雅鲁藏布江下游弧唇裂腹鱼的年龄结构与生长特性
2022-12-21张富斌胡华明曹文宣林鹏程
王 健 张富斌 胡华明 巩 政 曹文宣 林鹏程
(1.中国科学院水生生物研究所, 武汉 430072; 2.水利部长江治理与保护重点实验室, 长江设计集团有限公司, 武汉 430072;3.流域水安全保障湖北省重点实验室, 长江勘测规划设计研究有限责任公司, 武汉 430072; 4.西华师范大学环境科学与工程学院, 南充 637009; 5.枣庄学院生命科学学院, 枣庄 277160)
裂腹鱼亚科鱼类(Schizothoracinae fishes, 以下简称裂腹鱼类)主要分布于青藏高原及其邻近地区,我国有11属97种或亚种[1,2]。裂腹鱼类具有生长缓慢、寿命较高、性成熟晚、繁殖力低等生物学特征, 使其对高原河流水资源的过度开发和环境退化较为敏感[2]。雅鲁藏布江流域横贯西藏, 栖息有多种裂腹鱼类。近年来, 受生境退化、过度捕捞、水电梯级开发和生物入侵等因素影响, 雅鲁藏布江流域以尖裸鲤(Oxygymnocypris stewarti)、拉萨裂腹鱼(Schizothorax waltoni)、巨须裂腹鱼(S.macropogon)和双须叶须鱼(Ptychobarbus dipogon)等为代表的裂腹鱼类资源急剧下降, 高原鱼类的资源保护问题日益突出[2]。因此, 需要对裂腹鱼类的自然种群特征及变动进行系统研究。
雅鲁藏布江下游地处西藏东南部, 主要位于墨脱县境内, 是印度洋湿暖气流进入高原内部的主要通道。区域内雨水丰沛, 河谷深切, 河道比降大, 水流湍急。加之两岸阶地堆积松散, 泥石流、塌方、山体滑坡等自然灾害时有发生[3]。随着经济社会发展和国家“十四五”规划的实施, 雅鲁藏布江下游的水域生态环境将受到一定的干扰。作为雅鲁藏布江下游的优势物种, 弧唇裂腹鱼(Schizothorax curvilabiatus)对于维持下游河流生态系统的结构和功能具有重要的生态价值[1]。但弧唇裂腹鱼的研究仅涉及形态、分类、栖息习性和摄食等方面的简单描述, 缺少详细的基础生物学数据[4,5]。
鱼类生长是其生活史中重要的生物学过程。年龄与生长特征是研究鱼类生物学和种群动态的基础和前提, 获取准确的年龄数据是评估鱼类生长率、种群补充量和死亡率的关键[6—8]。在此背景下对弧唇裂腹鱼的年龄和生长特征进行分析, 以揭示物种的生长特性和种群生长潜力, 为变化环境下雅鲁藏布江下游鱼类资源的保护提供科学依据。
1 材料与方法
1.1 样本采集和年龄材料获取
2015年12月至2016年11月逐月在雅鲁藏布江下游墨脱段开展鱼类采集工作(图1), 获取弧唇裂腹鱼928尾。采样网具为定置刺网(网目4—8 cm)、撒网(网目6 cm)和地笼(网目1 cm)等(调查采样均在渔政主管部门备案, 禁用网具取得捕捞许可)。样本获取后测定体长、体重、内脏重和性腺重(长度精确至l mm, 重量精确至0.1 g)。根据性腺发育判断样本性别。取出微耳石、鳃盖骨和脊椎骨, 耳石用清水洗净后放入离心管中保存, 脊椎骨和鳃盖骨置于自封袋中冰冻保存。

图1 雅鲁藏布江下游弧唇裂腹鱼的采样区域Fig.1 Sampling area of S.curvilabiatus in the lower reach of Yarlung Zangbo River
1.2 年龄材料处理
耳石用指甲油将微耳石包埋并固定于载波片一边, 使其凸面朝上, 随后用800—2500#的水磨砂纸依次打磨, 再用抛光纸抛光。打磨时, 不时地移至显微镜下观察耳石轮纹。当看到耳石中心较为清楚时, 利用丙酮溶解指甲油, 将耳石翻面包埋, 用相同的方法打磨和抛光直至耳石中心和外周轮纹都较清晰为止[9]。
脊椎骨将脊椎骨置于沸水中加热约10min,去除附着的软组织后置于1% 的H2O2中浸泡24h。晾干后用剪刀将脊椎骨从中间剖开, 置于解剖镜下观察(入射光), 并用二甲苯透明处理。
鳃盖骨将鳃盖骨置于沸水中加热约1min,去除附着的软组织, 再将其置于1%的 H2O2中浸泡24h后晾干, 最后在解剖镜下观察(透射光)。
1.3 年龄材料鉴定和比较
年龄的确认和计数方法参照殷名称等[10,11]。同一鉴定者对弧唇裂腹鱼的耳石磨片, 脊椎骨和鳃盖骨进行两次独立读数, 两次读数间隔3周以上。并对耳石、脊椎骨和鳃盖骨的年轮清晰度进行5等级评分: 1为非常好, 2为好, 3为一般, 4为很差, 5为难以辨认[12](表1)。

表1 年龄材料轮纹清晰度等级划分表Tab.1 Readability level of annuli on different calcified structures
1.4 数据分析
体长体重关系通过幂函数(W=aSLb)对弧唇裂腹鱼的体长体重关系进行回归分析, 式中W为体重(g),SL为体长(mm), a, b为常数, a为生长的条件因子, b为幂指数系数。同时将体长体重数据进行对数化处理, 采用协方差分析(ANCOVA)来检验不同性别体长体重的关系是否具有显著性差异[13]。并对异速生长指数(b)和3的差异性进行t检验[14],以此推测弧唇裂腹鱼是否为匀速生长类型鱼类。
生长方程统计雌雄个体不同年龄组实测体长的平均值和范围。采用von Bertalanffy生长方程Lt=L∞[1-ek(t-t0)]拟合弧唇裂腹鱼的生长特征,式中Lt为t龄时的实测体长,L∞为渐进体长,k为生长曲线的平均曲率;t0为理论上体长和体重等于零时的年龄。表观生长指数(Ø)被用于同属鱼类生长性能的比较,计算公式如下:Ø=1gk+2lgL∞[15]。数据用平均数±标准差表示, 统计显著性为α=0.05。统计分析采用SPSS 16.0和Origin Pro 2016, 图像分析和处理采用Photoshop 2020。
2 结果
2.1 年龄材料特征
随机选取了部分样品的三种年龄材料进行加工处理, 最终对耳石、脊椎骨和鳃盖骨都成功鉴定年龄的有236尾(体长112—508 mm, 鉴定结果见表2)。三种年龄材料上均存在年轮, 但年轮特征各异。

表2 弧唇裂腹鱼各个年龄组的样本数和体长Tab.2 Number and standard length of S.curvilabiatus at different ages
微耳石有清晰轮纹。经过打磨后的微耳石中心有个核区, 在透射光下呈现出明显的由宽带和窄带(或明带和暗带)组成的轮纹。年轮间距在耳石上呈规律性分布, 靠近核区的轮纹的间距较宽大, 越往边缘延伸轮纹间距越来越窄(图2)。
脊椎骨为双凹型, 在解剖镜下可观察到明暗相间的轮纹, 轮纹呈同心椭圆排列。椎骨中央具一小孔, 小孔周围薄而透明, 轮纹少且细, 较难确认首轮的位置(图2)。
弧唇裂腹鱼的鳃盖骨呈不规则扇形, 鳃盖骨上的年带由乳白色宽带和暗黑色窄带组成。窄带和宽带之间有明显的界限, 这一界限为年轮标志(图2)。

图2 弧唇裂腹鱼不同年龄材料轮纹特征 (圆点表示年轮)Fig.2 Annuli characteristics of different calcified structures of S.curvilabiatus (dots show annuli)
耳石、脊椎骨和鳃盖骨的清晰度评分见表3。耳石轮纹最为清晰, 1、2等级的耳石占总数72.03%;脊椎骨次之, 1、2等级的耳石的脊椎骨占总数58.90%; 鳃盖骨上年轮清晰度最差, 1、2等级的鳃盖骨只有47.88%。综上可见, 耳石对弧唇裂腹鱼的年龄鉴定效果最佳, 脊椎骨次之, 鳃盖骨最差。

表3 弧唇裂腹鱼不同年龄材料的清晰度评分Tab.3 Distribution of readability scores for different calcified structures of S.curvilabiatus (n=236)
从图3可知, 耳石磨片两次读数一直保持较高的吻合率, 鳃盖骨差异性较大。在 9龄之前, 脊椎骨两次年龄读数能保持很好的一致性, 但随年龄的递增, 鉴定结果出现差异。整体来看, 弧唇裂腹鱼耳石精确性都要优于脊椎骨和鳃盖骨。
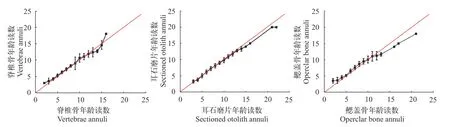
图3 同一观察者两次年龄鉴定结果偏差图Fig.3 Age bias plots for pairwise age estimates from two independent readings
由表4可以看出, 三种年龄材料所鉴定的平均年龄存在显著性差异(P<0.05)。用耳石作为年龄材料所鉴定的平均年龄最高(8.87龄), 显著高于脊椎骨(7.62龄)和鳃盖骨(7.81龄)所鉴定的平均年龄(P<0.05)。10龄以下和10龄以上样本分开统计时, 在1—10龄样本中, 耳石鉴定的平均年龄(7.01龄), 显著高于鳃盖骨(6.60龄)和脊椎骨(6.37龄)所鉴定的平均年龄 (P<0.05)。大于10龄时, 脊椎骨(11.63龄)和鳃盖骨(12.0龄)所鉴定的平均年龄较为接近, 但都显著低于耳石所鉴定的平均年龄(13.2龄;P<0.05)。

表4 弧唇裂腹鱼不同年龄材料所鉴定的平均年龄Tab.4 Mean values of age estimates from different calcified structures of S.curvilabiatus
2.2 体长分布
弧唇裂腹鱼的体长为48—508 mm, 均值为(227.4±111.4)mm。65.30%的个体体长集中在100—300 mm (图4)。在 928尾样本中, 雌性395尾,体长为85—508 mm; 雄性474尾, 体长为81—455 mm;性别未辨的个体59尾, 体长为48—170 mm。统计分析表明, 雌鱼和雄鱼的体长分布存在显著性差异(Kolmogorov-SmirnovZ检验,Z=2.397,P<0.05)。

图4 弧唇裂腹鱼体长分布图Fig.4 Frequency distribution of the standard length groups of S.curvilabiatus
2.3 年龄结构
在928尾样本中, 因极少数耳石本身就不具备轮纹或者耳石在打磨处理过程中有磨损, 故成功鉴定微耳石年龄的样本为916尾。其中, 雌鱼年龄组样本量为389尾,年龄为2—43龄; 雄鱼年龄组样本量为470尾,年龄为2—23龄, 性别未辨个体的年龄组样本量为57尾,年龄为1—4龄。雌雄个体大于15龄的高龄组均少见,年龄分布主要集中在15龄以下。优势年龄组为3—5龄, 占全部样本的44%(图5)。不同性别不同年龄组的样本数和平均体长见表5。

图5 弧唇裂腹鱼年龄组成图Fig.5 Frequency distribution of estimated age groups of S.curvilabiatus
2.4 生长特征
体长体重关系将性别未辨个体、雌性群体、雄性群体和种群总体的体长和体重数据分别进行拟合(各方程拟合时的样本量为各群体采集到的样本数量; 图6), 结果表明弧唇裂腹鱼体长体重关系符合幂函数公式。
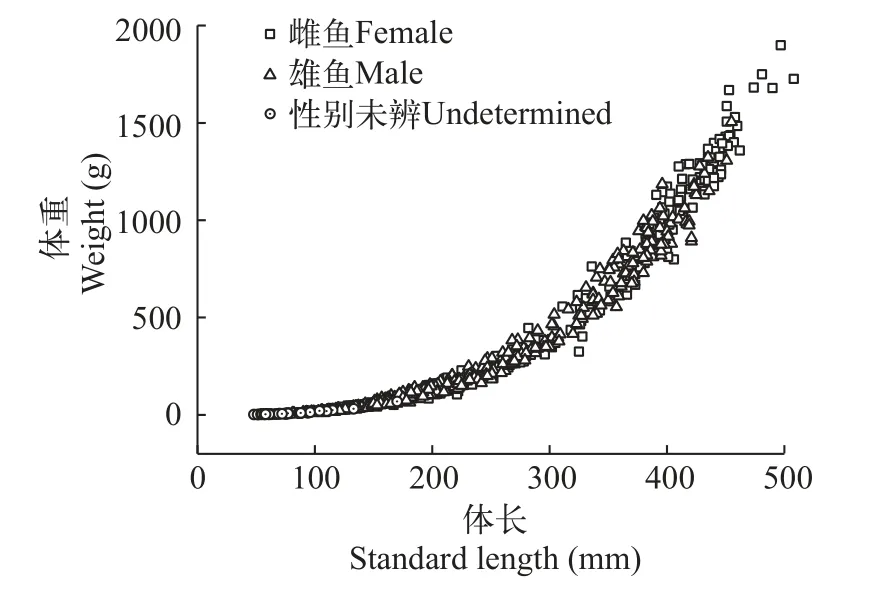
图6 弧唇裂腹鱼体长体重关系图Fig.6 Length-weight relationship of S.curvilabiatus
性别未辨个体:W=3.122×10–5SL2.855,R2=0.983,n=59;
雌性群体:W=2.500×10–5SL2.914,R2=0.993,n=395;
雄性群体:W= 2.114×10–5SL2.944,R2=0.990,n=474;
种群总体:W=2.279×10–5SL2.931,R2=0.993,n=928。
弧唇裂腹鱼幼鱼的b值小于3, 为异速生长阶段,表明此阶段体长生长快于体重生长。经协方差分析, 显示弧唇裂腹鱼雌鱼和雄鱼间的体长体重关系差异并不显著(F=0.833,P>0.05), 故将其合并为种群总体的幂函数方程。经t检验, 表明种群总体b值与3差异显著(t=8.788,P<0.05), 表明弧唇裂腹鱼为异速生长鱼类。
生长方程经检验, 2—4龄的雌鱼和雄鱼,其同年龄组平均体长没有显著性差异(two-sample,P>0.05)。von Bertalanffy生长方程拟合时, 将性别未辨个体中2—4龄的样本数据(n=15)分别纳入雌鱼(n=389)和雄鱼(n=470)的年龄数据中, 以获取更为准确的t0值[12](表5)。根据实测体长拟合的体长生长方程如下:

表5 弧唇裂腹鱼不同年龄组的样本数和平均体长Tab.5 Number of specimens and range of standard length at age of S.curvilabiatus
雌鱼:Lt=590.2[1–e–0.096(t–0.282)](R2=0.932,n=404)
雄鱼:Lt=575.3[1–e–0.090(t+0.011)](R2=0.918,n=485)
将体长体重关系式代入体长生长方程, 得到体重生长方程:
雌鱼:Wt=3016.8[1–e–0.096(t–0.282)]2.931
雄鱼:Wt=2798.9[1–e–0.090(t+0.011)]2.931
雌鱼和雄鱼群体的表观生长指数(Ø)分别为4.5243和4.4740。
弧唇裂腹鱼的年龄和体长呈现一定的关系(图7)。在低于10龄时, 相同年龄雌雄性的体长非常相近,之后随着年龄的增长, 差异越来越大, 相同年龄的雌性体长大于雄性。

图7 弧唇裂腹鱼的年龄与体长关系图Fig.7 The relationship between age and standard length of S.curvilabiatus
生长速度和加速度将弧唇裂腹鱼的生长方程经过一阶求导得到其生长速度方程, 再通过二阶求导获得其生长加速度方程。
雌鱼:dL/dt=56.66e–0.096(t–0.282)
dL2/dt2= −5.44e−0.096(t–0.282)
dW/dt=848.85e–0.096(t–0.282)[1–e–0.096(t–0.282)]1.931
dW2/dt2=81.49e–0.096(t–0.282)[1–e–0.096(t–0.282)]0.931[2.931e–0.096(t–0.282)–1]
雄鱼:dL/dt=51.78e–0.090(t+0.011)
dL2/dt2=–4.66 e–0.090(t+0.011)
dW/dt=738.33 e–0.090(t+0.011)[1–e–0.090(t+0.011)]1.931
dW2/dt2=66.45e–0.090(t+0.011)[1–e–0.090(t+0.011)]0.931[2.931e–0.090(t+0.011)–1]
弧唇裂腹鱼雌鱼体长增长的速度和加速度的变化趋势与雄鱼相似(图8a和8b)。随着年龄的增长, 雌鱼和雄鱼的体长增长速度均呈递减趋势, 且递减速度逐渐减缓, 无限接近于0; 雌鱼和雄鱼体长增长的加速度随着年龄的增长呈递增趋势, 递增速度逐渐减缓, 无限接近于0而小于0。
弧唇裂腹鱼雌鱼的体重加速度变化趋势与雄鱼相似(图8c和8d), 均呈先升后降的趋势。当体重生长加速度为0时, 体重生长速度达到最大值, 此为该鱼的体重生长拐点。弧唇裂腹鱼雌鱼拐点年龄(ti)为11.5龄, 雄鱼拐点年龄(ti)为11.9龄, 对应的体长分别为388.8和379.0 mm; 对应的体重分别为887.9和823.8 g。

图8 弧唇裂腹鱼的生长速度和生长加速度Fig.8 Growth rate and acceleration of S.curvilabiatus
3 讨论
3.1 年龄鉴定
已有的研究显示, 鳞片、脊椎骨、耳石、鳍条、匙骨和鳃盖骨等钙化组织是裂腹鱼类年龄鉴定的常用材料[16—24]。不同年龄材料轮纹清晰的程度因研究物种、个体生长差异而不同。刘艳超等[23]用微耳石、星耳石、脊椎骨、鳃盖骨、臀鳞、胸鳞、侧线鳞和背鳞8种年龄材料对西藏拉萨河上游双须叶须鱼的年龄特征进行了比较, 认为微耳石最适宜用于双须叶须鱼的年龄鉴定。Sabah和Khan[24]-利用耳石和脊椎骨等年龄材料对克什米尔河谷三种裂腹鱼的年龄和生长进行比较研究, 结果表明耳石用于中唇裂腹鱼S.curvifrons和S.niger的年龄鉴定较脊椎骨更准确, 而用脊椎骨鉴定扁嘴裂腹鱼S.esocinus的年龄比耳石准确。Ma等[16]和Zhou等[22]的研究结果显示, 脊椎骨鉴定两种裂腹鱼属鱼类的年龄时, 低龄鱼的鉴定结果与耳石差异不大; 而在鉴定高龄鱼时, 其结果显著低于耳石; 相比较耳石和脊椎骨, 鳃盖骨的轮纹比较密集难以区分, 且对于高龄鱼而言, 干扰纹偏多, 容易造成误判。
在本研究中, 鳃盖骨轮纹清晰度不及耳石和脊椎骨, 其年轮读数也显著低于耳石和脊椎骨; 而脊椎骨在低龄鱼中(低于10龄)与耳石的鉴定结果相当, 但在高龄鱼中(10龄以上)年龄判读结果的准确率和辨识度均较耳石差。综上所述, 耳石是鉴定裂腹鱼类年龄较为合适的材料[16,19—23]。采用体外标记和氯化四环素荧光标记方法也表明, 用耳石鉴定鱼类年龄比鳞片更为准确, 尤其是生长缓慢的长寿命鱼类[6]。主要原因在于耳石生长独立于机体生长, 在慢生长和高龄个体中, 耳石比其他骨质材料生长更快, 且不会存在轮纹的重吸收现象, 故其更能如实反映周期性的季节生长, 并以规律性轮纹呈现为年龄[2,25]。
3.2 生长特性
由于高原特殊的生态环境, 裂腹鱼类往往都具有较高的寿命, 且雌性个体年龄大于雄性个体。已有的报道显示, 雅鲁藏布江异齿裂腹鱼雌雄个体的最大年龄分别为50龄和40龄[26], 双须叶须鱼雌雄个体的最大年龄分别为45龄和24龄[17], 拉萨裂腹鱼雌雄个体的最大年龄分别为40龄和37龄[22]。在本研究中, 弧唇裂腹鱼雌鱼和雄鱼的最大年龄分别为43龄和23龄, 与上述研究结果接近。
生长系数k和表观生长指数(Ø)都是评估鱼类种群生长潜力的关键参数[2]。Branstetter[28]将生长系数k分为三类: 0.05—0.10属生长缓慢型, 0.10—0.20属均速生长型, 0.20—0.50属快速生长型。生长方程的拟合结果显示, 弧唇裂腹鱼的生长系数k值低于0.1, 其中雌性为0.096, 雄性为0.090; 雌性和雄性群体的表观生长指数(Ø)分别为4.5243和4.4740。上述参数与Zhou等[22]、Ma等[26]、朱秀芳和陈毅峰[29]、Li and Chen[30]和Huo等[31]对雅鲁藏布江中上游裂腹鱼类的研究结果一致(表6)。同时, 弧唇裂腹鱼的最大年龄和渐进体长L∞较高(雌性为590.2 mm, 雄性为575.3 mm)。综上可以判断, 弧唇裂腹鱼为生长缓慢, 寿命较长的鱼类, 属于典型的K-对策选择者[32]。
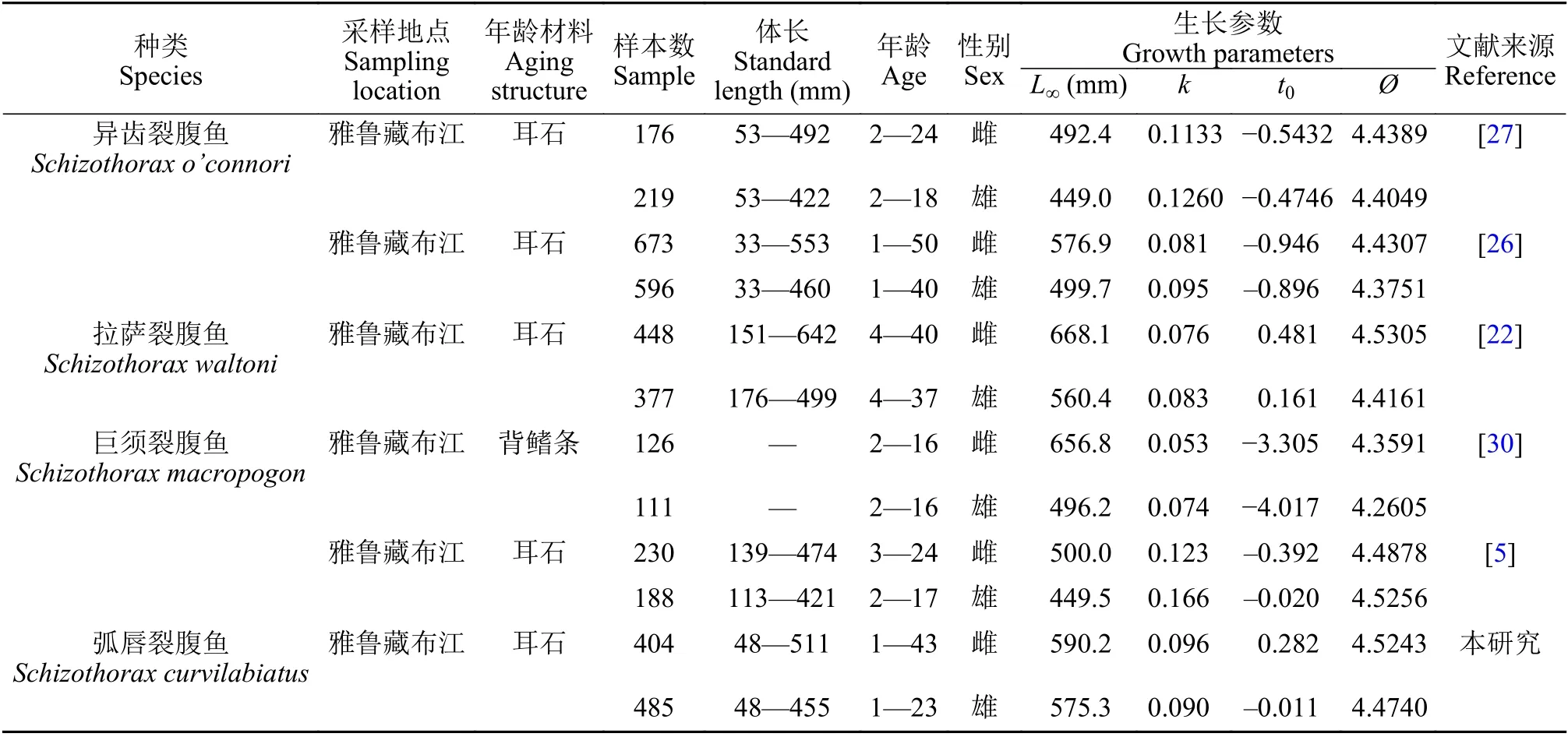
表6 文献中雅鲁藏布江裂腹鱼属鱼类生长特性对比Tab.6 Comparison of growth characters of Schizothorax fishes in the Yurlung Zangbo River in different studies
鱼类个体生长是内源因子和外源因子共同作用的结果。除物种间的遗传差异外, 食物资源、温度、光照及其他环境因子也会影响鱼类的个体生长[2]。由于入侵水域营养生态位的空缺, 加之适口饵料丰度较大, 抚仙湖的黄颡鱼种群表现出很好的生长潜力[33]。近年来, 乌伦古湖饵料生物的数量和生物量较1987年均大为减少, 河鲈(Perca fluviatilisLinnaeus)的喜好饵料摇蚊幼虫生物量仅为1987年的1/10, 从而导致河鲈营养状况不佳, 影响河鲈生长速度[34]。雅鲁藏布江下游河谷深切, 水流湍急,流速可达16 m/s, 加之地质活动频繁, 滑坡和泥石流时有发生, 导致干流河道水生生物饵料资源相对匮乏[1,35,36]。故推测弧唇裂腹鱼生长缓慢是对食物资源匮乏等恶劣环境的一种长期适应[2,4,35]。此外, 同为植食性的裂腹鱼亚属鱼类, 弧唇裂腹鱼的表观生长指数(Ø)高于异齿裂腹鱼, 可能与雅鲁藏布江下游海拔较低, 水温相对较高有关。水温较高使弧唇裂腹鱼具有较高的新陈代谢速率, 能量转换效率可能高于异齿裂腹鱼。弧唇裂腹鱼的表观生长指数与谢从新等[5]对拉萨裂腹鱼和巨须裂腹鱼的研究结果较为接近, 但高于朱秀芳和陈毅峰[30]对巨须裂腹鱼的研究结果, 可能与所用年龄材料不同有一定关系(表6)。
3.3 弧唇裂腹鱼的保护管理
研究指出, K-对策选择鱼类对环境变化非常敏感, 种群一旦衰退, 其资源恢复速度缓慢[32]。近年来, 人为活动对雅鲁藏布江下游河流环境的干扰逐渐加大。一方面随着旅游开发、人口经济的快速增加、墨脱河段的渔业捕捞强度逐步增大, 导致弧唇裂腹鱼种群以3—5龄为主, 种群低龄化现象突出;另一个方面随着国家“十四五”规划的深入推进, 涉水工程及沿江公路的实施将改变雅鲁藏布江下游的河流形态、水文情势及水体理化性质, 进而对弧唇裂腹鱼等土著鱼类的栖息产生不利影响。
针对上述问题, 并结合弧唇裂腹鱼的生物学特性及流域鱼类的保护现状, 提出了以下措施与保护建议: (1)控制渔业捕捞是保护鱼类资源的重要手段。弧唇裂腹鱼拐点年龄为11龄, 对应的体长体重分别为380 mm和820 g。为此, 建议将其起捕规格限制在800 g以上, 同时加快完善渔业资源管理体系建设, 严厉打击非法捕捞。(2)栖息地保护是物种保护最有效的措施之一。雅鲁藏布江下游水域生态学的研究基础薄弱, 建议加强弧唇裂腹鱼产卵场、越冬场和索饵场等重要生境的调查与研究, 在此基础上选择合适的江段建设弧唇裂腹鱼水产种质资源保护区, 加强其种质资源的保护。(3)目前雅鲁藏布江中上游异齿裂腹鱼、拉萨裸裂尻鱼(Schizopygopsis younghusbandi)等主要鱼类的人工繁殖已获得成功, 形成了较为系统的规模化人工繁育技术体系[2]。相比之下, 雅鲁藏布江下游鱼类的人工繁育技术尚不成熟, 建议加强弧唇裂腹鱼的人工繁殖及苗种批量培育等关键技术的研究, 发展可持续的人工养殖, 减少对弧唇裂腹鱼野生种群资源的依赖。
