化时期可分为未分化期、花原基分化期、萼片分化期、花瓣分化期、雄蕊分化期、雌蕊分化期这6个时期,其花芽分化集中于7月下旬—9月中旬,但不同无性系进入不同分化时期的起止时间和持续时间不同,且其各个时期具有明显的重叠现象。西伯利亚杏于3月中旬进入花芽膨大期,4月上旬进入始花期,其花期持续约10 d,且各个无性系间的物候基本一致,可统一进行花期调控等栽培管理措施。西伯利亚杏不同无性系间完全花比例的差异较大,同一无性系不同类型果枝完全花比例的大小顺序表现为:花束状果枝>短果枝>中果枝>长果枝。因此,在西伯利亚杏无性系的栽培管理中,应采取有针对性的修枝、促花、保花等措施。
