青海湖主湖区与湖水淹没区的细菌群落结构差异分析
2022-12-21周欣雅马凯丽虞功亮敖鸿毅
张 炜 周欣雅 马凯丽 李 华 虞功亮 敖鸿毅 霍 达
(1.青海师范大学化学化工学院, 西宁 810000; 2.中国科学院水生生物研究所中国科学院藻类生物学重点实验室, 武汉430072; 3.中国科学院大学, 北京 100049; 4.青海省国家公园科研监测评估中心, 西宁 810000)
青海湖地处青藏高原东北部, 平均海拔约3200 m,位于我国东部季风区、西北部干旱区和西南部高寒区的交汇地带[1], 具有明显的地区性气候特点, 干旱、少雨、多风、太阳辐射强烈和气温日较差大,属于高原半干旱高寒气候区[2]。青海湖是我国最大的内陆湖泊和最大的咸水湖[3], 其形成初期属于淡水湖, 拥有大小入湖河流70余条, 但是由于东边出水口处日月山的抬升, 使湖盆封闭, 致使湖水无法汇出, 最终湖水性质转变为现在的微咸水水体[4]。近几十年来, 青海湖水位不断上升。这些上升的湖水淹没周边的草地及砂石, 形成小型的浅水水体[5]。随着畜牧业在青海湖周边的快速发展, 高营养负荷土壤也随着地表径流冲入这些湖滨淹没水体中并逐步进入青海湖及其子湖, 显著地改变了水体中微生物的群落结构及其组成[6,7]。
围绕着青海湖及周边湖泊的浮游细菌群落结构, 许多学者开展了相应的工作。Zhang等[8]在青藏高原东北部研究了盐度及季节性如何影响自由生活细菌与颗粒附着型细菌的群落装配。研究发现扩散限制决定了淡水湖泊的群落组装而随着盐度增加,异质性选择逐渐成为群落装配的决定性因素。Ren等[9]在青海湖主湖和子湖水体中进行了微生物群落结构的调查, 研究发现溪流和湖泊中的微生物群落在丰度上具有显著差异, 这些差异导致了微生物代谢潜力的显著差别。这些研究暗示生境多样性显著地影响青海湖的微生物群落结构。有关海湖流域内不同类型的子湖及湖水淹没区等水体的研究尚未见报道, 因此对于这些不同类型的水体与青海湖高度相关并具有重要的生态功能亟待进一步研究。
浮游细菌驱动湖泊生态系统中多种元素循环,在分解有机质和驱动能量流动等方面发挥了重要的作用[10]。游离生活型浮游细菌具有极高的多样性, 他们数量庞大、循环迅速及响应环境变化灵敏,是水体元素循环最主要的驱动者[11]。附着型浮游细菌依靠黏附在动物、植物和微生物群体上生存,他们的代谢模式往往与其附着的生物高度关联[12]。大量研究揭示传统基于培养的微生物多样性研究具有很大的局限性[13]。随着高通量测序技术的发展, 全面深度的测序将揭示微生物广泛的多样性。由此, 微生物地理学, 即不同生存类型的微生物, 经过栖居地的环境条件筛选, 形成具有本土特征的微生物多样性格局成为当前研究的热点[14]。
微生物栖居环境是影响微生物群落结构的重要因素。在同一水体中, 不同类型栖居地微生物群落差异的研究可更好地了解水环境微生物如何适应环境变化。近年来, 随着高通量测序技术的发展,微生物多样性的研究不再局限于可培养微生物的调查。基于高通量测序技术的扩增子测序使得对特殊生境中微生物的大范围研究成为可能[15]。受全球气候变化的影响, 青藏高原气候趋于变暖变湿,青海湖亦受影响。2004年起青海湖水位保持抬升、水域面积持续增大。如今青海湖水位上涨, 湖水淹没周边放牧的缓地草原, 这种水位的抬升给青海湖带来了丰富的氮磷营养盐, 提高了湖水的营养水平, 同时也形成了具有局部联通性的破碎化斑块。这为我们提供了研究不同类型水体中微生物群落变化的研究模型。
本研究将青海湖主湖与周边湖滨淹没区水体构成的关联水体作为研究模型, 探究微生物群落结构在不同水体类型中的变化, 探究在不同类型水体中特化的可能具有关键代谢功能的微生物群体, 为进一步了解微生物驱动高原盐碱湖泊的能量代谢过程提供帮助。
1 材料与方法
1.1 采样方法、时间及区域
本次调查于2020年8月在青海湖共设置主湖区样点16个, 不同类型湖滨带淹没区样点11个, 现场采用YSI多功能水质分析仪进行pH、溶解氧(DO)、水温(T)、TDS和盐度的检测。对调查区域主湖区及滨湖淹没区均采用北原式采水器采取表层水, 将部分水用于浮游细菌的过滤, 剩余水样进行水化学指标测定。总磷、溶解性磷、总氮、硝酸盐、氨氮、亚硝酸盐及高锰酸盐指数参照文献[16]进行检测。采样点具体坐标如图1所示。
图1 采样点示意图Fig.1 Diagram of sampling sites
1.2 样品采集、处理及DNA高通量测序
将适量体积的(200—500 mL)的水样利用负压抽滤装置富集至聚碳酸酯膜(0.2 μm Millipore GTTP04700)抽干, 锡纸包裹, 并避光低温(–80℃)保存,以备水体总DNA提取。DNA提取使用经典的苯酚-氯仿-乙醇沉淀法, 具体操作步骤见参考文献[15]。采用细菌16S rRNA基因V3—V4区通用引物对DNA样品进行PCR扩增。50 μL反应体系中包含: 1 μL DNA模板, 3 μL 25 mmol/L MgSO4, 5 μL 25 mmol/L dNTPs, 1 μL每个双端引物(338F: 5′-ACTCCTA CGGGAGGCAGCA-3′; 806R: 5′-GGACTACHV GGGTWTCTAAT-3′)[17], 10 μL 5 × Fast Pfu聚合酶,纯水补足体系。扩增程序如下: 98℃预变性2min,30个扩增循环(98℃变性30s, 50℃退火30s, 72℃延伸60s), 72℃终延伸5min, 4℃保温。PCR产物纯化后检测其纯度, 并使用Qubit 2.0进行DNA浓度的定量。参照说明书构建双端300 bp(PE300)文库, 使用Illumina Miseq平台进行测序。
1.3 统计与分析
二代扩增子高通量下机样品数据使用QIIME 2分析[18]。使用Trimmomatic[19]过滤低质量数据。使用Usearch的Unoise 3[20]算法进行降噪、去除嵌合体得到扩增子序列变异和特征表, 得到的ZOTUs使用SILVA 132数据库[21]进行注释。将得到的OTU Table根据获得的最低丰度样品进行抽平, 利用R语言stat包和dplyr包对数据进行统计分析, 各分类单位的丰度采用平均值±标准差的方式进行表示。数据均值比较方式为Welcoxon秩和检验。采用 R语言的Vegan包进行OTU的α多样性分析、β多样性分析。环境因子数据与物种分布数据采用消除趋势对应分析(Detrended correspondence analysis, DCA)后根据特征根值选择最终分析模型为RDA。使用R语言vegan包的rda()进行模型构建, 并使用单因素方差分析(ANOVA)模型参数进行显著性检验。
2 结果
2.1 青海湖主湖区与淹没区浮游细菌群落相对丰度差异分析
通过16S高通量扩增子测序分析发现, 青海湖主湖区及淹没区在分类门水平上相对丰度最高的为变形菌门(Proteobacteria, 44.8%), 且数据波动较小(标准差5.3%), 其次是拟杆菌门(Bacteroidetes,25.9%±7.8%)、蓝细菌门(Cyanobacteria, 13.6%±5.4%)、放线菌门(Actinobacteria, 7.54%±9%)、柔壁菌门(Tenericutes, 3.32%±2%)。数据表明变形菌门和拟杆菌门的细菌在青海湖主湖及湖滨淹没区中均广泛存在(图2A)。从门级水平上来看, 青海湖主湖和湖滨淹没区的微生物丰度差异很大(图2B)。对于高丰度微生物来说, 主湖与淹没区在高丰度物种的组成差异是显著的。在wilcox 差异检验中除一些低丰度类群, 如酸杆菌门(Acidobacteria)、 热境丝菌门(Caldiserica)、CandidatusDependentiae、出芽单胞菌门(Gemmatimonadetes)和暂定名称马尔古利斯氏菌门(CandidatusMargulisbacteria)以外,其余各门均存在极显著差异(P<0.01)。蓝细菌门在主湖中相对丰度显著高于淹没区且总丰度较高。

图2 青海湖细菌群落结构门级差异Fig.2 Bacterial community structure in Qinghai Lake at Phylum level
从属级水平来看(图3), 水月亮菌属(Aquiluna)、玫瑰色杆菌属(Roseibacterium)及MWH-UniP1类群在淹没区的丰度显著高于青海湖主湖区。与之相对应的是Ambiguous、LD29、Nodosilinea、极地杆菌属(Polaribacter)、Polycyclovorans及未培养细菌的丰度在青海湖主湖相对较高(P<0.05)。这些微生物的在两个区域中的差异暗示了这些微生物在不同环境下表现出各自不同的功能。
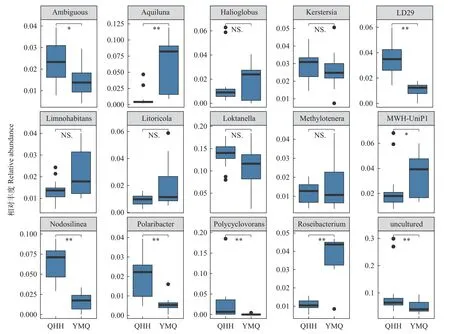
图3 属水平分类下各丰富细菌分布差异(仅相对丰度>1%的分类元纳入统计)Fig.3 Differences in the distribution of abundant bacteria at the genus level (Only taxa with relative abundance >1% are included in the statistics)
2.2 物种多样性指数分析
我们使用Shannon-Winner指数法分析了青海湖及滨湖淹没区各样品的多样性, 结果表明, 淹没区样品微生物多样性显著高于主湖水体中(图4)。两者平均值分别为4.01±0.145和3.67±0.136。Chao1多样性趋势与Shannon多样性相同, 淹没区样品多样性显著高于青海湖主湖。(Wilcoxon test,P<0.01)这表明淹没区水体微生物的种类更为丰富。基于非度量多维尺度法分析样品间的差异表明, 青海湖主湖的微生物群落结构较为集中, 且与淹没区水体可以较为明显的在二维坐标系下被区分; 而淹没区水体的微生物群落结构差异度则相对较大(图5)。个别样品与其他样品在降维结果中相离较远, 这可能是由淹没区水体的复杂环境构成决定的。对于样品的Beta多样性, 我们使用4种不同的方式计算了样品间的群落相似性。我们计算了样品间的Jaccard距离和Bray距离并分别使用Hclust聚类及UPGMA方式进行聚类, 为了便于理解样品间的相似性, 结果以(1- Distance)呈现。四种分析结果均与NMDS的降维分析结果一致。青海湖的大多数样品间群落结构存在极高的相似性(图6)。这表现在Beta多样性距离矩阵聚在一起。仅有QHH07及QHH15样点与淹没区样品存在一定的相似性。对于淹没区样品来说YMQ.05样品则与本研究中所有的其他样品差异度都很大, 其他各样品均遵循相同生境相似的群落结构。

图4 青海湖及滨湖淹没区物种多样性的Alpha多样性指数Fig.4 Alpha diversity index of species diversity in Qinghai Lake and the inundation area

图5 物种群落结构的非度量多维尺度法分析Fig.5 A non-metric multidimensional scaling approach to the analysis of species community structure

图6 样品间Beta多样性分析Fig.6 Beta diversity analysis between samples
从水化学指标来看, 除总氮、总磷、水温和氨氮外, 青海湖主湖与淹没区在多个化学指标均呈现显著差异。其中pH、TDS和硝酸盐为极显著差异(P<0.001)。淹没区水体相较于青海湖主湖呈现出高pH、高可溶磷及高亚硝酸盐的趋势。青海湖主湖则有更高的可溶性有机物、硝酸盐和盐度(图7)。RDA分析(图8)显示影响青海湖主湖与淹没区分布差异的主要环境因子是pH(ANOVA,P<0.001)、总磷(P<0.05)和温度(P<0.05)。这说明, 虽然淹没区与青海湖主湖中差异环境因子众多, 但影响物种丰度分布的环境因子十分有限。
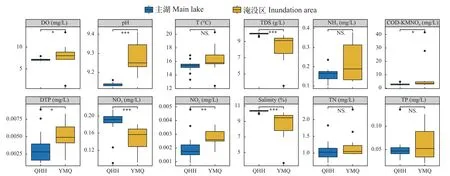
图7 青海湖主湖与淹没区水化学指标差异Fig.7 Differences in water chemistry indicators between the main lake and the inundated area of Qinghai Lake
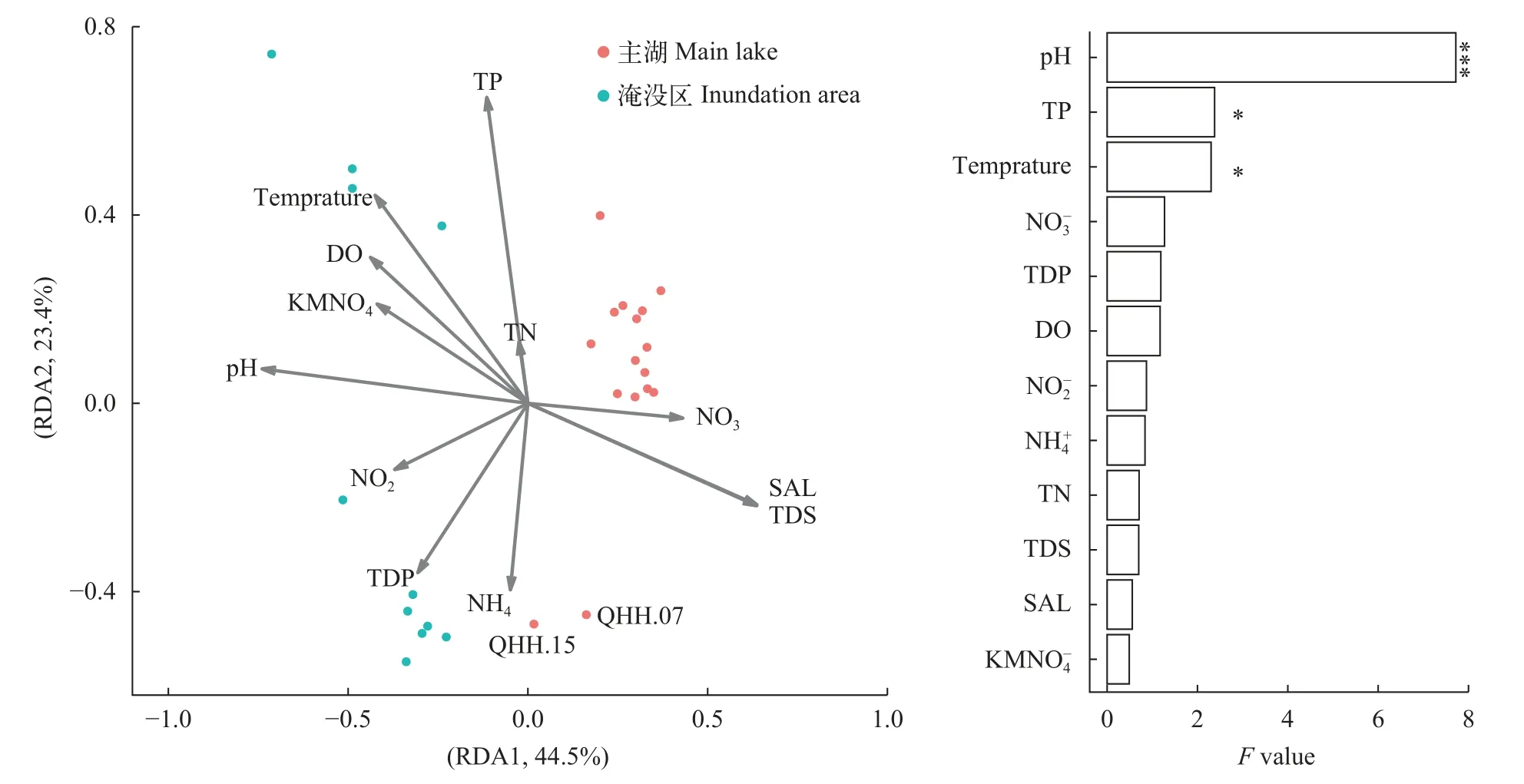
图8 环境因子与物种分布相关的冗余分析Fig.8 Redundancy analysis showing the relationship between environmental factors and species distribution
3 讨论
青海湖水体中变形菌、拟杆菌与蓝细菌是丰度较高的类群, 与之前其他的青海湖水体研究结果一致[22]。同时也与一些淡水水体中的研究相同[23],这表明了这些类群在淡水生态系统中的基础性作用以及其对于盐度的广泛适应性。值得注意的是:在青海湖中节线藻(Nodosilinea)的总体丰度很高,同时在主湖水体的丰度显著高于淹没区。这表明该类群适应高盐度浮游生存环境。节线藻在最近的研究中被报道在淡水[24]和陆地生态系统[25]中广泛分布, 有些研究还发现了部分节线藻可以进行固氮[26]。最近的一项研究在加拿大高原碱水湖泊沉积物中发现了与亚洲湖泊沉积物高度一致的核心微生物类群[27]。本研究进一步证实了节线藻在高原咸水湖泊的碳、氮循环中可能发挥着重要作用。
之前对于青海湖微生物群落特征已有一些报道, Ren等[9]对青海湖流域的湖水和入湖河流的微生物群落进行了密集的实地采样, 结果发现入湖河流和青海湖主湖水体中的微生物多样性存在差异,且群落的Beta多样性结构与功能多样性密切相关。在本研究中, 青海湖主湖中的微生物群落保持较高的一致性, 表现在Beta多样性分析中主湖点位的高度聚集, 同时Alpha多样性指数的平均值波动较小, 这说明青海湖主湖水体混合较为均匀, 区域特征不明显。微生物在水环境中承担着重要的元素循环作用, 复杂的微生物网络构成与环境互作使得群落结构响应环境变化过程十分灵敏[28]。结合水化学指标的特征我们发现, 青海湖淹没区中亚硝酸盐、可溶性磷指标相对较高, 这些元素往往是水体中较为活跃的元素。亚硝酸盐是硝化作用的中间产物, 能够通过氨氧化细菌和亚硝酸盐氧化细菌之间的紧密配合立刻转化为硝酸盐[29]。同样的可溶性磷容易被藻类利用, 最终结合为有机磷等更为稳定的形式被保存在湖泊中[30]。在本研究中, 更高的多样性可能给青海湖淹没区水体带来更多种类的分解者。淹没区的总可溶性有机物在淹没区显著更低说明这些分解者将水体中的有机物分解, 将小的颗粒悬浮物分解成为其他浮游生物可利用的营养物质。淹没区更低的盐度可能给更多的分解者生存提供了有利的空间。
青海湖淹没区在历史上曾经是裸露的草地、土地或是砂石, 水位上升被淹没后, 原本生存在这些植物、矿物表面的微生物经过水体冲刷进入到淹没区中, 这使得淹没区水体中出现了相对较高比例的附着性微生物。对于附着型微生物来说, 其生存依赖于附着基质, 这决定了这些微生物类群在营养物质代谢模式上与浮游类群存在明显的差异。Yang等[22]的研究对青海湖水体的表面沉积物进行了高通量测序。结果发现, 青海湖沉积物表面的微生物群落组成主要受到盐度而非地理距离的影响。我们的研究发现对于淹没区水体微生物与主湖类群显著不同, 且不同附着基质间也存在少量差异。根据我们系统的分析, 淹没区特定属丰度显著高于主湖区, 例如Aquiluna, 由于其不可培养性, 对于其研究仅限于近年来基于测序技术的研究。Leoni等[31]首次报道了Aquiluna在盐度约为14.5%的水体中的存在。我们在青海湖的研究进一步推测Aquiluna在具有固着物的淹没区水体中丰度较高, 暗示了其生活习性可能偏向黏附在基质上, 且具有较强的盐度耐受能力。相比之下, 针对Roseibacterium及MWHUniP1在咸水中的研究已经比较多[32], 这些种类所具有的共同特点是都常被发现在咸水底质或沼泽中, 暗示了对于这一类生境的独特适应性。
4 结论
本研究将青海湖主湖水体与其相关的淹没区水体进行了种群结构及其多样性的比较, 阐明了不同类型水体的微生物群落结构特征。青海湖主湖微生物群落结构在各采样点间差异较小, 证明了青海湖主湖的水体混合较为均匀。与之相对应的是在淹没区水体中, 不同的附着基质造成了各区域的微生物差异较大。淹没区水体中富集的微生物差异类群展现出了明显的营附着生存特征。值得注意的是, 造成两种水体蓝藻丰度最大的属是节线藻属, 我们的研究发现这种丝状蓝细菌在青海湖主湖各个样点均广泛存在, 且主湖丰度显著高于淹没区,这暗示节线藻可以适应咸水环境在驱动咸水水体的元素循环中发挥巨大作用。
致谢:
感谢中国科学院水生生物研究所的王一郎博士、陈友信博士在样品收集及整理中的帮助, 同时感谢天津农学院的曹琪同学在文章修改过程中的工作。
