豆类蛋白组装机制及其与蛋白加工特性关联性的研究进展
2022-12-15刘业学李雪莹刘佳萌魏立坤路福平王稳航
李 玉, 刘业学, 李雪莹, 刘佳萌, 魏立坤, 路福平, 王稳航
(1.工业发酵微生物教育部重点实验室/天津科技大学 生物工程学院, 天津 300457;2.北京市产品质量监督检验研究院, 北京 101300; 3.天津科技大学 食品科学与工程学院, 天津 300457)
豆类是蛋白质的重要来源,球蛋白(globulin)和白蛋白(albumin)是豆类蛋白中的两种主要成分。球蛋白是盐溶性蛋白质,约占豆类蛋白总质量的70%,其又可以细分为11S球蛋白(legumin)和7S球蛋白(vicilin和covicilin);而白蛋白是水溶性的,占豆类蛋白质量的10%~20%[1]。豆类蛋白最重要的作用是其能够赋予食物功能特性,如:乳化性、发泡性、凝胶性和成膜性,因而在食品加工过程中备受关注。此外,根据豆类蛋白的氨基酸图谱可知其具有含量较高的人类必需氨基酸,如:赖氨酸、亮氨酸、天冬氨酸和精氨酸,因而豆类蛋白具有较高的营养价值[2]。豆类中的蛋白质和多肽在机体健康方面也发挥着重要作用[3]。如水解扁豆、豌豆、大豆和鹰嘴豆蛋白产生的多肽具有显著的抑制血管紧张素-1转换酶(ACE)的特性[4];并且还具有结合胆汁的能力,有利于去除胆汁酸并增加胆固醇的代谢,降低血液中的胆固醇水平[5]。
蛋白质分子在一定条件下会发生聚集组装,这是一种蛋白质结构单元之间或者蛋白质与其他高分子(酚类和多糖)之间以非共价键(氢键、疏水相互作用以及静电相互作用)或者共价键为驱动力,进行相互聚集组装的行为。组装行为使蛋白质物化特性发生改变,这将为蛋白质改进现有结构或创造出新的结构奠定基础,从而获得其他方式无法提供的具有新颖结构(组装结构)的功能属性,为各种食品应用制备不同的材料。另外,蛋白质与活性化合物的组装行为可有效降低这些化合物对化学、生物或物理降解的不稳定性,有助于其保持最佳的活性特性。因此,了解豆类蛋白在不同环境下的组装行为对豆类蛋白在食品加工中的进一步开发利用具有重要意义。
1 豆类蛋白分子的组装行为及影响机制
蛋白质的结构会在环境因素,如pH值、温度和离子类型及强度的影响下发生改变,进而驱动蛋白质进行组装;另外,交联酶会诱导蛋白质分子间进行交联而成为蛋白质组装的强效驱动力;蛋白质与多糖和酚类化合物之间的相互作用会促使蛋白质发生组装行为。
1.1 pH值对豆类蛋白组装的影响机制
溶液的pH值会决定蛋白质表面电荷的类型及分布,影响蛋白质分子内折叠和蛋白质分子间的相互作用,影响蛋白质构象的稳定性和胶体的稳定性,从而影响蛋白质的聚集组装行为[6-7]。根据pH值的不同,蛋白质会组装成两种不同的形态:当pH值远离蛋白质等电点时,就会形成半柔性蛋白纤维聚集体;而当pH值接近蛋白质的等电点时,就会形成球状的聚集体[8]。
蛋白质的半柔性纤维丝始于蛋白质结构的部分展开,半柔性蛋白纤维丝形成的公认模型是成核- 伸长型(图1)[9-11]。天然状态的球蛋白具有紧密的刚性结构,因此其纤维化过程需要先通过环境条件的剧烈变化将其刚性结构转变为松弛的部分展开构象。一般认为,在正常的环境条件下,溶液中蛋白质的天然球状构象与部分展开构象处于动态平衡状态。当处于极端pH值条件下,蛋白质表面带有致密的电荷,显著增加分子内和分子间的排斥相互作用,从而导致动态平衡向部分与完全展开状态之间转移。这种部分展开的构象是球蛋白纤维化的重要先决条件,因为完全展开的球蛋白缺乏有序结构,而部分展开的构象则产生了特定的分子间相互作用,例如氢键、静电力和疏水相互作用,可用于驱动低聚物和原纤维丝的形成[10]。在低pH值条件下可促使豆类蛋白的球状结构部分展开并发生水解作用[8-9]。通过水解获得的蛋白质短肽或单体自组装成核,然后在蛋白核的基础上添加蛋白质单体成长为原纤维丝,而蛋白质的聚集速率与温度和水解速率有关,并随着温度和水解速率的升高而升高[11]。Loveday等[12]研究了pH值为1.6~2.4对蛋白纤维形成的影响,结果发现这几种pH值条件下均可形成蛋白纤维,但是在pH值为1.6的条件下形成的速率最快,这可能是由于在更低的pH值条件下蛋白质能够更快水解。在pH值为2的条件下,大豆球蛋白和大豆分离蛋白均可获得半柔性蛋白纤维聚集体[13]。
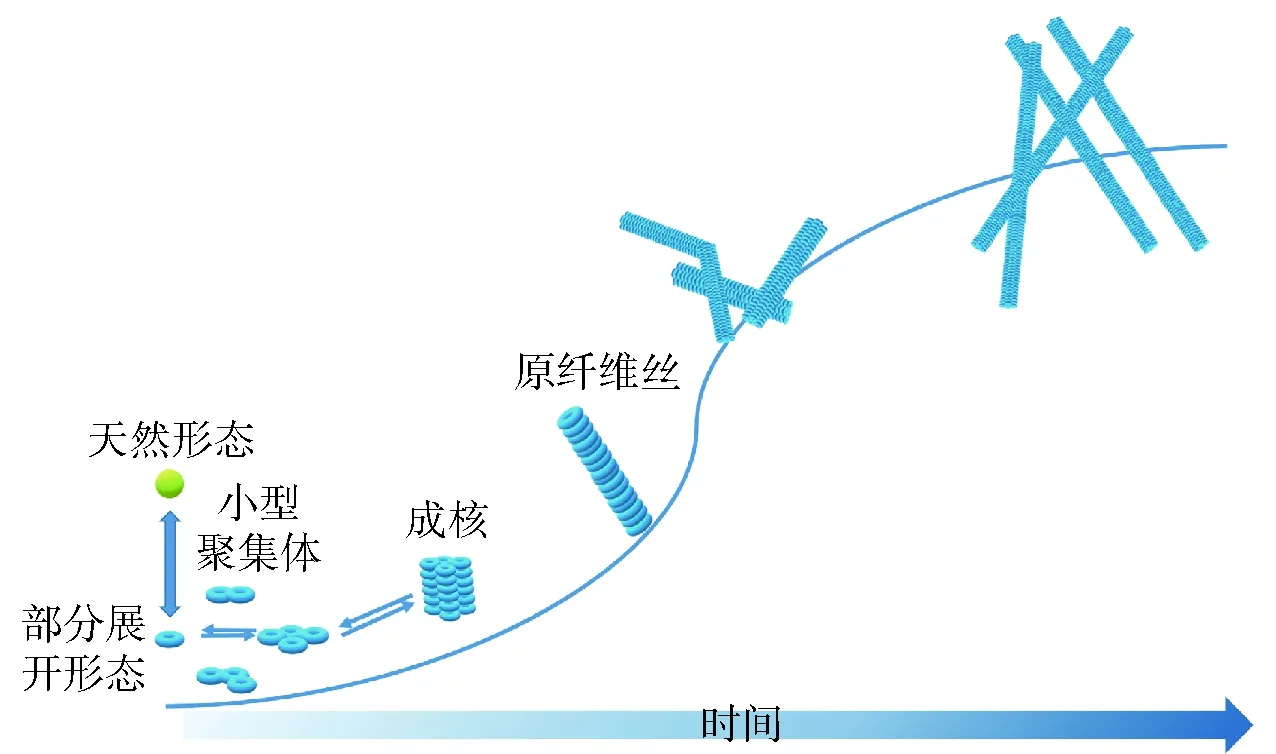
图1 蛋白纤维形成机制示意图
1.2 温度对豆类蛋白组装的影响机制
温度通过影响蛋白质扩散速率,蛋白质间相互作用,蛋白质/聚集体的溶解度,构象稳定性以及化学降解等性质而影响蛋白质的组装行为[7]。当温度升高到变性温度以上时,球蛋白就会部分展开,从而暴露先前埋藏在结构中的一些基团;同时,升温还会增加蛋白质扩散和碰撞的速率而加速蛋白质聚集过程,最终可能导致不同分子之间通过疏水相互作用、氢键和二硫键等作用力使蛋白质聚集[14]。
Guo等[15]研究了大豆蛋白中的7S球蛋白和11S球蛋白在100 ℃和pH值为7的条件下处理30 min后的聚集行为。结果发现在温度超过60 ℃时,β-伴大豆球蛋白会形成可溶性的蛋白聚集体,而当温度升高至100 ℃时,则会形成不可溶的蛋白聚集体。将大豆蛋白分散溶液在pH值为5.9和95 ℃条件下热处理30 min,可获得纳米凝胶颗粒。这些纳米颗粒的特征是具有一个疏水核心(由7S的β-亚基和11S的碱性亚基组成)和一个亲水外壳(由7S的α和α′亚基以及11S的酸性多肽组成),其内部结构主要由二硫键和疏水相互作用稳定[16]。豌豆分离蛋白在加热85 ℃,60 min后,其11S球蛋白先解离,然后通过疏水相互作用及二硫键重新组合,最终形成高分子量的无规则结构的蛋白聚合物;与11S球蛋白相比,7S球蛋白由于其结构中缺乏半胱氨酸残基,而具有较低的热稳定性,因此仅通过非共价相互作用形成聚集体[17]。在相同的pH值条件下,不同的加热时间则会影响蛋白纤维的长度,较长时间的加热水解可将蛋白质所形成的短的蛋白纤维转变为长链的蛋白纤维。芸豆7S球蛋白溶液经加热15~60 min后可以观察到短的蛋白纤维链,而当加热时间超过10 h后,便可获得长链的蛋白纤维[18]。豌豆蛋白在85 ℃下处理20 h后则会形成微米级长度的纤维聚集体;另外,由此得到的豌豆蛋白纤维具有与大豆蛋白纤维相似的结构特征且比使用乳清蛋白获得的纤维更具柔韧性、分支性和卷曲性[19]。
1.3 离子类型及强度对豆类蛋白组装的影响机制
1.3.1离子类型的影响机制
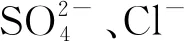
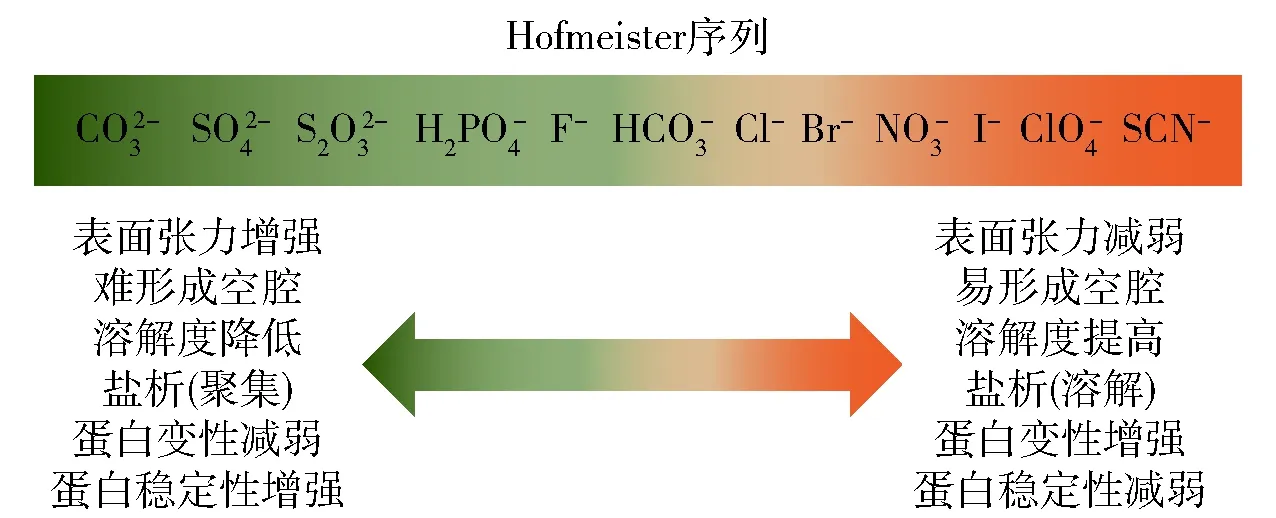
图2 Hofmeister序列
1.3.2离子强度的影响机制
溶液中的离子强度是影响蛋白聚集组装的另一个重要条件。通过改变溶液中的离子强度,可以调节蛋白质纤维的韧性及长度。在pH值为2时,蛋白质组装形成纤维聚集体,在低离子强度下蛋白质纤维呈现出刚性特性;提高离子强度后,蛋白质纤维则变得柔软;在pH值7时,低离子强度下会形成小的细长聚集体,而在高离子强度下,小聚集体则会更倾向于随机结合形成较大的蛋白质组装体[23]。利用离子可以促使蛋白质聚集这一特性,Li等[24]发现,在大豆蛋白聚集物/右旋糖酐混合溶液中加入盐可以屏蔽胶体上的电荷,减少两者间的静电相互作用,从而实现大豆蛋白聚集组装。
1.4 交联酶对豆类蛋白组装的影响机制
蛋白质除了受外部环境影响而发生聚集组装外,交联剂的使用也是驱动蛋白质进行组装的一种重要手段。在交联剂的作用下,蛋白质分子内部或者蛋白质分子之间通过共价键或者非共价键作用促使蛋白质交联聚集,是改善重组蛋白的理化性质、提升蛋白质功能特性的重要手段之一。近年来,酶法作为一种温和的交联方法越来越引起人们的关注。可用于食品蛋白交联的酶有:谷氨酰胺转氨酶、酪氨酸酶、漆酶以及赖氨酰氧化酶等[25]。
1.4.1谷氨酰胺转氨酶的影响机制
微生物来源的谷氨酰胺转氨酶(microbial transglutaminase, MTG, EC 2.3.2.13)是现阶段研究最深入、应用最广泛的驱动蛋白质组装的一种酶。MTG诱导的交联反应在不改变pH值、风味、颜色以及营养价值的情况下,驱动蛋白质进行聚集组装,进而改善蛋白质的功能特性和温度稳定性[26]。在MTG诱导蛋白质聚集组装中,蛋白质的聚集程度与蛋白质的浓度有关。例如,研究发现当大豆分离蛋白质质量分数小于3%时,MTG诱导的交联反应会导致蛋白质组装,形成小的蛋白聚集体;而当质量分数高于3%时,MTG则会诱导小的蛋白聚集体发生进一步的组装,进而使蛋白质发生凝胶化[27]。蛋白质的种类也会影响MTG驱动蛋白质组装行为的发生。Djoullah等[28]在利用MTG驱动豌豆蛋白的白蛋白组分和球蛋白组分在化学变性(DTT)和物理变性(热处理)下的聚集组装的研究中发现,在质量分数为10%的蛋白质浓度下,豌豆球蛋白经MTG交联后会顺利完成聚集组装,进而形成坚固而有弹性的化学凝胶,而白蛋白组分则无法完成组装行为。在大豆分离蛋白的交联中也发现了类似的现象,相对于11S球蛋白亚基,7S球蛋白的亚基是MTG作用的更好底物[29]。Gan等[30]深入分析发现,11S球蛋白亚基对MTG的敏感性较低,是因为它们所含的谷氨酰胺和赖氨酸(酶的主要底物)水平要比酸性多肽低。这些结果表明,酶促交联反应驱动的蛋白质聚集行为与蛋白质的氨基酸组成及构象高度相关,在MTG诱导豆类蛋白的聚集及组装应用中,必须合理选择蛋白质的来源及种类,才能使交联酶在蛋白质的组装中发挥更有效的作用。
1.4.2酪氨酸酶的影响机制
与谷氨酰胺转氨酶类似,氧化酶类中的酪氨酸酶(tyrosinase, EC 1.14.18.1),也称为酚氧化酶,可以在没有辅助因子的情况下催化大分子网络的形成。在大多数动植物中,酪氨酸酶具有相当广泛的底物特异性。Glusac等[31]指出,豌豆蛋白是酪氨酸酶的一种良好的底物,在酶处理30 min内即可驱动豌豆蛋白形成组装聚合物。Chen等[32]则比较了酪氨酸酶和MTG交联后形成的明胶和壳聚糖凝胶,发现酪氨酸酶更能诱导凝胶的形成。Isaschar-Ovdat等[33]研究表明,酪氨酸酶交联大豆11S球蛋白后可增加凝胶硬度并形成更有序的网络结构但弹性有所降低。
1.4.3漆酶的影响机制
漆酶(EC 1.10.3.2)可以利用氧分子作为末端氧化剂,催化酚类化合物发生单电子氧化,从而产生水和自由基,自由基底物可进一步进行非酶反应,包括歧化、聚合、水合和断裂。值得注意的是,漆酶催化反应的唯一副产物是水,因此漆酶在豆类蛋白食品加工行业具有巨大的潜力[34]。研究结果表明,大豆分离蛋白经漆酶交联组装后可以显著改善其凝胶强度,储存模量以及持水性,并且还会使大豆分离蛋白凝胶具有更加均匀的网络结构[35]。
1.4.4赖氨酰氧化酶的影响机制
赖氨酰氧化酶(EC 1.4.3.13)也许是蛋白质交联中最具有应用前景的氧化还原酶之一,它是铜依赖性的胺氧化酶,负责胶原蛋白和弹性蛋白的氧化交联,同时将O2还原为H2O2。在反应中,赖氨酸侧链被氧化为相应的醛,并且通过两个不同的途径产生交联反应:在第一种情况下,两分子醛化合物缩合形成醛醇缩合产物;在第二种情况下,来自第二个赖氨酸残基的胺与醛反应形成席夫碱,从而完成交联反应[36]。Bakota等[37]发现赖氨酰氧化酶通过赖氨酸残基的氧化交联,进而驱动多域肽组装成纳米纤维结构。虽然赖氨酰氧化酶现在还没有应用在豆类蛋白的组装中,但基于其可以促进多肽组装成纳米纤维这一优异性质,将会在豆类蛋白的组装和实际应用中发挥越来越重要的作用。
1.5 多糖和酚类对豆类蛋白组装的影响机制
1.5.1多糖的影响机制
蛋白质- 多糖相互作用在生物系统(维持细胞完整性和细胞分裂)、控制食品的宏观特性(流动、稳定性、质地和口感)以及可食用包装的生产中发挥着重要的作用。研究表明,多糖与豆类蛋白之间有两种相互作用:物理相互作用和美拉德反应(图3)[38]。物理相互作用指的是蛋白质与多糖之间的非共价作用,包括静电作用、氢键作用和疏水作用,其中静电作用是主要驱动力。与美拉德反应相比,蛋白质- 多糖间的物理相互作用具有方便、易操作等特点,因而在调控蛋白质组装方面具有更加便捷的优势。
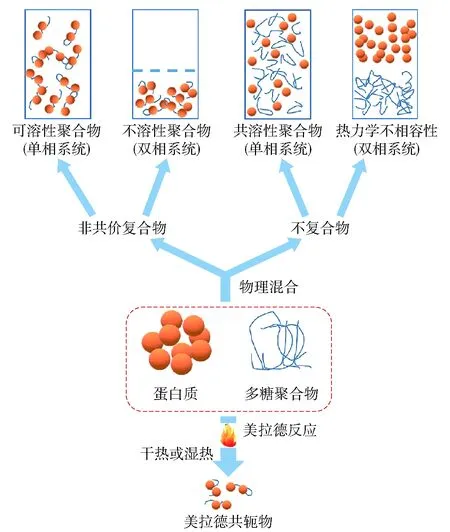
图3 豆类蛋白和多糖相互作用示意
溶液的pH值会影响生物聚合物在溶液中的官能团的电离,进而改变两者间的静电作用,因此在蛋白质- 多糖的聚集组装中发挥着重要的作用。如果将蛋白质- 多糖混合物溶液的pH值从中性调整为酸性,可以通过使用3个临界 pH 值(pHc、pHφ1、pHφ2)来划分4个不同的pH区域,根据它们的相互作用,对应于多糖与豆类蛋白混合物的4种不同相行为(pH>pHc,共溶;pHc~pHφ1,可溶性络合物;pHφ1~pHφ2,络合物凝聚层;pH 1.5.2酚类的影响机制 酚类化合物广泛存在于天然植物性食品原料中,具有较强的抗氧化性能,因此在食品领域有着广泛的应用。酚类化合物与蛋白质之间的共轭组装行为会影响蛋白质的功能特性,如乳化性能、凝胶性能、溶解度、热稳定性和发泡性能。酚类和豆类蛋白间的结合机制(见图4)[41]可以分为单位点结合[图4(a)]和多位点结合[图4(b)]两种结合机制。单位点结合是指酚类化合物仅与一个蛋白质位点相互作用,当酚类化合物与蛋白质的摩尔比例较高时,两者则会通过该方式进行结合;多位点结合仅适用于足够大小的酚类化合物,能够与蛋白质上一个以上的位点相互作用,从而促使蛋白质之间形成交联[41]。另外,根据作用方式的不同又可以分为非共价相互作用(疏水键、离子键和氢键)和共价相互作用(碱性反应、自由基接枝、多酚氧化酶、漆酶、酪氨酸酶)[42]。影响蛋白质- 多酚相互作用的因素有很多,例如温度、pH值、蛋白质类型、蛋白质浓度、酚化合物的类型和结构、盐浓度以及化学试剂(Na2SO3)[43]。这些因素均会引起蛋白质结构发生变化,使其暴露出更多的结合位点,进而通过疏水作用、氢键、静电作用以及共价键与酚类化合物相互结合产生组装行为。 图4 酚类化合物与豆类蛋白的结合机制 绿原酸和槲皮素会使大豆蛋白的等电点向低pH区域转移,这可能是由于酚类化合物与蛋白质的反应可引起蛋白质的交联,降低了蛋白质分子的净电荷,从而诱导蛋白质进行聚集组装;另外,蛋白质的二级和三级结构也会发生变化,从而影响分子的表面性质,使其具有亲水性,进而对蛋白质的溶解度产生影响[44]。Ali等[45]发现,在pH值7和pH值9条件下,大豆球蛋白和没食子酸相互作用后其热变性温度分别从(92.5±0.1)℃和(92.4±0.3)℃提升至(95.6±0.1)℃和(95.1±0.2)℃,这表明酚类化合物与蛋白质通过氢键和疏水相互作用发生聚集组装行为,形成蛋白质- 多酚复合物,从而提升了蛋白质的热稳定性。Yan等[46]利用EGCG(表没食子儿茶素没食子酸酯)接枝大豆分离蛋白,结果显示大豆分离蛋白的β-转角和β-折叠结构含量减少,而α-螺旋和无规则卷曲结构的含量增加,表明EGCG可以使大豆分离蛋白暴露更多的蛋白内部疏水基团,进而改变蛋白的二级结构,这为大豆分离蛋白聚集组装提供了重要的前体条件。 蛋白质的乳化性、发泡性、凝胶性和成膜性等功能特性是在储存、加工、制备和食用时引起蛋白质行为变化的物理和化学特性。蛋白质的组装行为对改善豆类蛋白的品质和加工特性也起着重要作用。合理控制蛋白质的组装,可以显著提高蛋白质的加工特性,改善和保持食品特殊品质。 乳化特性的提升会促进脂肪乳液的形成和稳定,使蛋白质广泛应用于香肠、意大利面、汤、蛋糕等食品中。蛋白质的组装可有效改善其乳化特性,如Wan等[47]研究发现,甜菊糖苷- 大豆分离蛋白二者的组装聚集体在油水界面上相互吸附,形成大豆分离蛋白和甜菊糖苷混合界面层,减小乳滴粒径从而显著提高了乳液的物理稳定性。大豆分离蛋白与茶多酚的偶联组装也会显著提高大豆分离蛋白的乳化能力,最高可达43%,乳化稳定性最高可达59%;并且当使用0.75%绿茶多酚时,大豆分离蛋白- 茶多酚偶联物表现出最佳的乳化性能[48]。 在烘焙食品、糖果和甜点等食品加工应用中,蛋白质则主要起泡沫形成和稳定泡沫的作用。蛋白质的发泡特性改善可以形成更稳定的薄膜来截留气体。通过对蛋白质的结构及聚集组装状态进行控制,可以实现蛋白质发泡性的有效调控。多糖类物质,如黄原胶,会驱动蛋白质在界面处进行组装并形成聚集体,降低排水率和减少泡沫的破裂来增加大豆蛋白的发泡特性;同样,将黄原胶添加到羽扇豆、豌豆和蚕豆的分离蛋白中也会增加起泡能力,这可能要归因于界面处蛋白质吸附的改善和水相黏度的增加,这使得水流失变得困难[49]。Sui等[50]发现,多酚化合物会通过非共价作用和共价作用驱动蛋白质进行组装,实现了大豆分离蛋白的发泡性能的提升;并且共价作用下的大豆分离蛋白- 花青素复合物比非共价复合物表现出更强的发泡能力,这可能要归因于共价相互作用可以更有效地提高蛋白质膜的稳定性。 凝胶特性与蛋白质基质的形成和维持有关,在一定条件下提高蛋白质凝胶的硬度、弹性、内聚性和咀嚼性等功能特性,可以有效地改善食品品质,例如肉类、凝乳和奶酪。组装聚集体的存在会嵌入可溶性蛋白质所形成的凝胶网络中,从而形成坚硬的凝胶[51]。Liu等[52]报道大豆分离蛋白经MTG作用组装后,其凝胶特性得到很大程度的改善。当溶液中存在不相容的多糖时,促进了蛋白质分子之间的吸引,并通过减少蛋白质分子与周围溶液接触的机会促进了大豆蛋白的分子排列,驱动蛋白质进行聚集组装,从而促进了蛋白质的凝胶化。Zhao等[53]发现,当大豆蛋白溶液中分别含有0.5%的魔芋胶、吉兰糖胶和可得然胶(curdlan gum)时,均可显著提升大豆蛋白的凝胶特性 。 基于蛋白组装的薄膜具有较低的水敏感性和较强的机械性能。Jensen等[54]使用大豆分离蛋白与纤维素以95∶5(g/g)的比例通过自组装获得了由大豆分离蛋白- 纤维素制成的复合薄膜。与大豆分离蛋白薄膜相比,这些源自自组装复合膜的拉伸强度和杨氏模量值显著提升,并且其断裂伸长率表现出更刚性的机械行为。 豆类蛋白具有较高的营养价值,含有高比例的赖氨酸、亮氨酸、天冬氨酸和精氨酸等必需氨基酸,已广泛应用于各类营养补充产品的加工中。豆类蛋白优异的食品加工特性使其可作为动物蛋白的替代品,在未来健康食品产业中发挥越来越重要的作用。豆类蛋白分子的组装是进一步开发利用豆类蛋白的重要策略。环境因素,如pH值、温度、离子类型和强度以及大分子物质会显著影响蛋白质的组装行为;蛋白组装体则会改善蛋白的乳化性、发泡性、凝胶性和成膜性等加工特性。 为了更高效地利用豆类蛋白,需要进一步研究豆类蛋白的分子结构、组装方式与酶解消化间的相关性,探讨豆类蛋白与其他食物成分的相互作用对其消化特性的协同或干扰,丰富豆类蛋白营养健康理论。同时,还需要对豆类蛋白食品的加工技术进行大力发展与深度革新,比如,根据不同豆类的蛋白质组成及营养特性选择合适的豆类加工品种;优化豆类蛋白的分子组装策略使其在宏观状态下呈现不同的质地结构,并实现营养和食品加工双特性的改善;更有效地依赖物理技术和酶技术提高豆类蛋白加工的绿色和健康属性;对传统食品加工技术进行改良以提高豆类蛋白终端产品的感官品质等等。这一系列工作的突破,无论是在传统食品的深度应用,还是未来食品的研究开发,豆类蛋白都将发挥重要作用。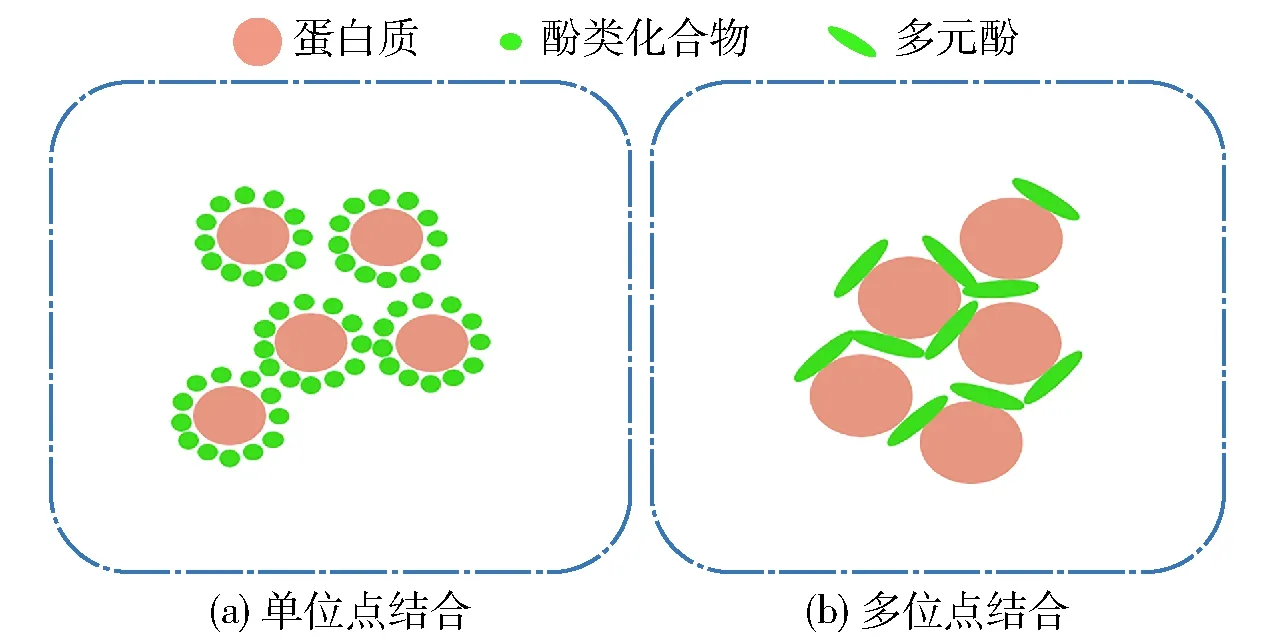
2 豆类蛋白组装行为与加工特性的关联性研究
3 展 望
