卡拉胶/α-淀粉酶复合冻融稳定性对馒头的影响
2023-01-10黎钧铸黄文晶沈汪洋黄庆荣金伟平
黎钧铸,黄文晶,沈汪洋,王 展,黄庆荣,金伟平
(武汉轻工大学食品科学与工程学院,湖北 武汉 430023)
冷冻面团技术是把面团低温冷冻处理为半成品或成品,方便烘焙类食品加工的商业化生产的一种工艺[1]。冷冻面团加工有效解决了食品淀粉老化、口感变劣、货架期短等问题,同时降低了面制品的生产成本,提高面团质量控制能力以及产品的新鲜度和稳定性[2]。但也出现一些问题,例如冷冻过程会使面筋结构损坏、酵母活性减弱、后期醒发时间延长、酸味明显、膨胀力变差、影响外观等[3]。
α-淀粉酶是一种常用于面制品加工的酶制剂。一方面,它能促使淀粉分解产生更多的糖来补偿面团加工过程中糖的损失,为发酵提供能量[4];另一方面,α-淀粉酶分解淀粉部分产物为麦芽糖、糊精等,抑制淀粉重结晶,从而延缓面制品老化、增加产品比容。陈书明等通过测试不同浓度α-淀粉酶、木聚糖酶对冷冻酵母的影响,发现冷冻1 d 时,添加0.4 g/kg α-淀粉酶的面团中酵母存活率达到98%以上,抗冻性明显提高[5]。
食品是一种多组分共存的复杂体系,共存物对α-淀粉酶的结构变化及其加工敏感性均有一定的影响。Giannone 等人将脂肪酶制剂与4 种不同商品淀粉酶进行复配使用,结果显示α-淀粉酶和脂肪酶在防止老化方面呈现显著协同作用,有效改善样品的柔软性和耐咀嚼度[6]。林娟娟等人发现在面团中添加α-淀粉酶、脂肪酶和半纤维素酶,使面包组织结构更加细腻,口感较柔软[7]。王建芳等人将淀粉酶和谷氨酰胺转氨酶混合加入燕麦和小麦混合粉面团中,使面团的色泽以及流变特性更接近小麦面团,但当体系存在黄原胶时,这两种酶的改善效果未得到提升[8]。
卡拉胶作为一种天然亲水胶体可显著改善冷冻食品的外观,降低冰晶的形成速度,改善面筋蛋白流变学特性[9],从而提高冷冻食品体系的稳定性。亲水胶体的添加可减少自由水,增加结合水,通过组分间相互作用形成新结构,从而提升产品体系在低温环境下的稳定性。在食品加工过程中,多糖胶体的存在及分子结构对蛋白质/酶结构的影响和相互作用是一个亟待研究的问题。单一α-淀粉酶对面团的作用效果已有很多研究,而λ-卡拉胶/α-淀粉酶复合物在冷冻面团及其面制品中的研究相对较少。作者首先研究了λ-卡拉胶对冻融过程中α-淀粉酶性质的影响,进而探究了复合物对冷冻面团发酵性能及馒头的比容、质构、孔隙分布等参数的影响。
1 材料与方法
1.1 材料与试剂
α-淀粉酶(EC 3.2.1.1,来源于地衣芽孢杆菌)、λ-卡拉胶:均购自阿拉丁(中国上海);其他化学品包括碘、碘化钾、氯化钠、可溶性淀粉、考马斯亮蓝:均购自国药化学试剂有限公司(中国上海);多用途麦芯小麦粉:购自益海嘉里粮油有限公司(中国上海);高活性干酵母:购自安琪酵母有限公司(中国宜昌)。研究中使用的化学品均为试剂级,在使用前没有经过任何纯化。
1.2 仪器与设备
UV-5500PC 型紫外可见光分光光度计:上海元析仪器有限公司产品;F-4600 型荧光分光光度计:日本日立公司产品;ME104E 型电子分析天平:瑞士梅特勒-托利多公司产品;NG 吹泡稠度仪:法国肖邦技术公司产品;发酵流变仪:法国肖邦技术公司产品;食品体积测定仪:瑞典波通仪器公司产品;食品图像分析仪:瑞典波通仪器公司产品;TA 物性测试仪:英国Stable Micro System 公司产品;FiveEasy Plus 型pH 测定仪:瑞士梅特勒-托利多公司产品。
1.3 实验方法
1.3.1 λ-卡拉胶/α-淀粉酶复合物的制备取一定量α-淀粉酶和λ-卡拉胶样品分别配制质量浓度为0.2 mg/mL 的母液,α-淀粉酶室温搅拌30 min 至完全溶解,λ-卡拉胶85 ℃搅拌2 h 完全溶解。将配制好的α-淀粉酶和λ-卡拉胶母液以体积比10∶1 的比例混合,α-淀粉酶终质量浓度为0.1 mg/mL,调节溶液pH 为5.0,置于4 ℃冰箱充分溶解,过夜后使用。
1.3.2 冻融循环处理将配置好的α-淀粉酶以及λ-卡拉胶/α-淀粉酶复合物置于-20 ℃冰箱冷冻,12 h 后取出置于室温解冻2 h,记作第1 次冻融(F1),再将解冻好的样品放入-20 ℃的冰箱中冷冻12 h,取出后在室温中解冻2 h,记为第2 次冻融(F2),同样的方法进行第3 次冻融(F3)。
1.3.3 α-淀粉酶的酶活测定依据GB1886.174-2016 酶制剂活性测定法,测定α-淀粉酶的酶活,并略做修改。吸取1 mL 可溶性淀粉溶液于离心管中,加入0.25 mL 的0.2 mol/L 磷酸盐缓冲液(pH 6.0),摇匀后,置于60 ℃恒温水浴中预热8 min 以上。加入0.05 mL 稀释好的待测酶液,摇匀反应5 min,吸取0.05 mL 反应液,加到预先盛有0.025 mL 盐酸溶液和0.25 mL 稀碘液的离心管中,摇匀,并以0.025 mL 盐酸溶液和0.25 mL 稀碘液为空白,于660 nm波长下用酶标仪测定其吸光度,计算酶液的酶活性。用Bradford 法在595 nm 处测定其吸光度,计算蛋白质比例,得到酶活(U/mg)。
1.3.4 内源荧光分析将经冻融处理后的α-淀粉酶以及λ-卡拉胶/α-淀粉酶复合物稀释50 倍,使用日立F-4600 荧光分光光度计测内源荧光,仪器设置参数如下:激发波长280 nm,扫描波长范围300~500 nm,扫描速度240 nm/min,狭缝宽度5 nm,激发电压700 V。
1.3.5 粒径与电位分析使用马尔文动态光散射仪对经冻融处理后的α-淀粉酶以及λ-卡拉胶/α-淀粉酶复合物进行粒径分布以及电位的测量。粒径稀释100 倍,避免多重光散射。
1.3.6 面粉水分质量分数的测定参考国家标准食品中水分的测定(GB 5009.3—2016)中的直接干燥法,计算出面粉水分质量分数。
1.3.7 发酵加水量的测定根据国家标准小麦粉面团流变特性测定(GB/T 14612.4—2005)中的表格查出被测面粉水分质量分数应加入的氯化钠溶液毫升数。
1.3.8 发酵性能的测定取250 g 面粉,3 g 干酵母,5 g 盐和150 mL 未冻融处理的样品溶液(0.1 mg/mL α-淀粉酶溶液;0.1 mg/mL λ-卡拉胶溶液;λ-卡拉胶/α-淀粉酶复合物(酶质量浓度0.1 mg/mL);空白溶液),采用肖邦吹泡仪和面,和面结束后用保鲜膜包好面团,立即放入冰箱冷冻。12 h 后拿出解冻,取315 g 面团置于发酵流变仪中,测试参数为:28.5 ℃,4 h。
1.3.9 冷冻面团的制备500 g 面粉,6 g 干酵母,120 mL 水(发酵实验加水量的80%)。在和面机中依次均匀倒入面粉、酵母和样品溶液,常温下和面6 min 后取出,迅速切割并揉捏成80 g/个的面团,用保鲜膜包好并做好标记,放入冰箱贮存12 h,即得到冷冻面团。
1.3.10 馒头的制备 参照GB/T 35991—2018 方法并略有改进[10]。将冷冻面团在常温下解冻,设定恒温发酵箱的温度为35 ℃,相对湿度为83%,放入面团发酵30 min,再放入电蒸锅的上层蒸15 min 左右,停火后勿打开锅盖,继续焖3 min 后取出冷却待用。
1.3.11 馒头孔隙分布的测定将馒头切成厚片状,校准仪器后,取中间一块切片放在C-Cell 图像分析仪的样品盒中,分析得到馒头切片的6 张图像和气孔的各项数据[11]。
1.3.12 馒头质构的测定取馒头样品,在馒头中心处切出边长为2 cm 的正方体,用TA 物性测试仪的TPA 模式进行测定。参数设置:探头型号为P/45R;测前速度、测中速度、测后速度均为3 mm/s;下压程度50%;触发力5.0 g,得到馒头的硬度、弹性等数据[12]。
1.3.13 馒头比容的计算参照GB/T 35991—2018,取馒头样品称其质量,用食品体积测定仪测定馒头的体积和高度,并计算出馒头的比容[10],公式如下:
式中:v为比容,mL/g;V为体积,mL;m为质量,g。
1.4 数据统计分析
每个实验设置3 组平行实验,最终数据以3 次平行实验的平均值和标准偏差来表示,结果通过SPSS18 软件做单因素方差分析,显著性水平P<0.05,图表采用Origin 2019 绘制。
2 结果与分析
2.1 冻融次数对α-淀粉酶及λ-卡拉胶/α-淀粉酶复合物结构的影响
不同冻融处理后的α-淀粉酶和λ-卡拉胶/α-淀粉酶复合物的剩余酶活见图1。随着冻融次数增加,α-淀粉酶剩余酶活呈下降趋势。冻融1 次,α-淀粉酶酶活下降到88%,此时α-淀粉酶发生冷冻变性[13]。胡海玥等人发现经冷冻处理后的大豆分离蛋白稳定性以及乳化性质都有所下降[14]。增加冻融次数,α-淀粉酶酶活性的下降趋势趋于平缓并最终稳定在79%。然而λ-卡拉胶/α-淀粉酶复合物剩余酶活在冻融一开始就大幅下降。在相同的冻融次数下,复合物的酶活也显著低于原酶,推测λ-卡拉胶与α-淀粉酶的相互作用在冻融过程中促进了α-淀粉酶结构的展开。

图1 α-淀粉酶和λ-卡拉胶/α-淀粉酶复合物经不同次数冻融处理后的剩余酶活Fig.1 Enzyme activity retention of α-amylase and λcarrageenan/α -amylase complex after different freeze-thaw cycles
内源荧光强度常用来表明蛋白质在三级结构水平上的构象变化[15],图2显示了α-淀粉酶及λ-卡拉胶/α-淀粉酶复合物经不同次数冻融后内源荧光的变化。蛋白质的内源荧光来自色氨酸、酪氨酸和苯丙氨酸,其荧光光谱对环境极为敏感,其中色氨酸的荧光强度最大,当最大吸收波长大于330 nm时,证明此时色氨酸残基暴露于外部的极性环境中[16]。在冻融1~2 次后,α-淀粉酶内源荧光强度增大,表明色氨酸残基微环境极性增加,更多发光基团暴露于溶液中。冻融到第3 次时,荧光强度显著降低,α-淀粉酶三级结构发生变化,色氨酸的微环境从疏水内核转移到蛋白质表面,极性增大,从而导致荧光强度快速下降,同时位于疏水区域的活性位点也发生改变,酶活性降低。λ-卡拉胶/α-淀粉酶复合物内源荧光强度随着冻融次数的增加而降低,复合物荧光强度的降低是因为λ-卡拉胶与α-淀粉酶通过静电相互作用形成复合物,蛋白质分子上的色氨酸残基在结合中被掩盖[17-18]。在相同冻融次数下,复合物的荧光猝灭程度要显著大于α-淀粉酶的猝灭程度,三级结构变化更为剧烈,这与酶活的变化趋势相同。
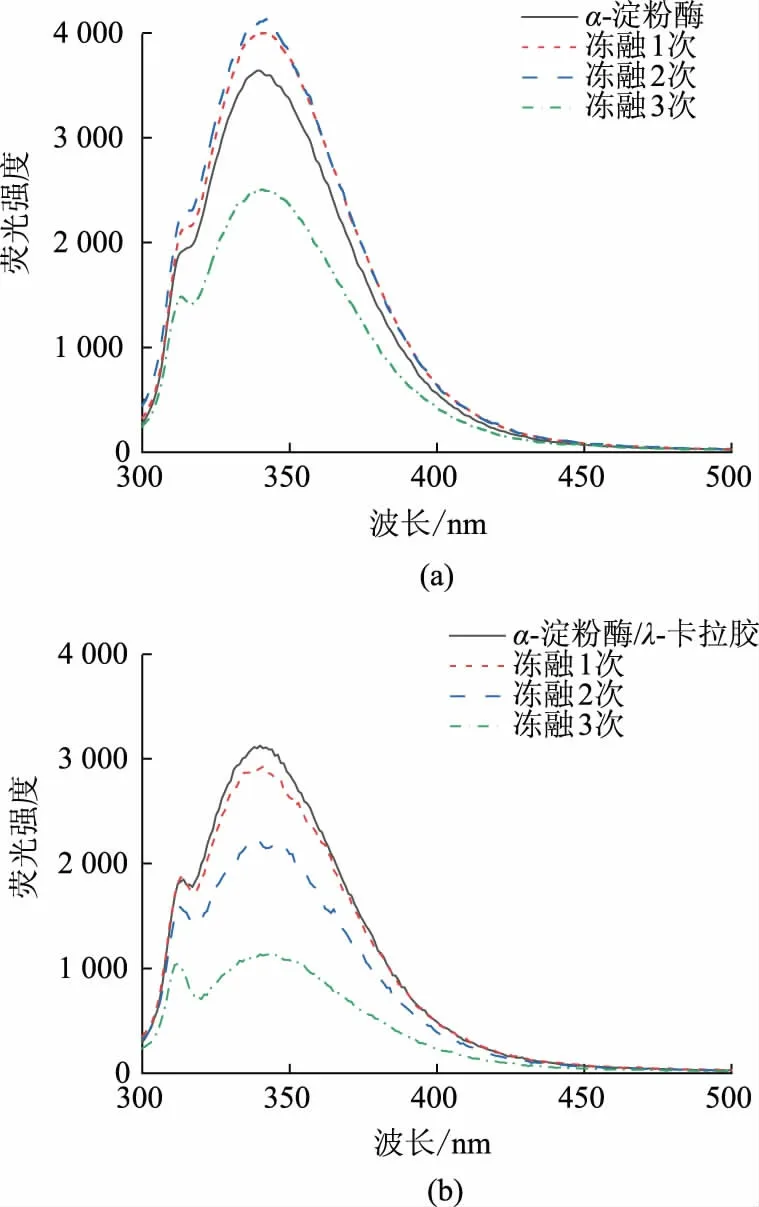
图2 α-淀粉酶和λ-卡拉胶/α-淀粉酶复合物经不同次数冻融处理后的内源荧光强度Fig.2 Intrinsic fluorescence of α -amylase and λ -carrageenan/α-amylase complex after different freeze-thaw cycles
如图3所示,粒径表明冻融前后α-淀粉酶以及λ-卡拉胶/α-淀粉酶复合物的聚集情况。单独的α-淀粉酶水合粒径约为5 nm,当α-淀粉酶与λ-卡拉胶形成静电复合物之后粒径增大到24 nm。经多次冻融后,α-淀粉酶粒径随着冻融循环次数的增加依次增大,在冻融3 次后粒径增大到190 nm,此时α-淀粉酶三级结构展开,在疏水相互作用的驱动下发生聚集。α-淀粉酶与λ-卡拉胶通过静电相互作用形成的复合物在冻融多次后粒径无明显增大,推测此时多糖的存在抑制了α-淀粉酶的聚集。
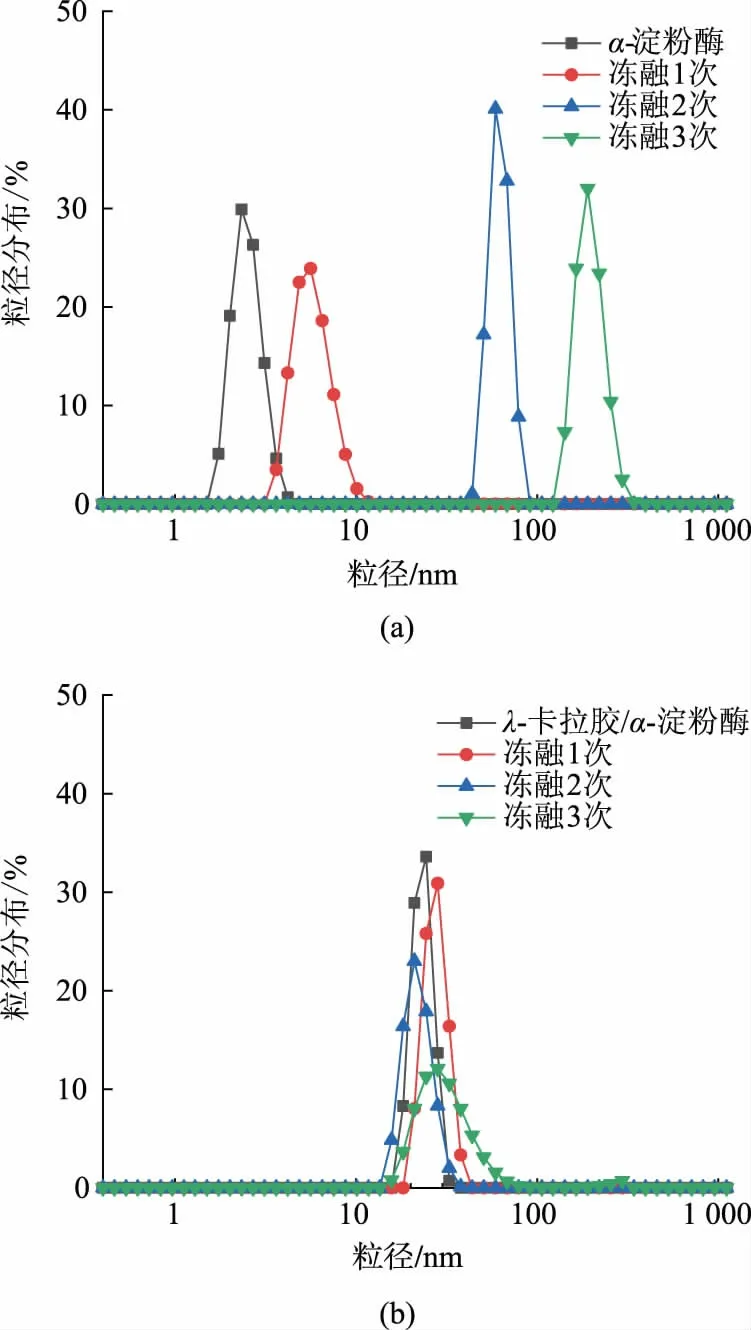
图3 α-淀粉酶和λ-卡拉胶/α-淀粉酶复合物经不同次数冻融处理后的粒径分布Fig.3 Particle size distribution of α-amylase and λcarrageenan/α -amylase complex after freeze -thaw cycles
2.2 α-淀粉酶及复合物对面团发酵性能的影响
F4 面团流变发酵仪通过记录面团发酵期间面团发酵的高度、二氧化碳气体的产生量来测量面团面筋网络的持气能力和酵母的发酵能力,见表1。可以看出,添加α-淀粉酶、λ-卡拉胶和复合物后的面团发酵膨胀高度均高于空白组,即α-淀粉酶、λ-卡拉胶和λ-卡拉胶/α-淀粉酶复合物均能促进面团发酵。α-淀粉酶可将淀粉分解为糊精、麦芽糖或葡萄糖等简单化合物,为酵母提供充足的碳源,促进酵母的生长和繁殖,极大提高酵母发酵产气,增大面制品体积[19]。λ-卡拉胶/α-淀粉酶复合物的膨胀高度要小于α-淀粉酶,酶在促进发酵上的作用显著大于卡拉胶,说明酶的活性起决定性作用[20]。由于λ-卡拉胶与α-淀粉酶形成静电复合物后酶活显著下降,因此在冻融过程中α-淀粉酶分解淀粉的能力降低,促进酵母增殖效果有限,导致了面团膨胀小。
从总产气体积的数据可知,含有α-淀粉酶和λ-卡拉胶的组别总产气量均高于空白组。一方面λ-卡拉胶中大量的羟基使它与面团中的水分子以氢键形式结合,减少水分的散失,利于酵母繁殖并提高酵母细胞的活性[21]。另一方面,λ-卡拉胶可提升水相黏度,增加产气气体包裹液膜的延展性和强度,优化面筋网络结构,提升面团持气能力[22]。复合物组与α-淀粉酶组则无显著性差异,这是因为λ-卡拉胶不参与面团气体的产生,α-淀粉酶在其中发挥主要作用,因此总产气体积无显著变化。
2.3 α-淀粉酶及复合物对馒头比容的影响
馒头比容受酵母的产气能力与面筋网络结构的持气能力两个因素影响,图4为不同添加组分对馒头比容的影响。含有α-淀粉酶、λ-卡拉胶和复合物的馒头比容均比空白对照组大(2.51 mL/g),其中添加α-淀粉酶的比容最大,达到2.73 mL/g。在冻融过程中大约有20%的酵母细胞死亡,α-淀粉酶水解淀粉产生小分子糖可供能酵母菌,增加产气量。另一方面,冷冻过程形成的冰晶会破坏面筋蛋白网络结构,制约馒头体积增大。少量的α-淀粉酶可部分破坏淀粉颗粒,使淀粉与面筋蛋白之间的结合更充分,增加面筋强度,提升持气能力,增大馒头比容,使外观更蓬松更美观。类似的现象在添加木聚糖酶的全麦粉面包中也有发现,木聚糖酶水解阿拉伯木聚糖,导致面包比容增大[22]。另外,λ-卡拉胶作为一种阴离子多糖,能通过氢键吸附大量自由水,提高面筋蛋白持水性。添加了复合物的馒头比容要低于添加α-淀粉酶组,但显著高于λ-卡拉胶组和空白组,说明复合物能有效保护面团网络结构的完整性,保持面团持气性。但由于α-淀粉酶和λ-卡拉胶形成复合物后,λ-卡拉胶抑制了α-淀粉酶的活性,同时部分λ-卡拉胶参与静电复合,结合水能力降低,削弱酶对馒头品质的影响。

图4 不同添加组分对馒头比容的影响Fig.4 Effects of different additives on the specific volume of steamed bread
2.4 α-淀粉酶及复合物对馒头质构的影响
馒头的质构特性是与感官性质联系较为紧密的指标,其中,弹性与馒头品质呈正相关,弹性数值大,品质好;而硬度与馒头和面包品质呈负相关[23]。面团质构的变化主要有3 大影响因素:一是面团在冷冻过程中,由于冰晶的作用使水分重新分布,引发面筋蛋白分子解聚,形成的面筋黏弹网络结构遭到破坏[24];二是冷冻导致面筋蛋白低温变性,使其隐藏在其内部的部分疏水基团暴露,面筋蛋白持水性减弱[25];三是损伤淀粉的含量增加,淀粉吸水率增强,使水分从面筋基质中流出,面团发黏且不易成型,弹性和可伸展性下降[26]。
图5为不同添加组分对馒头硬度和弹性的影响。与空白组相比,添加α-淀粉酶、λ-卡拉胶和λ-卡拉胶/α-淀粉酶复合物均不同程度降低了馒头的硬度,增加弹性。λ-卡拉胶对馒头硬度有显著影响,但对弹性影响不大,推测原因为λ-卡拉胶的硫酸根阴离子基团能与面筋精氨酸中带正电荷的基团通过静电相互作用形成复合物,改变面筋蛋白疏水性,从而提高面团持水能力[27]。α-淀粉酶可以显著降低馒头的弹性,提升馒头的柔软度,对比复合物组发现,馒头的硬度降低,弹性增加不明显,推测α-淀粉酶和λ-卡拉胶之间发生了相互作用,λ-卡拉胶类似于α-淀粉酶抑制剂。结果表明,酶制剂和亲水胶体都能改良馒头的品质,但在实际生产中应该结合其对面团、馒头性质的影响来调整添加水平,而复合物对馒头质构特性的影响要弱于酶制剂,强于亲水胶体。
2.5 α-淀粉酶及复合物对馒头孔隙分布的影响
C-Cell 图像分析仪可得到馒头内部纹理气孔参数和切片图像。馒头内部组织结构的细腻程度直接影响消费者的体验感。有研究表明,气孔数量和孔洞数量与比容的相关系数高达90%。通常品质好的馒头具有气孔数量多、孔洞数量少、气孔直径小、孔壁薄、比容大等基本特征。如表2所示,3 组不同添加物馒头的气孔数量显著多于空白组,但孔洞数量和孔壁厚度均小于空白组,说明α-淀粉酶和λ-卡拉胶能使馒头的内部结构更细腻、更稳定。其中添加λ-卡拉胶/α-淀粉酶复合物组分的气孔数量和气孔面积略低于单独添加酶的组分,气孔直径则接近于单独添加多糖的实验组。总之,λ-卡拉胶的添加弱化了α-淀粉酶的作用效果,该结果与馒头的比容和质构等测试结果一致。

表2 添加不同组分对馒头孔隙分布的影响Table 2 Effects of different additives on the pore distribution of steamed bread
情况较典型的实验组的C-Cell 切片见图6。绿色部分为馒头的大孔洞,深蓝色部分为正常的气孔部分,而较浅的蓝色部分表示气孔分布不均匀。
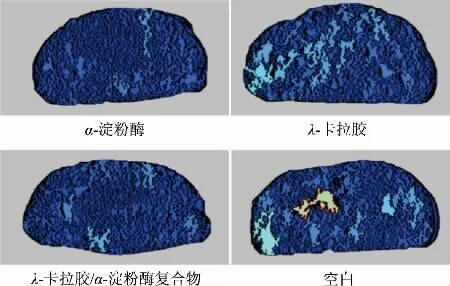
图6 不同添加组分对馒头孔隙组织的影响Fig.6 Effects of different additives on the pore structure of steamed bread
可以看到空白组含有绿色的大孔洞,λ-卡拉胶组有少许浅绿色的小孔洞,而添加α-淀粉酶和λ-卡拉胶/α-淀粉酶复合物的馒头几乎不存在孔洞且气孔分布较均匀。推测原因为复合物中过量的卡拉胶增加了面筋蛋白的持水能力,同时复合物的存在增强了面筋网络的稳定性,进一步说明了α-淀粉酶与λ-卡拉胶/α-淀粉酶复合物均能使馒头内部结构更加紧密细腻。
3 结 语
作者系统研究了α-淀粉酶、λ-卡拉胶和λ-卡拉胶/α-淀粉酶复合物对冷冻面团及其馒头制品性能的影响。结果表明,在冻融过程中α-淀粉酶结构受到较大影响,酶活显著降低,且λ-卡拉胶在冻融过程中通过相互作用显著钝化α-淀粉酶酶活。面团性能测试表明,α-淀粉酶能提升面团的发酵能力,基本消除冷冻对酵母活性和面团结构带来的负面影响,而λ-卡拉胶对面团发酵性能的促进作用有限。λ-卡拉胶/α-淀粉酶复合物对面团的作用劣于α-淀粉酶,推测原因为在冻融循环过程中多糖与酶结合,屏蔽了酶部分作用位点,从而抑制了其活性,但同时λ-卡拉胶也能增加面筋蛋白的持水性,稳定面筋网络。添加一定量的α-淀粉酶、λ-卡拉胶和λ-卡拉胶/α-淀粉酶复合物均能降低馒头的硬度,增加弹性,使馒头气孔分布均匀且减少孔洞。
