知母皂苷A-Ⅲ调控ASAP1-IT1/DNMT3b/YAP1轴在非小细胞肺癌中的作用机制研究*
2022-12-21付相君李艳娜赵文英
付相君,王 梅,张 远,李艳娜,耿 彪,赵文英
(1.皖南医学院,安徽 芜湖 241002;2.皖南医学院第一附属弋矶山医院)
肺癌是恶性肿瘤中最常见类型之一,也是全球癌症相关死亡的主要原因之一。肺癌主要包括小细胞型肺癌(small cell lung cancer,SCLC)和非小细胞型肺癌 (non-small cell lung cancer,NSCLC)两种,其中NSCLC是肺癌的主要类型,约占肺癌总病例的85 %[1]。目前,随着NSCLC治疗策略的发展,如小分子酪氨酸激酶抑制剂和免疫治疗的开发,使得部分NSCLC患者生存率有了明显改善。但是,NSCLC患者总体生存率仍然很低,尤其是转移性NSCLC患者的生存率尚无明显改善[2]。“国之瑰宝”中医药具有多组分、多靶点的药理特点,在NSCLC患者的预后治疗中被认为具有巨大潜力。
中药知母是临床常用药材,具有抗肿瘤、抗抑郁、抗风湿等药理活性,其主要成分为甾体皂苷类和双苯吡酮类化合物,其中甾体皂苷类成分又包括知母皂苷A-Ⅲ(timosaponin A-Ⅲ,TAⅢ)、知母皂苷BI、知母皂苷BII等[3]。TAⅢ是一种糖链主要位于C3位的甾体皂苷[3],其结构如图1所示。TAⅢ在多种癌症中表现出肿瘤细胞抑制活性,如肝癌、直肠癌、肺癌等。研究表明,TAⅢ在NSCLC中可通过抑制多种信号通路(如ERK1/2、Src/FAK 和β-catenin相关通路)来发挥抑癌作用,是治疗NSCLC潜在的候选药物[4-5]。然而,TAⅢ在NSCLC中虽可抑制相关肿瘤进展,但其抗癌作用的机制仍有待阐明。
长链非编码RNA(LncRNAs)参与了多种癌症的发生与发展。ASAP1-IT1、LHFPL3-AS2、CDH5-3:3等在NSCLC的发生过程中被报道具有重要作用[6-8]。其中,ASAP1-IT1可通过调节非小细胞肺癌中的PTEN/AKT和miR-509-3p/YAP1轴,促进细胞的增殖、侵袭和转移[9-10]。LncRNA通过不同机制调控基因表达,研究显示LncRNA可通过与蛋白结合发挥生物学作用[10]。YAP1是多种癌症的癌基因,YAP1蛋白在乳腺癌组织中高表达和低甲基化与DNMT3b相关[11]。本研究证明了TAⅢ在NSCLC中对ASAP1-IT1/DNMT3b/YAP1轴的影响,将有助于阐明TAⅢ对NSCLC作用的相关机制,为开发新型肺癌治疗策略提供理论基础。
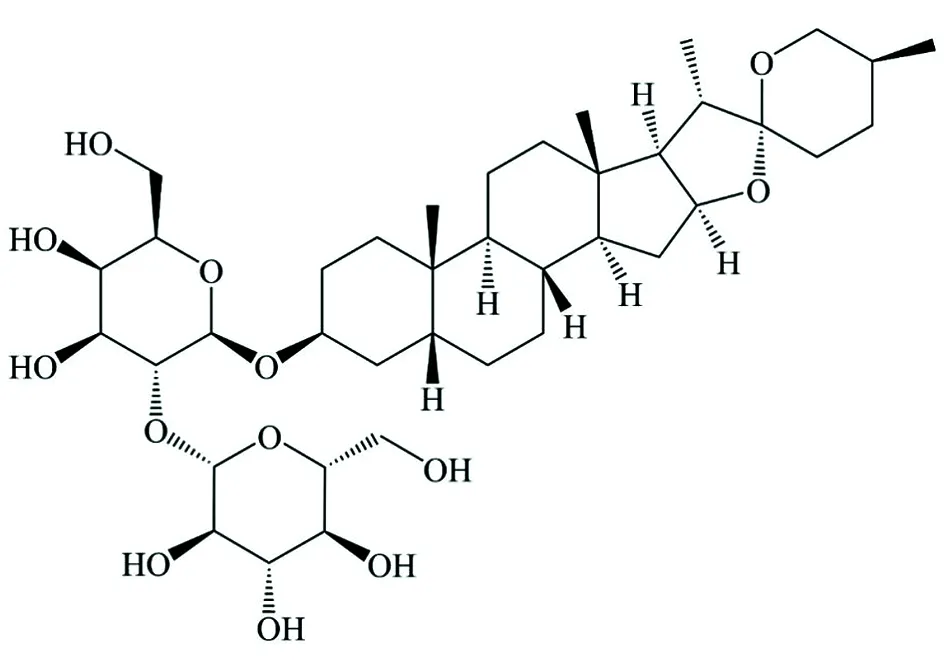
图1 TAⅢ的结构式
1 材料与方法
1.1实验材料 A549细胞(武汉普诺赛生命科技有限公司)、胎牛血清(Gibco)、链霉素(HyClone)、DMEM(HyClone)、转染试剂、6孔板(Corning)、TRIzol试剂(Life Technologies,Rockville) 、PCR引物(安徽通用生物公司)、逆转录试剂盒(Thermo Fisher Science)、SYBR PreMix Ex Taq II染料法荧光定量试剂盒(TaKaRa)、SDS-PAGE凝胶快速配制试剂盒(Beyotime)、RIPA裂解液(Beyotime)、PVDF膜(Thermo Fisher Science)、YAP1、DNMT3b、Bcl2、Bax、caspase3、β-actin抗体、二抗、BCA蛋白浓度测定试剂盒(Beyotime)、CCK-8试剂盒(Beyotime)、96孔板(Corning)、pGEM-T-Easy载体(Promega)、EdU-488细胞增殖检测试剂盒(Beyotime)、RIPTMRNA结合蛋白免疫沉淀试剂盒(Magna)、PierceTM免疫共沉淀试剂盒(Thermo scientific)、Annexin V-FITC细胞凋亡检测试剂盒(Beyotime)。
1.2实验方法
1.2.1临床样本收集 选取2020年4月至2021年9月皖南医学院弋矶山医院收治的65例非小细胞肺癌患者,患者术前均未接受任何治疗。
1.2.2细胞培养和细胞转染 A549细胞在含10 %胎牛血清、1 %链霉素的DMEM中,37 ℃和5 %CO2条件下培养。在对细胞进行转染时,遵循制造商的说明。使用了以下shRNA:sh-ASAP1-IT1:5′-GCUGCGACAAUAGACAUCGGAGUU-3′、sh-YAP1:5′-CUGCCACCAAGCUAGAUAA-3′和sh-DNMT3b:5′-GCCTGGTGATTCTGGGCGAGG-3′。
1.2.3RNA提取及qRT-PCR检测基因表达水平 使用TRIzol试剂提取总RNA,按照逆转录试剂盒说明书将总RNA逆转录成cDNA,使用荧光定量试剂盒进行qRT-PCR。定量逆转录聚合酶链式反应条件为:95 ℃ 5 min,95 ℃ 30 s,60 ℃ 30 s,72 ℃ 1 min,共40个循环。RNA和lncRNA以3-磷酸甘油醛脱氢酶(GAPDH)为内参,miRNA以U6为内参。用2-△△Ct法计算相对表达量。所有检测均为一式4份,每项检测独立进行3次。本研究中使用引物见表1。
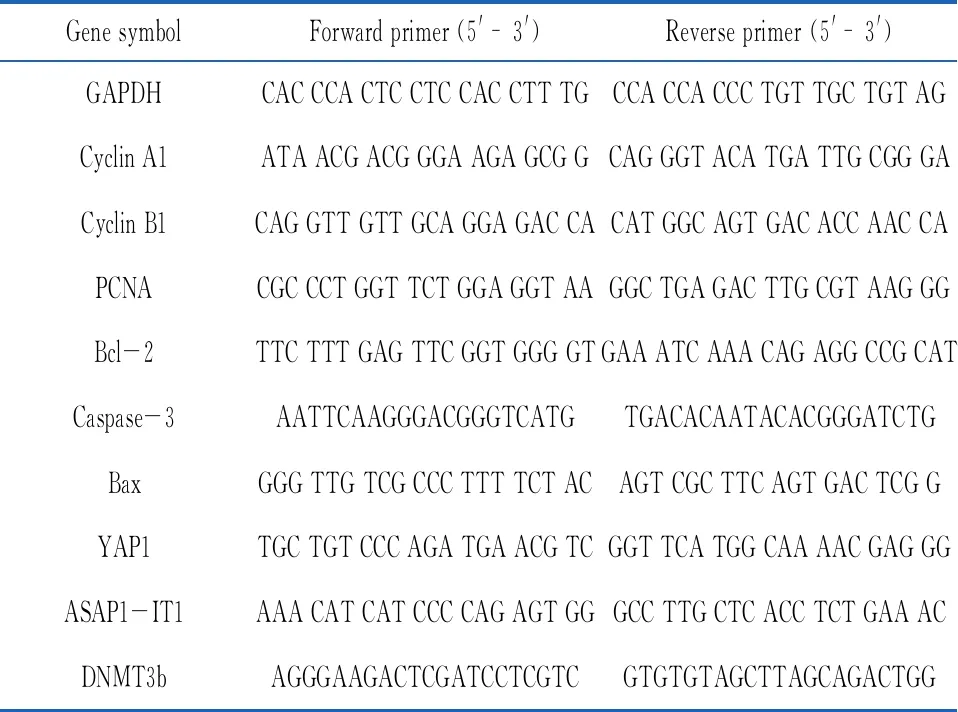
表1 qRT-PCR扩增片段序列
1.2.4CCK-8法检测细胞活力 将A549细胞以每孔4×103个细胞的密度被接种到96孔板中。细胞贴壁后更换培养基, 分别加入 0、5、10、15、20、25和30 μmol/L TAⅢ。作用72 h后, 按照CCK-8试剂盒说明书进行操作,通过酶标仪检测各孔在波长450 nm处的吸光度值。所有检测均为一式4份,每项检测独立进行3次。
1.2.5EDU测定法检测细胞增殖 将对数生长期的细胞接种到96孔板中过夜。用10 μmol/L EDU工作液(1X)孵育4 h。去除工作液后,进行固定及通透。然后按照EDU细胞增殖试剂盒(Beyotime)说明书进行下一步操作。图像在荧光显微镜下可视化。
1.2.6流式细胞术检测细胞凋亡 取1×104个处理后的细胞,离心后弃上清加入5 μL Annexin V-FITC混匀。加入10 μL 碘化丙啶染色液混匀。室温避光孵育15 min,随后置于冰上避光孵育。用流式细胞仪检测。
1.2.7Western blot检测蛋白表达 用RIPA裂解液提取细胞总蛋白后,使用BCA法测蛋白浓度。蛋白样品通过SDS-PAGE凝胶电泳,湿法转膜。PVDF膜用1 %牛血清白蛋白封闭2 h后,在4 ℃下与一抗ASAP1-IT1(1∶1 000)、YAP1(1∶1 000)、Bcl2(1∶1 000)、Bax(1∶3 000)、caspase3(1∶3 000)、β-actin(1∶3 000)、DNMT3b(1∶1 000)孵育过夜。TBST洗膜3次,每次10 min。室温下二抗(1∶5 000)孵育2 h。用Bio-Rad成像系统(Bio-Rad,Hercules)显示蛋白条带。
1.2.8亚硫酸氢盐测序聚合酶链式反应(Bisulfite sequencing PCR,BSP) 设计了硫化测序引物,对扩增产物进行琼脂糖凝胶电泳。将DNA片段克隆到pGEM-T-Easy载体。用BcaBEST测序引物和MS通用引物对阳性重组子进行聚合酶链式反应筛选,然后进行测序。甲基化频率用甲基化胞嘧啶鸟嘌呤(CG)数量与检测到的CG数量之比表示,并取平均值。
1.2.9RNA结合蛋白免疫沉淀法(RNA binding protein immunoprecipitation,RIP) 当A549生长到90 %融合后使用RIPTM RNA结合蛋白免疫沉淀试剂盒用于裂解、抗体孵育及捕获抗原。分为3组,每组两份收集上清液,分别用于蛋白质印迹分析及用qRT-PCR检测ASAP1-IT1的丰度。
1.2.10免疫共沉淀试验(Co-immunoprecipitation,Co-IP) 取对数生长期的A549细胞加入预冷的RIPA缓冲液。离心后将上清液转移到新的离心管中。在1 mL总蛋白中加入100 μL A蛋白琼脂糖珠,在4 ℃下孵育10 min。在离心后取上清液移入新的离心管中除去A蛋白小珠,在500 μL总蛋白中加入抗体,在4 ℃下缓慢摇动抗原-抗体混合物过夜。加入100 μL蛋白A琼脂糖珠捕获抗原-抗体复合体,在4 ℃下孵育过夜。离心后收集琼脂糖珠抗原抗体复合体。去除上清液,琼脂糖珠抗原抗体复合物用预冷的RIPA裂解液洗涤3次,800 μL/次。琼脂糖珠抗原抗体复合体用60 μL 2X样品装载缓冲液悬浮,轻轻混合。将样品煮沸5 min,去除抗原、抗体、珠子,离心,上清液电泳,收集剩余的琼脂糖珠。上清液也可以在-20 ℃下暂时冷冻,上清液煮5 min后再进行电泳。
2 结果
2.1ASAP1-IT1在非小细胞肺癌组织和细胞中的表达及其对细胞生长和凋亡的影响 为探讨ASAP1-IT1在非小细胞肺癌中的作用,采用qRT-PCR检测ASAP1-IT1在人非小细胞肺癌组织及癌旁组织中的表达。qRT-PCR结果显示,ASAP1-IT1在非小细胞肺癌组织中的表达明显高于癌旁组织(图2A)。同时,与人正常肺上皮细胞(BEAS-2B)相比,ASAP1-IT1在NSCLC细胞(A549、Calu-3、PC-9和SPCA-1)中表达上调(图2B)。EdU进一步检测ASAP1-IT1对A549细胞增殖的影响,sh-ASAP1-IT1组细胞增殖率明显降低(图2C)。经流式细胞术和Western Blot分析证实,sh-ASAP1-IT1可通过增加A549细胞的凋亡率上调Bax和Caspase-3的表达,抑制Bcl2的表达而明显增加A549细胞的凋亡率(图2D和图2E)。以上结果提示,在NSCLC的组织及细胞中ASAP1-IT1高表达并促进癌细胞增殖。

图2 ASAP1-IT1的表达和功能
2.2在TAⅢ给药基础上过表达ASAP1-IT1对细胞生长和凋亡的影响 首先检测了48 h的TAⅢ在A549细胞中的半数抑制浓度(IC50),结果表明其IC50为15 μmol/L (图3A)。在随后的实验中,TAⅢ的剂量均为15 μmol/L。发现使用TAⅢ后,ASAP1-IT1的表达显著减少,但过表达ASAP1-IT1可以明显增加ASAP1-IT1的表达(图3B)。接下来检测了A549细胞的增殖情况,发现TAⅢ+oe-NC组细胞增殖率降低,过表达ASAP1-IT1后细胞增殖率略有增加(图3C)。流式细胞术和Western Blot证实,使用TAⅢ可明显增加A549细胞的凋亡率或上调Bax和Caspase-3的表达,抑制Bcl-2的表达。同样,当ASAP1-IT1过表达时,这些指标被逆转(图3D和图3E)。提示TAⅢ可促进A549细胞凋亡且再次验证了ASAP1-IT1可促进NSCLC细胞的增殖。

图3 TAⅢ给药基础上过表达ASAP1-IT1对细胞生长和凋亡的影响
2.3YAP1在非小细胞肺癌组织和细胞中的表达及其对A549细胞生长和凋亡的影响 为了确定YAP1在非小细胞肺癌中的作用,用qRT-PCR检测了YAP1在人非小细胞肺癌标本及癌旁组织中的表达。结果显示非小细胞肺癌组织中YAP1的表达明显高于癌旁组织(图4A)。同时,YAP1在NSCLC细胞(A549、CALU-3、PC-9和SPCA-1)中也表达上调(图4B)。进一步检测YAP1对A549细胞增殖的影响,发现sh-YAP1组A549细胞增殖率明显降低(图4C)。流式细胞术和Western Blot证实,YAP1基因敲除可通过上调Bax和caspase-3的表达,抑制Bcl2的表达明显增加A549细胞的凋亡率。(图4D和图4E)。可得出结论,YAP1在A549细胞中高表达并对其有促增殖作用。
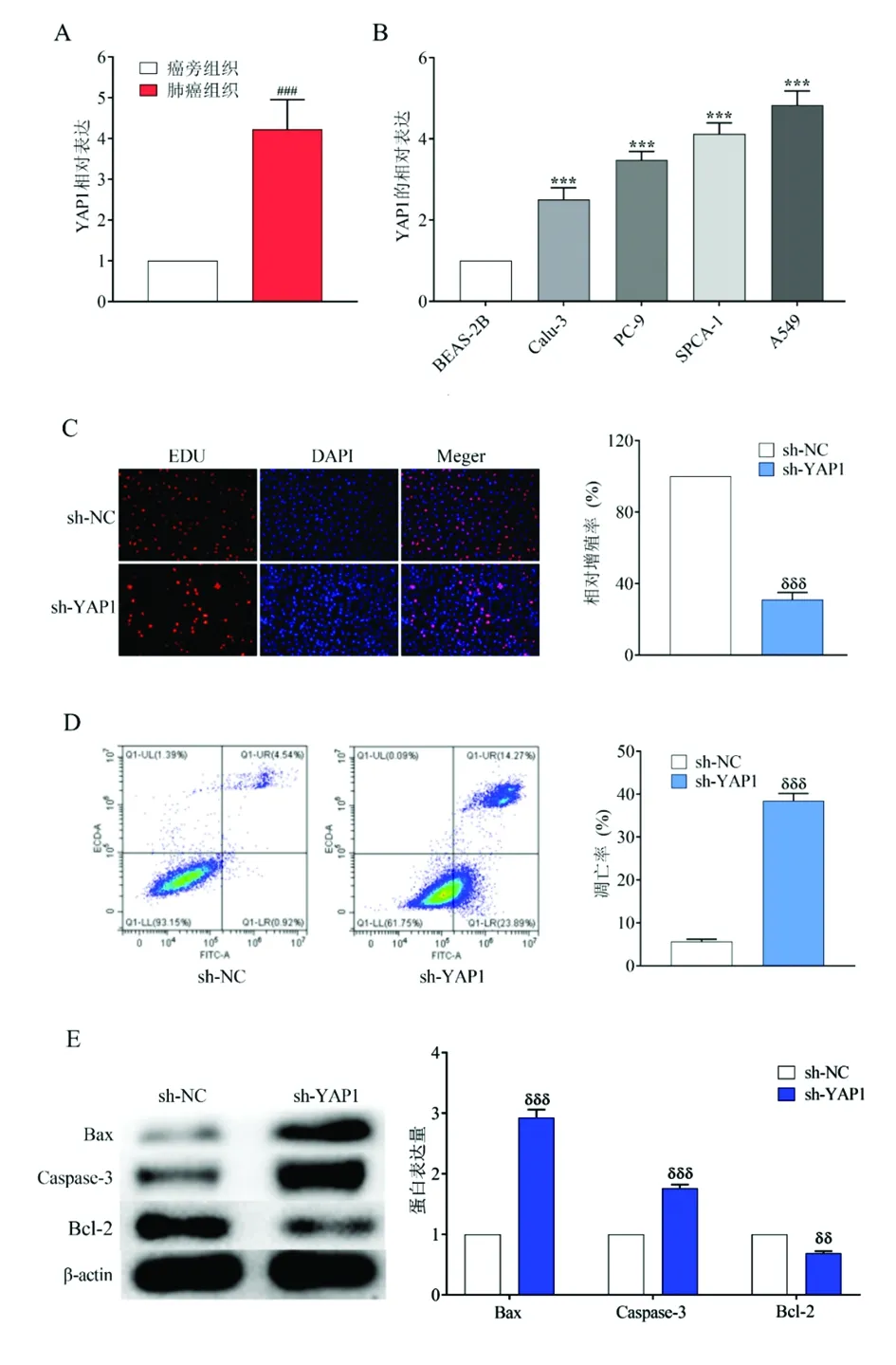
图4 YAP1的作用及表达
2.4在TAⅢ给药基础上过表达YAP1对细胞生长和凋亡的影响 加入TAⅢ后发现YAP1的表达显著减少,但过表达YAP1后可以明显增加(图5A)。为进一步检测YAP1对细胞增殖的影响,检测了A549细胞的增殖,发现过表达YAP1后,TAⅢ抑制的细胞增殖率略有增加(图5B)。此外,流式细胞术和Western Blot证实,当YAP1过表达时,加TAⅢ后细胞凋亡率,Bax、Caspase-3和Bcl-2表达的变化可以逆转(图5C和图5D)。由以上可知,TAⅢ可抑制YAP1的表达。
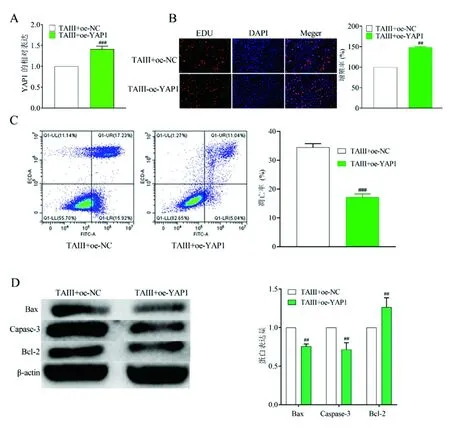
图5 TAⅢ给药基础上过表达YAP1对细胞生长和凋亡的影响
2.5ASAP1-IT1与DNMT3b相互作用增强YAP1表达 在A549中ASAP1-IT1和YAP1的表达呈正相关(图2A和图4A)。沉默ASAP1-IT1后YAP1的表达下降(图6A)。利用Methprimer网站(http://www.urogene.org/methprimer/),在YAP1启动子区域(图6B)确定了一个C-磷酸-G(CpG)岛。随后,证实甲基转移酶抑制剂5-azadc加入后减弱了ASAP1-IT1对YAP1表达的下调作用(图6C)。利用Meth Primer软件设计硫化序列引物(图6D)。BSP分析表明,沉默ASAP1-IT1促进了YAP1启动子的甲基化水平(图6E)。综上所述,ASAP1-IT1调控A549细胞中YAP1的转录表达和甲基化。最后,以RIP和Co-IP实验验证了ASAP1-IT1、DNMT3b和YAP1之间的相互作用关系。RIP结果表明,在与DNMT3b抗体共沉淀的RNA中检测到ASAP1-IT1的存在(图6F和图6G)。Co-IP实验表明DNMT3b和YAP1相结合(图6H)。
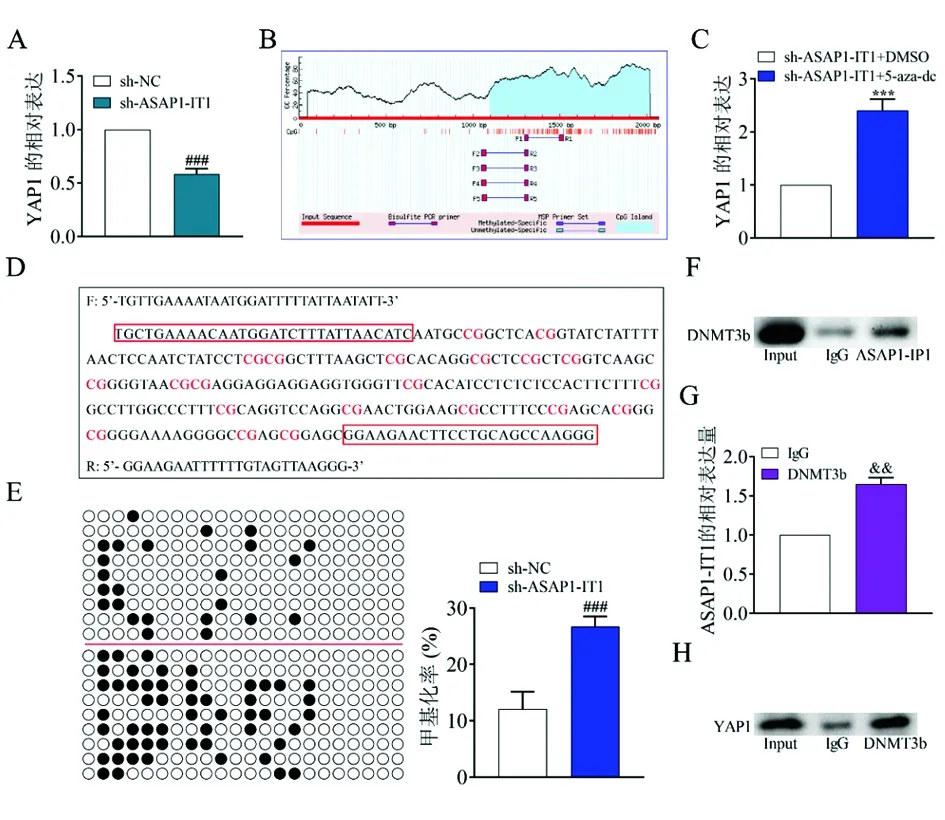
图6 ASAP1-IT1与DNMT3b相互作用增强YAP1的表达
2.6通过DNMT3b介导的YAP1基因敲除ASAP1-IT1在非小细胞肺癌中的肿瘤抑制作用 鉴于A549细胞中ASAP1-IT1基因的敲除可以抑制A549细胞的增殖和促进细胞凋亡,我们推测ASAP1-IT1基因可能通过沉默DNMT3b而导致CpG位点的低甲基化和基因的过度表达。因此,我们推测ASAP1-IT1可能通过调节YAP1在非小细胞肺癌中发挥作用。转染DNMT3b或沉默DNMT3b可提高或降低A549细胞中DNMT3b的mRNA水平(图7A)。通过EDU实验、流式细胞术和qRT-PCR可知,上调DNMT3b或YAP1的表达水平,使EDU阳性细胞数、凋亡率和凋亡指数增加 (图7B-7F)。
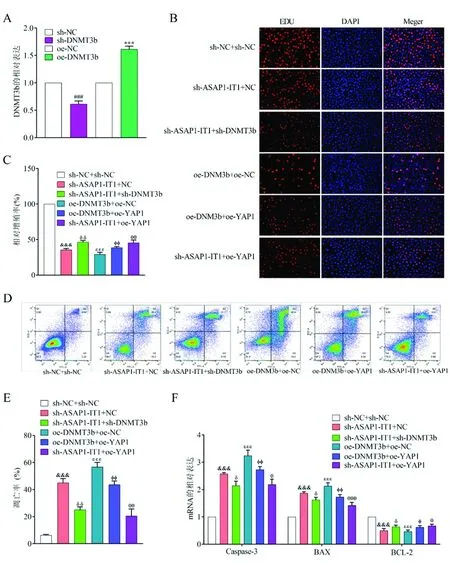
图7 通过DNMT3b介导的YAP1基因敲除ASAP1-IT1在非小细胞肺癌中的肿瘤抑制作用
2.7TAⅢ通过ASAP1-IT1/DNMT3b/YAP1轴对NSCLC的抑制作用 鉴于TAⅢ对A549细胞具有抑制增殖和促进凋亡的作用。因此,我们推测TAⅢ通过调节YAP1在非小细胞肺癌中发挥作用。首先用qPCR检测出在TAⅢ的作用下,A549细胞中DNMT3b的mRNA表达水平增加,但转染组降低(图8A)。然后分别用EDU、流式细胞术和qRT-PCR检测,结果显示DNMT3b水平下调或YAP1表达下调,EDU阳性细胞数和细胞凋亡指数都会发生变化(图8B-D)。
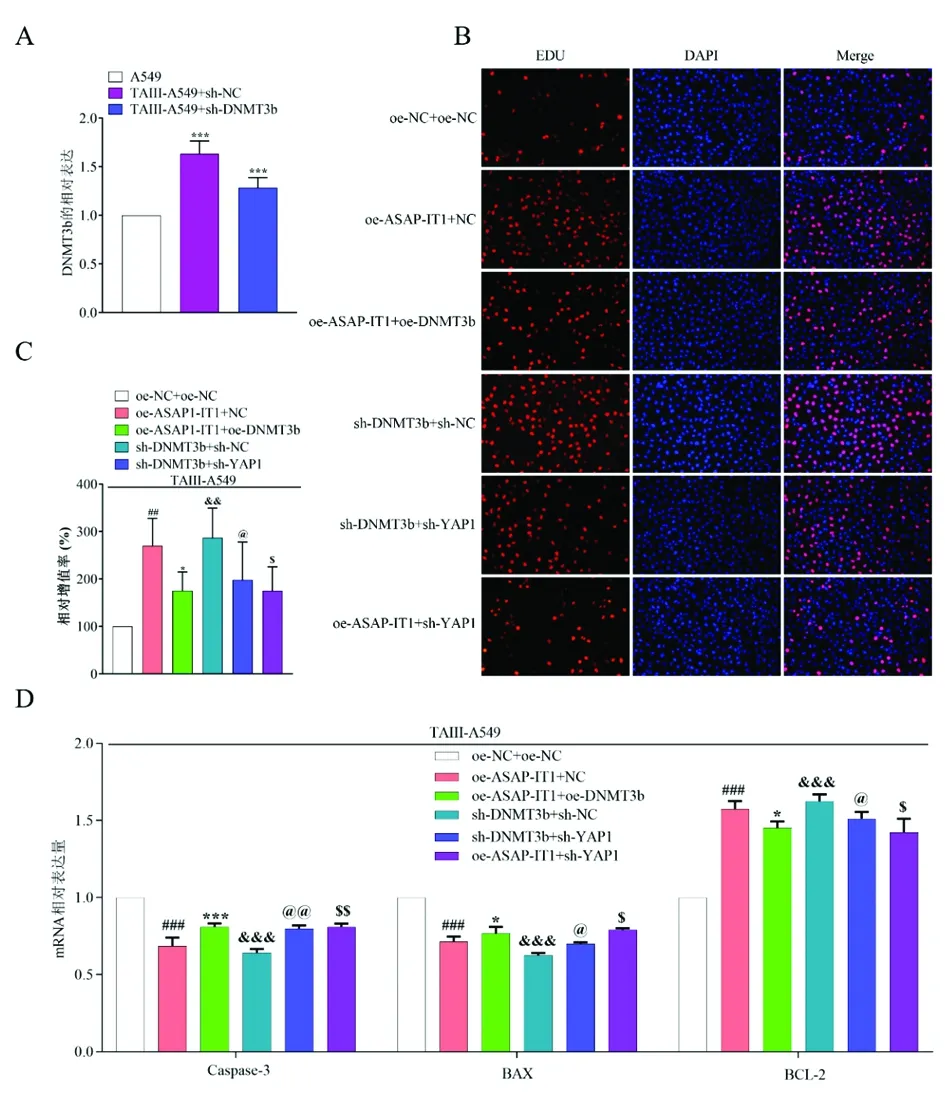
图8 TAⅢ在非小细胞肺癌中发挥肿瘤抑制作用,通过DNMT3b介导的YAP1调节ASAP1-IT1
3 讨论
传统中药知母是百合科植物知母的根茎,其有效成分复杂,药理学活性广泛。TAⅢ是中药知母活性单体成分之一,具有抗肿瘤、抗炎、降血压等多种作用[12]。NSCLC是癌症死亡的主要原因,也是医疗系统重点关注的癌症之一。TAⅢ在NSCLC中具有潜在的抑制活性,本研究就TAⅢ对NSCLC的作用机制进行了研究,报道补充了TAⅢ在NSCLC中的作用机制,再次验证了TAⅢ对NSCLC具有良好的抑制活性。
ASAP1-IT1在癌症的发生与发展中发挥着重要作用,其在卵巢癌中调控Hippo/YAP促进癌症进展,通过调节NSCLC中的miR-509-3p/YAP1轴影响癌细胞的增殖、侵袭和转移[10,13]。近期研究显示,YAP1在乳腺癌组织中的表达与甲基化酶DNMT3b呈负相关。本研究证明了在NSCLC中ASAP1-IT1与YAP1表达上调,可促进癌症进展。同时,RIP、Co-IP及BSP等实验结果证实,AⅢ可通过调节ASAP1-IT1的表达,影响DNMT3b对YAP1的作用,从而产生抑制作用。
综上所述,本研究结果表明,ASAP1-IT1在非小细胞肺癌中表达上调,促进癌细胞增殖,并且与DNMT3b相互作用对细胞凋亡产生影响,证明了在A549细胞中YAP1受到ASAP1-IT1和DNMT3b的调控。TAⅢ处理和沉默ASAP1-IT1通过调节YAP1的甲基化影响A549细胞的增殖和凋亡,证明了TAⅢ通过ASAP1-IT1/DNMT3b/YAP1轴的作用机制,对A549细胞增殖和凋亡产生影响。我们初步认为ASAP1-IT1和YAP1可能是NSCLC的潜在治疗靶点,TAⅢ具有开发为NSCLC治疗药物的潜力。
