花生花器官形态建成及异型雄蕊发育研究
2019-10-22王海霞陶莹芳丁雨龙
王海霞,陶莹芳,丁雨龙
(1. 南京林业大学南方现代林业协同创新中心/生物与环境学院/竹类研究所,江苏 南京 210037;2. 青岛农业大学化学与药学院,山东 青岛 266109)
花生(Arachishypogaea)属于豆科(Leguminosae)、蝶形花亚科(Papilionaceae)、花生属(Arachis),目前花生属内被描述的种已经增加到80个[1-2],根据形态性状、地理分布和杂交亲和性,将花生属分成9大区组,其中花生区组最大,包含花生栽培种(ArachishypogaeaL.)。Krapovickas和Gregory[1]根据主茎是否开花,将花生分为两个亚种Subsp.hypogaea和Subsp.fastigiata。亚种hypogaea被分为var.hypogaea和var.hirsute两个变种。亚种fastigiata被分为var. fastigiata和var. vulgaris两个变种。孙大容等根据花生荚果、籽仁、生育习性、开花习性等农艺学综合性状,将中国栽培花生的品种分为普通型、龙生型、珍珠豆型和多粒型四类[3],这与Krapovickas和Gregory的分类原则基本一致。
关于花生花芽分化的研究较早,崔澧等人将花生花器的分化过程分为9个时期[4]。王福青依据花生花器官形成及雌雄配子体的发育顺序,将花芽分化过程划为10个时期,发现雌雄配子体形成时间不同步[5-6]。白秀峰对“鄂花二号”、“白沙1016”等花生品种的花器进行了解剖观察,发现大孢子母细胞的发育迟于小孢子母细胞,但成熟先于小孢子母细胞[7];花生的花器较小,有雄蕊10枚,大部分品种的花中有4枚长椭圆形花药的雄蕊又称为长药雄蕊,药室较大,花丝较长;4枚圆球形花药的雄蕊又称为圆药雄蕊,药室较小,花丝较短;0~2枚退化雄蕊,即存在着雄蕊异型现象。在不同环境条件下,两枚退化的雄蕊发育成花药的比例不同[7]。赵世绪等人根据花蕾外部形态特征将小孢子发育分为4期,一朵花中长药中小孢子早期的发育较圆药中快,同一花药内小孢子的发育时期相同[8]。
虽然对于花生花器官发育过程有了一定的研究,但是这些研究大多基于光学显微镜的观察,缺乏形态发育上的系统研究。本研究将以五种类型花生品种为研究对象,从形态学、细胞学方面利用扫描电镜、石蜡切片技术对花生花器官模式形态建成以及异型雄蕊的发育过程进行研究,以期为解释花生花发育相关基因的时空表达以及花生育种提供形态解剖学依据。
1 材料与方法
1.1 材 料
本研究选用花生栽培的典型品种:普通型弗罗蔓生(Arachishypogaeavar.hypogaea'Florunner')、龙生型永城小麻壳(A.hypogaeavar.hirsuta'Yong-cheng Xiaomake')、多粒型四粒红(A.hypogaeavar.fastigiata'Silihong')、珍珠豆型白沙1016(A.hypogaeavar.vulgaris'Baisha1016')[9]以及异源四倍体野生种A.monticola的为材料(种子由青岛农业大学张晓军老师提供),于2018年4月末覆膜种植于青岛农业大学试验田。
1.2 方 法
1.2.1扫描电镜
对五品种的花生在主茎3、4、5、6、7、8叶时,取样6次,每次选有代表性的20株,分别取第一对侧枝第一节侧枝上的第一个花序的第一个花芽[5],用游标卡尺测量花蕾长度后,经70% FAA固定、0.1 mol/L PBS冲洗、乙醇系列脱水、乙酸异戊酯置换,在HCP-2型临界点干燥仪内进行临界点干燥,IB-5型离子溅射仪内进行镀膜,JEOL7500F环境扫描电子显微镜下观察并拍照。
1.2.2 石蜡切片法
按照1.2.1方法取不同发育时期的花蕾,用70% FAA固定24 h后,经酒精系列脱水,二甲苯透明,石蜡包埋,常规石蜡切片法制片,切片厚度8~10μm,利用铁矾—苏木精染色,中性树胶封片,Olympus-BH2型显微镜观察并拍照。
2 结果分析
经研究发现:四个花生栽培种和一个异源四倍体野生种A.monticola花器官的发育进程基本一致,仅退化雄蕊存在数量上的差异,永城小麻壳为1枚退化雄蕊,而弗罗蔓生、四粒红、白沙1016以及A.monticola的花中均有两枚退化雄蕊,退化雄蕊与心皮腹缝线对生。因此花生花器官形态建成及异型雄蕊发育研究结果将以永城小麻壳和弗罗蔓生为例进行阐述。
2.1 花生花器官的发生
根据扫描电镜观察,将花生花的个体发育分为8个发育阶段:阶段1:初级花序分生组织侧向分化为次级花序分生组织以及叶原基,次级花序分生组织横向分化出分生组织和苞片(图1, a);阶段2:花芽原基呈钝圆形圆锥体,苞片逐渐发育,被毛(图1, b);阶段3:花原基顶端变平,分化出萼片原基和共同原基(图1, c);阶段4:萼片伸长,向中间聚拢并包裹共同原基。共同原基分化出心皮原基和对萼雄蕊原基(图1, d);阶段5:第二轮的花冠原基和第三轮对瓣雄蕊原基同时发育(图1,e);阶段6:旗瓣原基,翼瓣原基,龙骨瓣原基出现。雄蕊开始分化花药,心皮逐渐闭合并弯曲,内部形成子房室。此时有一枚或两枚与心皮腹缝线对生的雄蕊未见花药发育,仅存花丝(图1,f),将来发育为退化雄蕊。对萼的长药雄蕊的花药明显起棱,对瓣的圆药雄蕊发育较慢,两者之间的区别逐渐显现,心皮腹缝线对生的对萼雄蕊比心皮背部对萼雄蕊的花药略小(图1,g);阶段7:5枚花瓣覆瓦状排列,旗瓣发育较快,翼瓣的先端逐渐加宽、伸长,两片龙骨瓣愈合向上方(图1,h)弯曲发育,逐渐将雌雄蕊包含在内;雄蕊花丝迅速伸长,此时可见永城小麻壳一枚退化雄蕊(图1,i),弗罗蔓生一长一短雄蕊退化(图1,j)退化雄蕊仅存花丝,顶端表皮细胞皱缩,角质层具条纹,无花药(图1,k)。可育雄蕊的花药表皮细胞膨大、光滑。雌蕊伸长,柱头开始分化(图1,l);阶段8:成熟可育雄蕊的花药在大小和形状上存在显著差异,对瓣圆药雄蕊的花药体积约为对萼长药雄蕊花药的1/8(图1,i、j)。细长的花柱弯曲呈勾状,柱头上形成毛状乳突,高于花药(图1, m),子房上位;雄蕊基部联合成单体雄蕊管,基部呈90°弯曲(图1,n);龙骨瓣包住柱头和花药,未见花蜜腺发育,花发育成熟,永城小麻壳的对萼雄蕊、对瓣雄蕊中的花粉粒形状、大小和纹路相同(图1,o、p)。

I.花序分生组织;Br.苞片;C.心皮;Vx.旗瓣;A.翼瓣; F.花原基;K.龙骨瓣;P.花瓣原基;Pg.花粉粒;C.心皮原基;S.萼片;Sg.柱头;Sta.退化雄蕊; Stp.对瓣圆药雄蕊; Sts.对萼长药雄蕊;Stu.单体雄蕊管I:Inflorescence meristem; Br:Bract; C: carpel primordial; Vx: vexillum; A: alae; F: floral primordium; K:keel; P:petal primordium; Pg:Pollen grain;S: sepals; Sg: stigma; Sta: staminodes; Stp:antepetal stamen; Sts:antesepal stamen; Stu:staminal tube 图1 花生花器官发生的扫描电镜图Fig.1 Scanning electron micrographs of flower organogenesis of Arachis hypogaea L.
注:a-n.花生花发育的不同时期(f、j、m为弗罗蔓生,其余为永城小麻壳)。a. 阶段1:初级花序分生组织侧向分化出次级花序分生组织;b. 阶段2:次级花序分生组织横向分化为花分生组织(F)和苞片(Br),叶(L),萼片(S);c,d. 阶段3:花原基的顶端变平,形成共同原基;e. 阶段4:共同原基分化出第二轮的对萼雄蕊原基(Sts)和第三轮的心皮原基(C);萼片伸长,向中间聚拢并覆盖其他花原基;g. 阶段5:对萼的长药雄蕊和对瓣的圆药雄蕊相间排列,对萼雄蕊(Sts)发育较快,花瓣原基(P)和对瓣雄蕊原基(Stp)同时发育,心皮开始分化、卷曲。h. 阶段6:花冠原基分化出五个花瓣原基(1枚旗瓣,2枚翼瓣,2枚龙骨瓣);l, m. 阶段7:5枚花瓣呈覆瓦状排列,旗瓣发育快,较其余四枚花瓣大,两片龙骨瓣愈合向上方弯曲发育,逐渐将雌雄蕊包含在内,心皮完全闭合并伸长,柱头弯曲、开始分化柱头,高于花药;n. 阶段8:花丝基部联合成单体雄蕊管(Stu),基部呈90度弯曲,龙骨瓣包住柱头和花药。o.永城小麻壳对萼雄蕊产生的三沟型花粉粒。p.永城小麻壳对瓣雄蕊产生的三沟型花粉粒。f.与心皮腹缝线对生的退化雄蕊;i.永城小麻壳有一枚退化雄蕊;j.弗罗蔓生有一长一短两枚雄蕊退化; k.退化雄蕊顶端表皮角质层皱缩,仅存花丝,未见花药。
Note: a-n: Different stages of peanut flower development (f, i, m 'Florunner'; the other 'Yong-cheng Xiaomake') a. The primary inflorescence meristem (I1) laterally differentiates a secondary inflorescence meristem (I2) during stage 1; b. at stage 2, the I2 forms a floral meristem (F), bract (Br), sepal(S) and leaf (L); c,d. Stage 3: the apex of flower primordium became flat and the common primordia formed; e. Stage 4: The common primordia differentiate the second and third whorl organs. Carpel(C), antesepal stamen (Sts), antepetal stamen(Stp), petal (P); g. Stage 5: Antesepal stamen (Sts) and antepetal stamen(Stp) are arranged alternately, antesepal stamen (Sts) develop faster, petal(P) and antepetal stamen (Stp) develop multaneously, carpels begin to differentiate, the carpel (C) begins to differentiate and curl , the adaxial carpel suture is visible (J); h. Stage 6: The floral organ primordia are completely differentiated, one vexillum (Vx), two alae petals (A), and two non-fused keel petals (K). Filament elongated; l, m. Stage7: Five petals were imbricate, vexillum development faster than the other four petals, two keel petals fused and curved, gradually included pistil and stamen, carpels completely closed and elongated, stigma began to differentiate; n. stage 8: The sepals have been removed. Stamens have completely differentiated the anthers; the petals develop. The formation of the staminal tube (Stu) begins; f. Staminodes opposite the sutures of carpels; i. One staminode in 'Yong-cheng Xiaomake'; j. Two staminode in 'Florunner'. k. Dehydrated epidermis of staminodes with only filaments, no sporogenous tissue at apical;o. Tricolplate pollen in antesepal stamen; p. Tricolplate pollen in antepetal stamen.
2.2 雄蕊发育的细胞学观察
花芽原基基部的外围细胞进行平周分裂和垂周分裂,形成三种异型雄蕊。永城小麻壳10枚雄蕊中包含5枚长药对萼雄蕊,4枚圆药对瓣雄蕊和1枚退化雄蕊,长、圆花药相间而生。对萼雄蕊中与旗瓣对生的2个长花药被药隔分隔成两药室,体积较其余3个四药室的长花药小;4枚对瓣雄蕊中有2个三药室圆花药、2个两药室圆花药,其中2枚三药室的圆药雄蕊与旗瓣对生(图2,a、b)。弗罗蔓生的花中有4枚长药对萼雄蕊,4枚圆药对瓣雄蕊及2枚退化雄蕊,2枚退化雄蕊位于与旗瓣对生的1枚两药室长药雄蕊和1枚三药室圆药雄蕊之间, 通常呈现为一长一短,且彼此着生位置相近,未见花药相关组织药室内壁、中层及绒毡层的出现(图2,d、e)。
在同一花粉囊内可以看到小孢子处于相同的分裂时期和发育阶段(图2,f);同一朵花中,长药雄蕊(对萼雄蕊)的花药较大,而圆药雄蕊(对瓣雄蕊)的花药小,在孢原细胞分化时,就有药室的差别,各花药的发育进程一致(图2,g)。随着花粉粒的发育成熟,药室横隔逐渐消解(图2,h)。十枚雄蕊花丝基部联合成一个单体雄蕊管,长药对萼可育雄蕊中的维管束比圆药对瓣可育雄蕊中的大(统计结果见表1)。永城小麻壳一枚退化雄蕊的维管组织面积为1895.655μm2,比其他9枚可育雄蕊的维管束小(图2,i)。弗罗蔓生中与心皮腹缝线正对的退化雄蕊花丝中未见维管束,而另一枚退化雄蕊的维管束仅比相邻圆药3室雄蕊大,但比其他可育对萼长药雄蕊的维管束小(图2,j)。

Pg:花粉粒;Sta:退化雄蕊;Sts:对萼雄蕊;Stp:对瓣雄蕊;Vb:维管束;C:心皮Pg:Pollen grain;Sta: staminodes; Sts:antesepal stamen; Stp:antepetal stamen; Vb: Vascular bundle C: carpel.图2 花生异型雄蕊的发育过程Fig.2 Development of heteromorphous stamen in peanut (Arachis hypogaea L.)
注:a:永城小麻壳5枚对萼雄蕊的长花药;b:永城小麻壳对瓣雄蕊的圆花药;c:龙生型偶见10枚可育雄蕊,其中6枚长药雄蕊,4枚圆药雄蕊;d:弗罗蔓生4枚对萼雄蕊的长花药药;e:4枚对瓣雄蕊圆花药;f:同一花粉囊内小孢子处于相同的发育阶段;g: 长药雄蕊与圆药雄蕊发育同步;h: 药室横隔逐渐消解;i: 永城小麻壳雄蕊的维管束;j: 弗罗蔓生雄蕊的维管束。
Note: a: Long oval anthers of antesepal stamens in 'Yong-cheng Xiaomake'; b: Short oval anthers of antepetal stamens in 'Yong-cheng Xiaomake'; d: Long oval anthers of antesepal stamens in 'Florunner'; e: Short oval anthers of antepetal stamens in 'Florunner'; f: Microspores in the same pollen sac are at the same developmental stage; g: Two fertile stamens develops synchronously; i: Vascular bundles in 'Yong-cheng Xiaomake'; j: Vascular bundles in 'Florunner'.

表1 永城小麻壳和弗罗蔓生中花丝维管束面积
注:以近轴端的退化雄蕊为1,顺时针标号依次为2,3,4......10。维管束的测量值为3朵处于小孢子母细胞时期的花同一切面测量,每个维管束测量3次,取平均值。
Note: The staminodes at the paraxial end are 1, and the clockwise numbers are 2, 3, 4……10. The measured value of the vascular bundle was measured on the same cut surface of three flowers in the microspore mother cell stage, and each vascular bundle was measured 3 times and averaged.
3 讨 论
花生花器官的发育与蒺藜苜蓿(Medicagotruncatula)、野生豌豆(Pisumsativum)相似[10-11]:花器官起始顺序是萼片—雄蕊、心皮—花瓣;花的各轮器官起始时间相互重叠,共同原基的出现打乱了由外向内的器官起始顺序;同一轮中出现不同属性的花器官,如花瓣与对瓣雄蕊;同一属性的器官出现在不同的轮中,如:对萼雄蕊和对瓣雄蕊;花发育后期花瓣特化,形成旗瓣、翼瓣、龙骨瓣,最终使得花呈两侧对称;常见花器官融合。花瓣融合(Sympetaly)被认为是进化的,它可以提高授粉效率,进而能加快植物多样化的速度。
永城小麻壳雄蕊的5枚对萼长花药有3个被药隔分成四药室,2个被分成两药室;4枚对瓣圆药雄蕊中有2个圆花药被分成三药室,2个被分成两药室,1枚退化雄蕊。弗罗蔓生的花中4枚长花药被药隔分成3个四药室、1个两药室;4枚圆花药分成2个三药室、2个两药室,2枚退化雄蕊(图3)。长药雄蕊、圆药雄蕊以轴两侧对称分布,呈环状相间排列,2个花生品种的雄蕊药室分布相同,性状较为稳定,未出现药室数量变化的情况,与白秀峰等人认为3个长形花药是典型的四室,4个圆形花药与1个旗瓣对生的长药药室数有变化的研究结果不一致[7],这可能与开花期的环境温度有关。开花期高温胁迫对豆科植物花药的发育有较大的影响:豇豆(VignaunguiculataL. Walp)、菜豆(Phaseolusvulgaris)和鹰嘴豆(CicerarietinumL.)热敏基因型ICC 4567、ICC 10685的花中出现花药药室数量增加,药室表皮壁增厚以及部分不育花粉粒的现象[16-18]。
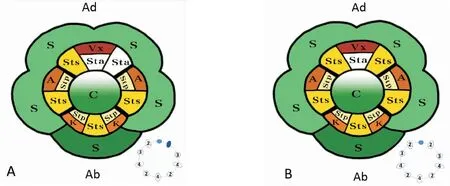
图3 花生花器官模式及雄蕊药室分布简图:永城小麻壳(左)、弗罗蔓生(右)Fig.3 Flower organ pattern and stamen chamber distribution diagram: 'Yong-cheng Xiaomake' (L) and 'Florunner' (R)
注:Ad, Adaxial(近轴端);Ab, Abaxial(远轴端);S, Sepals(5枚萼片,其中4枚连合,1枚分离);VX, Vexillum(旗瓣);A, Alae(翼瓣);K, Keel(龙骨瓣);Sts, antesepal stamen(对萼雄蕊);Stp, antepetal stamen(对瓣雄蕊);Sta,staminodes (退化雄蕊);C, carpel(心皮)。简图中2,3,4 表示药室数目,深蓝色为退化雄蕊。
Note: In the diagram, 2, 3, 4 indicate the number of pollen sac, and dark blue is the staminodes.
Paulino等人研究了羽扇豆属(Lupinus)品种'The Governor'和金雀花(Cytisusscoparius)两种开花授粉的花器个体发育以及花粉粒的功能,认为在这两个蝶形花亚科植物中异型雄蕊的发育与花粉结构和功能的差异相对应,有很强的功能划分:在羽扇豆中雄蕊异型性与花粉释放机制有关,对萼雄蕊产生可育花粉且花粉数量多,而对瓣雄蕊产生不育花粉且花粉数量少;在金雀花中一部分雄蕊生产花粉给传粉者提供食物,另一部分雄蕊产生的花粉则用来授粉,避免了花粉被传粉者过度采集,是一种为保存足够多花粉以供授粉的机制[19]。异型雄蕊的分化与各自产生的花粉活力的差异,表明雄蕊之间存在着分工,可能通过传粉媒介介导的选择进化,以减少由花粉的不同功能引起的冲突[19]。相比开花授粉花,闭花授粉花更原始[20]。所有闭花授粉花的共同特征是雄蕊数量或体积的减少和花冠闭合[21-22]。对花荵科的大花粘胶花(CollomiagrandifloraDouglas ex Lindl.,Polemoniaceae)同一花序轴上闭花受粉和开花受粉两种花的观察发现:闭花授粉花的花冠减少,两种花的花药大小和形状存在着显著差异[23]。闭花授粉的花中花药体积小,仅有两个药室,花粉数不足50粒;而开花受精的花中花药体积大,有四个药室,能产生500多个花粉粒。闭花授粉的花药中较少的初始孢原细胞和较短的孢子细胞增殖持续时间与较小的花药相关,其成熟时花粉粒数仅为开花授粉花的1/10[24]。花生为严格的闭花授粉[25],龙骨瓣的出现保存了大量花粉以供繁殖。
对云实属金凤花(Caesalpiniapulcherrima)的研究证实,花内雄蕊花药开裂时间的分化延迟了花内花粉的存在时间,通过多批、分次的传粉者散布,有效的促进了花粉的输出和成功授粉率[25]。本研究发现,花生花内两种异型可育雄蕊同步发育(图2,a、b;d、e),这与赵世绪等人“花药分化早期,长药中小孢子发育较圆药中快”的结论不一致[8]。但两者散粉时间不一致:长药雄蕊(对萼雄蕊)的花丝较长,先散粉;而圆药雄蕊(对瓣雄蕊)花丝较短,散粉晚。雄蕊在花丝与花药上的分化使得花粉在不同空间位置、不同时间散粉,可能是花生控制花粉呈现的装配与分发机制之一。席湘媛利用透射电镜观察了花生品种徐州68-4成熟花粉的超微结构,但未对两种异型可育雄蕊产生的花粉进行区分探讨[27]。本研究发现,同一品种花生花内的两种异型可育雄蕊均可产生三沟型花粉,两种花粉形态、大小和纹饰均相同。
永城小麻壳和弗罗蔓生这两个品种的花中长药雄蕊花丝维管束面积均比圆药雄蕊的大,一枚退化雄蕊的维管组织面积均比其他可育雄蕊小,弗罗蔓生中与心皮腹缝线正对的退化雄蕊未见维管束。雄蕊花丝中维管束的大小与花药的大小呈正相关,且退化雄蕊维管的发育程度与退化雄蕊的大小有关[28]。Rodríguez等人对具有不同大小退化雄蕊的玄参属(Scrophularia)玄参组(Sect.Scrophularia)的S.scorodonia,S.lyrata和砾玄参组(Sect.Tomiophyllum)的S.canina三种植物花的维管系统进行研究,发现较大的退化雄蕊比较小的退化雄蕊具有更多的维管系统。
永城小麻壳和弗罗蔓生两个品种的花不仅退化雄蕊数量有差异,退化雄蕊的着生位置也不同:永城小麻壳1枚退化雄蕊位于2枚两药室对萼雄蕊之间;弗罗蔓生2枚退化雄蕊相邻,位于1枚两药室对萼雄蕊与1枚三药室对瓣雄蕊之间,为一长一短。根据花药药室分布和花型两侧对称的规律可知,弗罗蔓生邻近三药室对瓣圆药雄蕊的1枚二药室对萼雄蕊花药未发育,仅存花丝,成为退化雄蕊。研究中偶见永城小麻壳花内有6枚长药雄蕊、4枚圆药雄蕊,可见退化雄蕊有进一步发育的可能,这与白秀峰的结果一致[7]。
Mayer和Chartesworth认为雄蕊的保留是由于雄蕊和雌蕊之间的遗传相关性所致,这延迟了花中的雄蕊或雌蕊功能的抑制[31]。Endress提出,雄蕊减少是因为雄蕊的正向存在对授粉不利,雄蕊可能在此过程中阻碍花的功能,在第五(奇数)雄蕊的减少上表现出广泛的变异与对称程度相关[32]。背腹不对称在花瓣和雄蕊中最为明显,每个花瓣和雄蕊可分为三种类型:背侧、侧面和腹侧[33]。两侧对称的花中正面最上部奇数雄蕊通常会减少,其与花冠筒融合以占据对称线交叉的位置[32],如金鱼草中背部雄蕊原基在发育的早期阶段滞育,形成一个退化雄蕊,在金鱼草中分离出控制花不对称性的第一个基因cycloidea[34],在花分生组织的背侧区域早期表达,影响着花原基的形成及其分化程度;表达持续到花原基后期,将会影响花瓣和雄蕊的不对称性、大小和细胞类型。
栽培种花生的染色体为异源四倍体(2n=4X=40),基因组大且遗传基础狭窄,又因花器官的性状稳定,因此花的遗传多态性极低。四个花生品种和一个野生种花的发育进程与形态基本一致,仅退化雄蕊存在着数量上的差异。龙生型花生(茸毛变种)在植物学性状和品质性状上变异较大,具有丰富的遗传多样性,比较原始,可以以此变种来研究花生的进化,了解花生进化历程[35]。还可根据不同目的,选择此类型的不同品种在育种上加以利用[36-37]。
花生花内异型雄蕊现象在被子植物系统进化中有何意义?两种可育雄蕊在功能上有何差别?退化雄蕊虽然丢失了雄性繁殖功能,但是否在进化过程中重新获得了有助于植物繁殖的新功能?仅从雄蕊发育的角度来解释是远远不够的,还需要多学科手段来进行深入、系统的研究。
