西藏湿地外来麦穗鱼生活史特征的适应性研究
2022-12-21孙欢欢隋晓云陈毅峰1
孙欢欢 朱 仁 冯 秀 熊 文 隋晓云 陈毅峰1,,3
(1.西藏大学理学院, 拉萨 850000; 2.中国科学院水生生物研究所, 武汉 430072; 3.中国科学院大学, 北京 100049;4.湖北师范大学生命科学学院, 黄石 435002)
生物入侵已经成为全球面临的三大环境问题之一, 在生物多样性[1]、生态系统结构和功能、社会经济发展[2]及人类健康[3]等方面的影响愈来愈严重。其中, 外来鱼类入侵已经对全球范围的生态系统产生了严重负面影响, 是全球鱼类生物多样性丧失的重要因素[4]。外来鱼类入侵后, 可通过捕食、竞争和破坏栖息地等多种方式改变原有生态系统的结构和功能, 从而导致本地物种的减少甚至灭绝[5], 对当地的生态系统和经济发展造成严重不良影响[6]。许多入侵鱼类为适应新环境, 其生活史特征会发生可塑性变化从而更好地适应入侵地环境[7,8]。例如, 欧白鲑(Coregonus albula)入侵到亚北极区上游初期随着种群密度增加, 其生长速度和繁殖力下降[9], 食蚊鱼(Gambusia affinis)入侵到低纬度地区后将更多的能量用于繁殖等[10]。因此, 从入侵鱼类生活史特征方面了解外来鱼类成功入侵的机制, 针对性地采取有效措施进行防治, 从而控制外来鱼类的负面影响[11]。
麦穗鱼(Pseudorasbora parva)隶属于鲤形目(Cypriniformes), 鲤科(Cyprinidae),亚科(Gobioninae), 麦穗鱼属, 原产于我国东部、日本和韩国。由于其食性杂、世代周期短、繁殖力高和生理耐受性强[12], 在不到50年的时间里, 已广泛分布于中亚、欧洲和北非等地[13], 且被欧洲列为最严重的水生生物入侵种之一[14,15]。麦穗鱼入侵到新区域后,通过竞争食物与栖息地、携带新的疾病, 改变栖息地特征等因素对土著鱼类的生存和繁衍及水生生态系统的稳定性构成威胁[16]。如入侵云南高原湖泊后造成当地多种特有鱼类数量减少甚至灭绝, 并产生了一系列连锁的生态影响[17]。
青藏高原通常被称为“第三极”, 是我国最大、世界海拔最高的高原[18], 其特殊的高原水生生态系统中鱼类结构较为简单, 生态环境脆弱, 外来鱼类一旦入侵后, 极易改变整个生态系统的结构和功能,造成巨大的生态负面影响[19]。最早记录麦穗鱼由人类活动而引入到青藏高原的时间为20世纪80年代[20], 目前麦穗鱼已在西藏成功建群且种群规模正在不断扩大[21], 然而有关该鱼在青藏高原生态系统中的生活史特征以及入侵机制等研究报道较少。
因此, 本研究以西藏拉鲁湿地和茶巴朗湿地为研究区域, 通过对麦穗鱼种群年龄、生长和繁殖等生活史特征的研究, 及其与我国的原产地种群生活史特征的比较, 系统探究麦穗鱼入侵到高原后的生活史特征的可塑性变化。在此基础上, 揭示外来鱼类入侵高原生态系统的机制和适应性变化。此外,本研究也能在一定程度上为高原地区外来鱼类入侵的防控和气候变暖背景下鱼类多样性的保护提供科学依据。
1 材料与方法
1.1 研究区域和样本采集
拉鲁湿地是世界海拔最高、面积最大的城市天然湿地[22], 位于西藏首府拉萨市西北角, 为国家级湿地保护区。茶巴朗湿地位于拉萨市曲水县茶巴朗村, 属市级重点湿地生态功能保护区[23]。两个湿地均附属拉萨河流域, 是西藏自治区内典型的高原湿地。为了进一步比较麦穗鱼在青藏高原与原产地生活史特征的差异, 本研究选取南京地区(原产地)的河流开展了麦穗鱼种群的样本采集。南京江宁地处我国东部、长江下游, 属北亚热带湿润气候, 四季分明, 雨水充沛。采样点的具体信息见表1。
2019年7月、2020年8—10月和2021年3—4月使用地笼(长8 m×宽0.25 m×高0.2 m, 网径0.5 cm,放置12h)、抄网(网眼尺寸1.5 mm)和便携式电鱼机(12 V, 2000—3500 W)在拉鲁湿地采集麦穗鱼255尾, 茶巴朗湿地214尾; 2020年12月和2021年4月使用地笼在南京江宁附近自然水域采集麦穗鱼样本616尾。用游标卡尺测量体长和全长(精确至0.01 mm), 用电子天平(HLD-30002型, 中国)称量体重(精确至0.01 g)及性腺重(精确至0.01 g)。运用便携式水质分析仪(YSI Professional Plus, USA)测量采样点的水温并记录整理, 取每尾样本的背鳍起点下方至侧线上方的鳞片5—10枚, 用于年龄鉴定。将鳞片在3%的氢氧化钠溶液中浸泡约8h, 用软刷刷掉表面黏液和杂质。年轮读数在时隔两周分别读取, 通过计算年轮的数量确定鱼的年龄。
同时, 在实地调查的基础上, 本研究在Web of Science(http://webofscience.com)和CNKI(http://www.cnki.net)等数据库中以“入侵麦穗鱼”和“麦穗鱼生活史特征”作为关键词在开展文献的检索和整合分析, 提取中国不同地理种群麦穗鱼生活史特征的数据, 包括最大体长、最大年龄、退算全长、繁殖力和卵径等。
1.2 数据处理与分析
根据采集的样品, 分别处理拉鲁湿地和茶巴朗湿地及南京江宁麦穗鱼种群的相关数据。采用幂函数W=aLb拟合雌雄样本的体长(L)与体重(W)的关系[24]。体长体重方程中指数b值反映鱼类在不同发育阶段和不同生境中的生长发育情况, 釆用Pauly的方法检验关系式中b值与3之间是否存在显著性差异[25]。当b与 3 无显著性差异时, 表明鱼类为匀速生长, 反之为异速生长。使用Image-Pro plus 6.0软件测量麦穗鱼的鳞片。Einar Lea公式Ln=Lrn/R、Rosa Lee公式L=a+bR和非线性BPH(body proportional)公式L=aRb等三种关系方程进行拟合, 选择判定系数(r2)最大者为其最佳回归方程, 并计算退算体长[26]与退算全长。麦穗鱼性腺发育时期的判定主要参照殷名称[27]和严云志等[28]的划分方法。繁殖力采用重量法估算, 取其卵巢重的一部分统计卵粒数, 推算个体绝对怀卵量。绝对繁殖力= (取样的卵粒数/样品重)×卵巢重, 相对繁殖力=绝对繁殖力/去内脏重。在雌鱼卵巢里选取具有沉积卵黄的卵粒进行卵径测量, 每尾随机抽取100粒卵用目测微尺测量其卵径[29]。
运用单因素方差分析(One-way ANOVA)检验拉鲁湿地和茶巴朗湿地麦穗鱼种群与南京江宁种群的体长组成、退算全长和繁殖力之间差异显著性; 运用配对t检验(Pairedt-test)比较拉鲁湿地和茶巴朗湿地麦穗鱼种群与南京江宁种群各年龄组实测体长与退算体长;K-S检验(Kolmogorov-Smirnov test)分析拉鲁湿地、茶巴朗湿地和南京江宁麦穗鱼雌雄群体之间差异显著性。以体长的自然对数值作为协变量进行方差分析, 检验雌、雄个体间体长与体重的关系是否存在差异。检验结果示P<0.05为显著性差异。使用Microsoft excel 2007(Microsoft, USA)、SPSS 13.0(IBM, USA)和Origin 9.0(EA,USA)等软件整理和统计数据并作图。
2 结果
2.1 年龄组成与体长体重分布
拉鲁湿地和茶巴朗湿地及南京江宁鉴定出有效年龄数据分别为250尾、212尾和611尾。结果显示, 拉鲁湿地和茶巴朗湿地麦穗鱼的最大年龄为4龄, 1龄样本均占比最多, 分别为37.20%和48.11%。南京江宁麦穗鱼最大年龄为5龄, 3龄样本量最多,占总数的61.21%(图1)。拉鲁湿地255尾和茶巴朗湿地214尾及南京江宁611尾麦穗鱼的平均体长分别为(40.79±11.71)、(44.12±9.39)和(63.50±9.38)mm,平均体重分别为(1.74±1.64)、(1.72±1.35)和(5.14±2.63)g。拉鲁湿地和茶巴朗湿地麦穗鱼的最大体长分别为85.06和76.00 mm, 最大体重分别为14.68和7.90 g; 南京江宁麦穗鱼最大体长和最大体重为101.00 mm和20.56 g(图2和图3)。拉鲁湿地和茶巴朗湿地麦穗鱼种群体长组成与南京地区种群相比较小, 且存在显著性差异(P<0.05)。拉鲁湿地和茶巴朗湿地麦穗鱼的最大体长与原产地相比也较低。

图3 麦麦穗鱼种群的体重分布 (A.拉鲁湿地和茶巴朗湿地;B.南京江宁)Fig.3 Body weight of P.parva Lhalu wetland and Chabalang wetland(A)and Nanjing Jiangning(B)
K-S检验(Kolmogorov-Smirnov test)结果显示,拉鲁湿地和茶巴朗湿地麦穗鱼雌雄群体体长和体重均不存在显著性差异(P>0.05), 南京江宁存在显著性差异(P<0.05), 由此将拉鲁湿地和茶巴朗湿地雌雄群体合并分析, 对南京江宁雌雄群体进行分别分析。
2.2 体长与体重的关系
麦穗鱼体长和体重的关系在拉鲁湿地为W♂=0.021L2.966(r2=0.982)、W♀=0.014L3.224(r2=0.986);茶巴朗湿地为W♂=0.011L3.282(r2=0.990)、W♀=0.014L3.123(r2=0.989); 南京江宁为W♂=0.014L3.146(r2=0.939)、W♀=0.028L2.763(r2=0.894)。经协方差分析, 拉鲁湿地和茶巴朗湿地麦穗鱼雌鱼和雄鱼间的体长体重关系差异不显著(P>0.05), 故将其合并为种群总体的幂函数方程分别为W♂+♀=0.018L3.085(r2=0.982)和W♂+♀=0.013L3.197(r2=0.988), 南京江宁麦穗鱼雌鱼和雄鱼间的体长体重关系差异显著(P<0.05), 因此将雌雄分开拟合(图4)。t检验结果显示, 体长与体重回归方程的幂指数“b”与3之间均有明显的差异(P<0.01)表明拉鲁湿地和茶巴朗湿地及南京江宁的麦穗鱼种群均为异速生长。
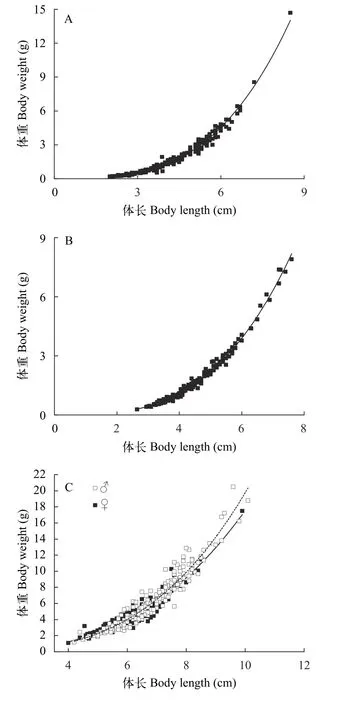
图4 麦穗鱼体长与体重的关系 (A.拉鲁湿地; B.茶巴朗湿地;C.南京江宁)Fig.4 Relationship between body length and body weight of P.parva in Lhalu Wetland (A), Chabalang Wetland (B)and Nanjing Jiangning (C)
2.3 体长的退算
拉鲁湿地和茶巴朗湿地及南京江宁麦穗鱼种群的鳞径与体长的关系式分别为L♀=21.676R+7.766(r2=0.886)和L♂=29.184R0.792(r2=0.886)、L♀=16.946R+15.048(r2=0.857)和L♂=18.290R+13.157(r2=0.858)、L♀=33.710R0.607(r2=0.686)和L♂=16.194R+20.485(r2=0.800)。拉鲁湿地和茶巴朗湿地及南京江宁麦穗鱼种群的实测体长与退算体长均无显著性差异(P>0.05), 茶巴朗湿地种群的各龄组平均退算体长略高于拉鲁湿地(P<0.05), 但均显著低于南京的种群(P<0.01)。根据已有文献报道的麦穗鱼长度退算数据多为退算全长, 本研究对拉鲁湿地和茶巴朗湿地及南京江宁麦穗鱼种群的鳞径与全长的关系式也进行了拟合,方程式分别为L♀=35.446R0.802(r2=0.886)和L♂=35.348R0.799(r2=0.895)、L♀=20.490R+18.380(r2=0.857)和L♂=22.591R+15.879 (r2=0.823)、L♀=43.558R0.564(r2=0.677)和L♂=42.266R0.640(r2=0.774)。茶巴朗湿地麦穗鱼种群的各龄组平均退算全长略高于拉鲁湿地的, 且通过方差分析结果显示两者存在显著性差异(P<0.05)。与已报道的数据比较, 拉鲁湿地和茶巴朗湿地麦穗鱼种群的每龄平均退算全长低于巢湖、洞庭湖、南京江宁等原产地种群(图5)。
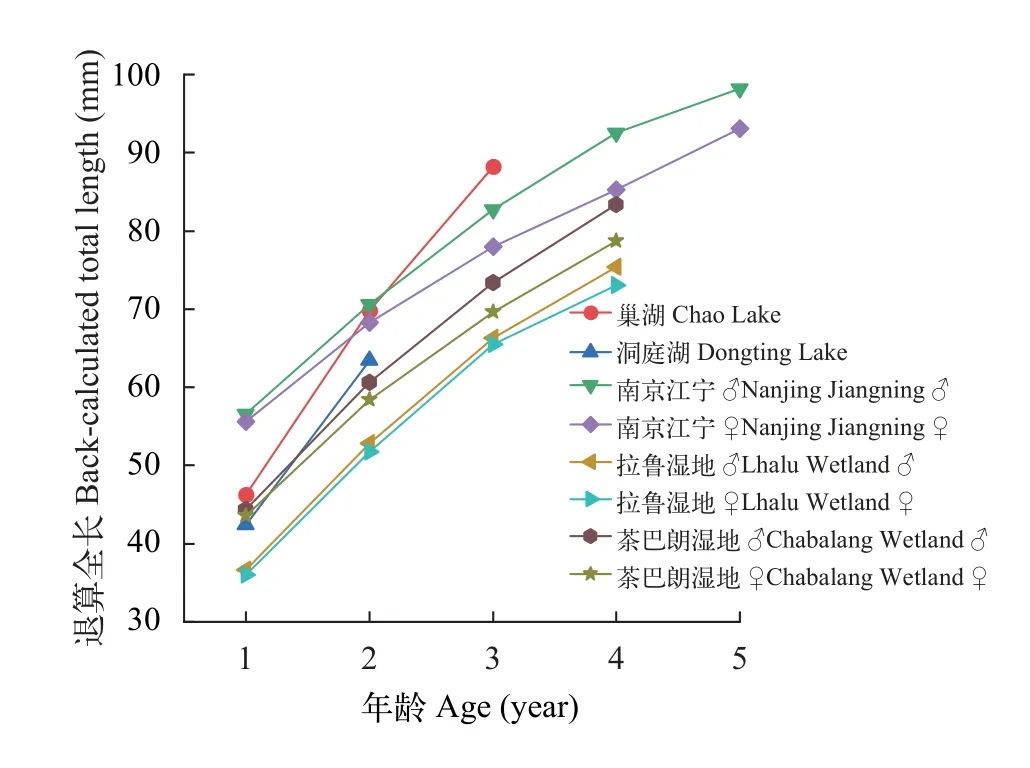
图5 不同地理种群麦穗鱼退算全长的比较Fig.5 Comparison of back-calculated total length of P.parva from different populations
2.4 繁殖力与卵径
拉鲁湿地和茶巴朗湿地及南京地区雌雄性比分别为0.91﹕1、1.08﹕1和1.44﹕1(表2)。茶巴朗湿地在繁殖期仅收集到一尾雌性麦穗鱼, 绝对繁殖力为364粒, 卵径为0.97 mm, 相对怀卵量520粒/g。拉鲁湿地平均绝对繁殖力为(926±476)粒、卵径为0.70—1.00 mm, 平均卵径为(0.92±0.10)mm, 相对繁殖力为218—1545粒/g, 平均相对繁殖力为(575±406)粒/g。南京江宁平均绝对繁殖力为(1220±704)粒, 卵径为0.80—1.40 mm, 平均卵径为(1.00±0.14)mm,相对繁殖力为136—1266粒/g, 平均相对繁殖力为(441±191)粒/g。拉鲁湿地麦穗鱼的平均绝对繁殖力与原产地的相比均较低。拉鲁湿地麦穗鱼的相对繁殖力稍高于南京的(P>0.05), 但低于巢湖、洞庭湖的麦穗鱼种群, 其卵径与太湖(卵径: 1.30—1.40 mm)和四川(卵径: 1.00—1.70 mm)地区麦穗鱼的卵径相比也较小。
3 讨论
3.1 年龄和生长
环境因素影响鱼类的生长速度和最终体长大小[30], 拉鲁湿地和茶巴朗湿地麦穗鱼的平均体长与原产地相比均较小(表2)[28,31—35], 同龄个体的体长相比于原产地也较短, 生长率与原产地相比较低(图5)。这与Jia等[21]和Ding等[36]的研究结果较一致, 麦穗鱼入侵到青藏高原以后, 由于高海拔和低温, 导致平均水温低, 生长季节短, 生长较慢[32,37]。其次, 食物的丰度和质量决定了生物对资源的吸收,以及资源分配到个体的生长中, 与低海拔地区相比,青藏高原的食物丰富度和对资源的利用率要低得多[21]。这可能也是麦穗鱼在高原地区生长较慢, 体型较小的重要原因之一[38]。此外, 种群密度[39]、生态位重叠[40]也对麦穗鱼的生长有所影响。麦穗鱼的年龄鉴定结果主要以低龄个体为主, 这也与天津地区[32]和白洋淀地区[33]所调查的结果相类似。麦穗鱼的最大年龄在不同的地方有所不同可能受到多种因素的影响, 如生态系统生产力[41]、性别差异和环境稳定性[42]等。

表2 不同地区麦穗鱼生活史特征的比较Tab.2 Comparison of life-traits of P.parva in different areas
3.2 繁殖力与卵径
在麦穗鱼入侵到拉鲁湿地后, 其繁殖力与原产地相比均较低, 卵径较小。此外, Grabowska和Przybylski[43]发现入侵中欧的淡水鱼类也表现出体长中小、繁殖力相对较低的生活史特征。可见繁殖力变动可能是促进其成功入侵的重要机制。其中, 温度是调节鱼类生长、发育和繁殖性能的关键非环境因子[44,45]。四川东部地区麦穗鱼在20—28℃时产卵[34]; 黑龙江地区麦穗鱼在水温达到15—19℃时也可开始产卵, 然而在本次调查中, 拉鲁湿地和茶巴朗湿地四月平均水温分别为11和12℃, 与以上研究样点开始繁殖温度相比更低, 且Gozlan等[13]认为水温是麦穗鱼生活史特征表达的主要决定因素, 因此较低的温度可能是茶巴朗湿地和拉鲁湿地麦穗鱼的繁殖力低于其他地区种群的繁殖力的原因之一。此外, 拉鲁湿地麦穗鱼的平均卵径相比于原产地也较小, 对于高原上的麦穗鱼来说, 由于资源短缺和低温, 麦穗鱼能够投入到繁殖的总能量是有限的。卵径和繁殖力之间的权衡通常与母体适合度和仔稚鱼存活率密切相关[46,47]。Smalas等[48]认为通过调节后代的大小来使母体最大化的适应当地环境很可能是高繁殖生物体的普遍现象。在总繁殖能量有限的情况下, 生产更小的卵可产生相对更多的后代, 从而提高麦穗鱼在高原环境中的生存适应能力[49,50]。
3.3 高原入侵鱼类管理
麦穗鱼为适应严苛的高原环境在入侵到拉鲁湿地后表现出了生长缓慢、繁殖力低和体型较小的生活史特征并与其较强的环境耐受性共同促进了其在当地的传播。丁慧萍[51]对茶巴朗湿地麦穗鱼的食性研究发现, 麦穗鱼和拉萨裸裂尻和异齿裂腹鱼食性重叠程度较大, 也会和土著鱼类竞争栖息地。麦穗鱼的入侵可能会对本土鱼类和水生生态系统构成新的潜在威胁。因此, 我们今后应当加强有关土著鱼类保护以及麦穗鱼入侵机制和防控等方面的研究。保护好土著鱼类, 确保自然生态系统的完整性和良好的调节能力, 是西藏发展的重要内容和可持续发展的战略选择[52]。此外, 对于麦穗鱼未入侵的地区, 应当制定相关政策法规禁止其被人为引入, 以防止麦穗鱼的进一步扩散, 使更多的水域受到影响。同时, 相关部门也应加强宣传和教育,提高公众对麦穗鱼和其他非本地鱼类产生危害的认识[53]。
