西藏拉鲁湿地和茶巴朗湿地外来鱼类群落结构及变动趋势
2022-12-21隋晓云孙欢欢贾银涛陈毅峰
朱 仁 隋晓云* 孙欢欢,2 贾银涛 冯 秀 陈毅峰,2*
(1.中国科学院水生生物研究所, 武汉 430072; 2.西藏大学理学院, 拉萨 850000)
湿地具有涵养水源、调蓄洪水及保持物种多样性等多方面的生态功能和作用[1]。全球湿地资源约有87%正在退化或丧失, 湿地生态系统已成为全球遭受破坏最为严重的生态系统之一[2]。与其他地区相比, 青藏高原湿地生态系统的初级生产力低下,群落结构简单且脆弱, 对全球气候变化和人类活动的反应极其敏感[3,4]。随着青藏高原地区社会和经济的快速发展, 湿地生态系统受人类活动的影响也在不断加剧, 外来生物的引入(动植物和病原菌等)[5]是其中重要的人类影响。
为了提高水产品产量和当地人民生活水平, 鱼类作为水产贸易品种或养殖对象引种至中国各地[6,7]。最早仅由英国殖民者将亚东鲑(Salmo trutta fraio)引入西藏[8]。至20世纪90年代起, 研究者发现鲤(Cyprinus carpio)出现在西藏部分城市公园和周边河流中[9,10]。进入21世纪后, 越来越多的外来鱼类, 包括黄鳝(Monopterus albus)、草鱼(Ctenopharyngodon idella)、鲫(Carassius auratus)、麦穗鱼(Pseudorasbora parva)、鲇(Silurus asotus)和小黄䱂(Micropercops swinhonis)等出现在西藏自然水体中[10—15]。这些外来鱼类被引入后, 可能通过竞争(栖息地和食物)、捕食、携带新的病原体、改变生境和栖息地结构等多种方式对当地土著物种造成严重威胁[6]。近年来, 有关高原外来鱼类的研究包括外来鱼类的引入、生态效应和入侵风险评估等方面已受到学界及公众的广泛关注[16—19]。然而, 有关西藏城市及周边重要湿地外来鱼类群落结构及变化趋势仍缺乏较深入的研究。
因此, 本研究实地调查了西藏拉鲁湿地和茶巴朗湿地外来鱼类的现状, 研究其群落结构, 并结合历史资料分析其变动趋势, 旨在为青藏高原湿地生态系统的保护及外来物种防控管理提供基础资料和科学依据。
1 材料与方法
1.1 研究区域
拉鲁湿地位于拉萨市(91°03′41″—91°06′48″E,29°39′25″—29°42′08″N), 总面积6.2 km2, 为国家级湿地保护区[20,21]。茶巴朗湿地是拉萨河下游的主要湿地(90°49′20″—90°50′30″E, 29°22′30″—29°22′59″N), 总面积0.2 km2, 属市级重点湿地生态功能保护区[22,23]。两个湿地均附属拉萨河流域, 是西藏自治区内典型的高寒草甸沼泽湿地, 处于温带半干旱季风气候区(图1)。年平均降水量为439.8 mm (降水主要集中在7—9月); 气温较低, 范围为16℃(6月或7月)至−1.6℃(1月)[21]。

图1 西藏拉鲁湿地和茶巴朗湿地示意图Fig.1 Map of Lhalu Wetland and Chabalang Wetland in Tibet Autonomous Region
1.2 调查方法
2019年7月、2020年8月和2021年4月在拉鲁湿地和茶巴朗湿地开展鱼类调查。为保证充分收集不同栖息水层的鱼类样品, 使用刺网(长10 m×高0.5 m,网目3 cm和8 cm)和地笼(长8 m×宽0.25 m×高0.2 m,网径0.5 cm)组合渔具, 在样点放置12h收集渔获物。对收集的鱼类标本参考朱松泉[24]、陈宜瑜[25]、乐佩琦[26]、褚新洛等[27]、伍汉霖[28]、李思忠和张春光[29]等书籍进行分类鉴定, 采用中国生物物种名录(2022版)数据库(http://www.sp2000.org.cn/)所使用的分类系统, 统计渔获物的全长(精确到0.1 cm)和体重(精确到0.01 g)等生物学信息。
1.3 指数分析
重要性指数采用Pinkas[30]相对重要性指数(Index of Relative Importance,IRI)来说明外来鱼类群落中各物种的现状。计算公式如下:

式中,N%为某外来鱼类个体数占所有外来鱼类渔获物总个体数的比例,W%为某外来鱼类重量占所有外来鱼类渔获物总重量比例,F%为某外来鱼类出现的年度数占3个年度(2019—2021年)的比例。IRI>1000为优势种。
功能多样性指数鱼类的功能性状划分采用陈康等[31]方法, 结合实地调查和文献资料以及Fishbase网站(http://www.fishbase.org)上的记述对采集到的鱼类物种识别其功能性状, 主要包含营养级、栖息水层(底层、中层和上层)、食性(底栖动物食性、浮游生物食性、肉食性和杂食性)、运动方式(静止或运动)、身体形状(细长形、梭形、长方形、椭圆形)、感觉触须(有或无)、最大个体尺寸(依据全长区间划分, 等级1: <10 mm、等级2:10—20 mm、等级3: 20—40 mm、等级4: 40—80 mm、等级5: >80 mm)和种群增长率(中或低)。运用凸壳体积法(Convex hull volume)计算鱼类群落的功能多样性(Functional diversity,FD), 包括功能丰富度(Functional richness,FRic)、功能均匀度 (Functional evenness,FEve)和功能分散度(Functional disperse,FDis)3个指数。数据分析在R 4.1.2软件中实现,功能多样性选择Gower距离矩阵在FD包中进行计算[32,33]。计算公式如下:
(1)功能丰富度FRic: 由功能性状空间内形成的最小凸多边形体积来计算[34]。
(2)功能均匀度FEve:
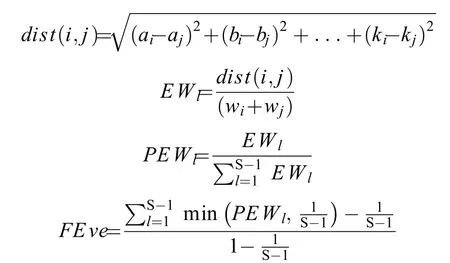
式中,a—k表示多维性状空间中物种i和j的8个功能性状;dist(i,j)表示物种i和j之间的欧式距离;EWl为分支长度;wi和wj分别为物种i和j的相对丰度;PEWl为分支长权重; S为物种数。
(3)功能分散度FDis:

式中,aj为物种j的相对多度,xij为物种j第i个性状值。zj是物种j到重心c的加权距离。
1.4 数据收集和处理
鱼类数据收集采用中国知网(CNKI, http://www.cnki.net)和汤姆森科学信息研究所(ISI, http://www.isiknowledge.com)上搜索研究区域内已发表的文献[12, 23, 35]和书籍[14, 36]。为了避免因同义词和物种名称变化等而引起的混淆, 在中国生物物种名录(2022版)数据库中对每个物种的有效拉丁名称进行了校正(http://www.sp2000.org.cn/)。鱼类的入侵风险评估参考Li等[16]和李雪健等[37]的研究结果。
2 结果
2.1 种类组成与优势种
2019—2021年在拉鲁湿地和茶巴朗湿地共采集到10种外来鱼类, 隶属于4目5科, 包括鲫、麦穗鱼、鲤、丁鱥(Tinca tinca)、棒花鱼(Abbottina rivularis)、泥鳅(Misgurnus anguillicaudatus)、大鳞副泥鳅(Paramisgurnus dabryanus)、鲇、小黄䱂和中华青鳉(Oryzias sinensis)。拉鲁湿地和茶巴朗湿地各有8种, 其中鲤和丁鱥仅见于拉鲁湿地, 棒花鱼和中华青鳉仅见于茶巴朗湿地。IRI指数结果显示,鲫、麦穗鱼、大鳞副泥鳅和鲤为拉鲁湿地外来鱼类的优势种, 而鲫、麦穗鱼和小黄䱂为茶巴朗湿地的优势种(表1)。

表1 2019—2021年西藏拉鲁湿地和茶巴朗湿地外来鱼类渔获物概况Tab.1 Non-native fish in Lhalu Wetland and Chabalang Wetland from 2019 to 2021
2.2 渔获物分析
共收集到外来鱼类2714尾、重7230.31 g, 分别占总渔获物的94.86%和70.71%。渔获物物种组成主要是鲫和麦穗鱼, 其个体数分别占外来鱼类总渔获物的38.39%和35.48%, 重量分别占63.95%和7.95%(图2)。依据类别统计, 水产贸易物种的个体数和重量百分比占外来鱼类渔获物的47.75%和90.85%, 非水产贸易物种占52.25%和9.15%, 即外来鱼类群落结构呈引入的水产贸易经济品种与野杂鱼均有的现状; 大型外来鱼类的个体数和重量百分比占外来鱼类渔获物的1.07%和13.71%, 小型外来鱼类占98.93%和86.29%, 即外来鱼类群落结构以小体型种类为主要组成(图3)。

图2 2019—2021年西藏拉鲁湿地和茶巴朗湿地外来鱼类数量和重量百分比(物种代码见表1)Fig.2 Percentage of non-native fish number and weight in Lhalu Wetland and Chabalang Wetland from 2019 to 2021 (species codes are shown in Tab.1)
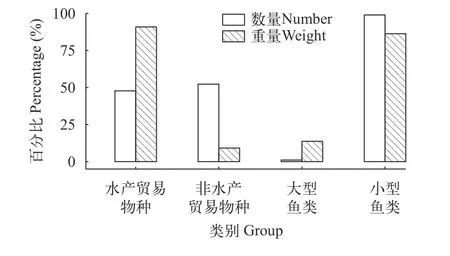
图3 2019—2021年西藏拉鲁湿地和茶巴朗湿地外来鱼类各类别数量和重量百分比Fig.3 Percentages of the number and weight of non-native fish groups in Lhalu Wetland and Chabalang Wetland from 2019 to 2021
2.3 物种生态属性
依据外来鱼类的生态属性分析, 杂食性、底栖动物食性和浮游生物食性的种类均为3种, 肉食性1种; 栖息底层的种类7种, 中层1种, 上层2种; 游动型的种类4种, 静止型6种; 营养级超过3.0的种类达8种; 繁殖力中等的种类6种(表2)。外来鱼类群落功能丰富度指数0.355、功能均匀度指数0.924、功能分散度0.417均高于土著鱼类的相应指数0.003、0.500和0.259, 即外来鱼类群落可占据更大的生态空间、功能性状更复杂, 物种的功能和性状在空间上差异性更大(图4)。

图4 2019—2021年西藏拉鲁和茶巴朗湿地外来鱼类和土著鱼类功能多样性指数Fig.4 Functional diversity of non-native and native species in Tibet wetlands from 2019 to 2021
2.4 种类变化
结合已发表的数据, 西藏拉鲁湿地和茶巴朗湿地共记录外来鱼类12种, 其中6种为高度入侵风险物种(表3)。鲫在历次调查均有发现, 麦穗鱼和小黄䱂在多次调查中被收集到。拉鲁湿地外来鱼类种类数从2001—2003年最初的2种增加到现在的8种。2001—2003年在拉鲁湿地仅发现外来鱼类黄鳝(Monopterus albus)和鲫, 2010—2011年则已增加到7种, 本研究首次收集到丁鱥, 但未采集到草鱼(Ctenopharyngodon idella)和黄鳝; 茶巴朗湿地为5—8种, 2009年调查收集到鲤、草鱼等8种,2013年、2017年及本研究收集到大鳞副泥鳅, 且本研究首次收集到中华青鳉, 但未收集到鲤和草鱼。
3 讨论
3.1 西藏外来鱼类的变动趋势
拉鲁湿地和茶巴朗湿地外来鱼类物种数从最初的2种增加到了现在的10种(表3), 且在西藏自然水体中新发现了丁鱥, 尽管未收集到的黄鳝和草鱼,但这两种是西藏自治区水产品贸易中的常见物种,依然存在进入自然水体的风险; 同时, 麦穗鱼、小黄䱂和棒花鱼等野杂鱼在渔获物中逐渐占据优势地位。研究结果表明, 外来鱼类物种组成呈经济水产养殖或贸易品种和野杂鱼均有的现状, 且小型鱼类占优势地位(表1)。

表3 西藏拉鲁湿地和茶巴朗湿地外来鱼类组成变化和入侵风险Tab.3 Variability of non-native fish composition and basic risk assessment from Lhalu Wetland and Chabalang Wetland in Tibet Autonomous Region
拉鲁湿地和茶巴朗湿地均附属拉萨河流域, 沟渠相接形成流通水系, 其外来鱼类组成在一定程度上反映拉萨河流域外来物种的种类及生存现状。20 世纪有关西藏自治区拉萨河流域渔业资源调查结果中未记录外来种[9]; 21世纪初陆续出现外来鱼类在拉萨河流域中被发现的报道[10,12]; 近年来, 拉萨河流域的外来鱼类的研究已受到众多学者关注[15,23,35,36,38,39]。本研究于2019—2021年开展拉鲁湿地和茶巴朗湿地外来鱼类调查, 除新发现的丁鱥外, 其他外来鱼类已在拉萨河流域乃至雅鲁藏布江流域中已有报道[10,11,40]。
拉鲁湿地和茶巴朗湿地是高原外来鱼类的缩影, 西藏外来物种问题已非常严峻, 复杂多样的外来鱼类组成, 必然给西藏自治区的水生态系统管理和外来种防治等工作带来了新困难, 亦给研究学者提出了新挑战。进一步全面调查及实行长期监测西藏湿地外来鱼类的计划能及时掌握物种组成、种群动态和群落变动趋势等基础资料, 为高原湿地生态系统的生物多样性保护提供科学依据。
3.2 西藏外来鱼类的引种途径及影响
在全球水生生态系统中, 鱼类是最常见的被引入种类[41]。随着西藏地区经济和社会的发展, 水产品市场和水族观赏市场出现了复杂多样的外来鱼类, 如有意引种的鲫、鲤及丁鱥等; 另外还有无意引进的麦穗鱼、棒花鱼、小黄䱂和中华青鳉等。盲目引种、无意引种及引种后疏于管理或弃养等导致引种后的失控或逃逸, 特别是西藏作为中国主要的佛教地区, 历来有鱼类放生的民俗, 无序人为放生活动是外来鱼类进入西藏自然水体中的主要原因之一[39]。
拉鲁湿地和茶巴朗湿地分布最广、种群数量最多、最稳定的外来鱼类是鲫、麦穗鱼、大鳞副泥鳅及小黄䱂, 已被证实在高原湿地成功定殖并建群[15,36]。从鱼类生态属性及现状分析, 这些鱼类均属于典型的r-对策者[19,42,43], 个体小、寿命短、世代更新快, 具有很高的种群重建能力[44], 最终均可能发展成为西藏湿地的入侵种。例如, 麦穗鱼被评定为全球最具入侵潜力的鱼类之一, 已成功入侵到欧洲以及中东地区的绝大多数国家, 在中国亦入侵至云南和西藏等高原水体中[19,45], 本研究结果显示该鱼已成为湿地鱼类群落中主要组成物种; 大鳞副泥鳅于2009年和2010—2011年分别在茶巴朗湿地和拉鲁湿地被发现[23,35], 现已形成稳定的自然繁殖种群[36], 仅经数年时间已达到了生物入侵过程的定居建群阶段。
拉鲁湿地和茶巴朗湿地生态系统封闭性较高,本地土著物种多样性较低, 且静缓流水体环境、水生植被丰富及泥沼底质等稳定的生态系统为外来鱼类提供了良好的栖息繁衍条件, 因此外来鱼类能迅速适应并成功建群甚至扩散[11]。本研究在拉鲁湿地和茶巴朗湿地收集到的外来鱼类在栖息水层、食性类型等生态习性呈多样化(表2), 群落功能多样性指数均高于土著鱼类(图4), 充分证实外来种可占据湿地生态系统中的空生态位以及具有食物竞争优势, 威胁到青藏高原土著鱼类的生存[13]。例如, 本次在西藏调查新发现的丁鱥, 具食性广、耐低温低氧、生长繁殖速度快、对环境的适应能力较强的特点[46], 在中国海河流域研究结果显示是高风险入侵物种之一[37], 若其成功入侵可能会与土著鱼类竞争生存空间、抢夺食物资源, 从而降低西藏湿地的鱼类多样性。
生物入侵已在世界各国造成了严重的生态、经济影响[11,47,48]。外来鱼类进入自然水域中的生态影响被认为是全球范围内水生动物数量减少的主要原因之一[49,50]。已有文献报道显示外来鱼类入侵已造成拉萨河流域土著鱼类资源的衰退[38]。因此, 加强对放生的科学管理及开展外来鱼类的长期监测, 是防范外来鱼类入侵及高原湿地生物多样性保护的迫切任务。
