三叶木通大小孢子发生和雌雄配子体发育研究
2022-12-06姜治国陈发菊
白 洁 周 涛 马 江 刘 文 姜治国 陈发菊*
(1. 三峡大学生物技术研究中心,宜昌 443002;2. 神农架国家公园管理局神农架金丝猴保育生物学湖北省重点实验室,神家架林区
442421)
三叶木通(Akebia trifoliata)隶属于木通科(Lardizabalaceae)木通属(Akebia),是一种木质藤本植物,同时也是药食两用植物。三叶木通原产于中国和日本,在我国主要分布于秦岭或长江以南各省区[1-2]。三叶木通全株均可入药,疗效独特,应用广泛,具有利尿、镇痛、祛风湿、抑制炎症、抗衰老、抗疲劳、提高免疫力、抑制肿瘤和降血脂等疗效[3],常用作地方入药,其历史悠久,记载于民间传统中草药[4],干燥的藤茎和果实作为药用正品收录于《中国药典》2020 版(https://www.chp.org.cn/)。三叶木通果实又称八月瓜、八月札、八月炸、九月炸、北方土香蕉、预知子等[5],其果肉白色滑嫩,富含糖、维生素C和12种氨基酸,属于第三代保健型水果,具备优良的应用价值和开发前景[6]。
植物生殖生物学研究对象是整个有性生殖过程,由植物胚胎学演化而来,与之相关的过程包括花芽分化、雌雄性器官的发育、传粉受精以及胚胎发育等方面[7]。木通科生殖生物学的研究,最早始于对木通科演化进化轨迹进行分类系统学的研究[8],有关木通科大血藤属(Sargentodoxa)[9-10]、猫儿屎属(Decaisnea)[11]和串果藤属(Sinofranchetia)[12]植物的大小孢子发生及雌雄配子体发育研究已经积累了比较详细的胚胎学资料。植物生殖发育过程任一环节出现异常都显著影响其植物生殖的有效性,进而引发其自我更新过程受阻等问题[13]。生殖发育过程中的异常现象主要表现在受精前生殖细胞结构和功能的发育异常,从而导致花粉和胚囊的败育,进而可能影响其传粉受精,这是植物结实率低的重要致因之一[14-15]。
三叶木通花多果少,结实率低,其果皮普遍较厚,种子多,可食率低,果实成熟开裂不耐储藏等特点,严重限制其药用和食用产业化的发展[16-17]。
因此,提高三叶木通产量,培育出结实率高、品质好、可食率高的优良品种是亟待解决的问题。目前,国内外对三叶木通的研究主要集中在化学成分与药理作用[18]、组织培养[19]、分子标记技术[20]以及花和果实发育相关基因[21-22]等方面。然而,植物结实率与大、小孢子发生和雌、雄配子体的发育密切相关。三叶木通大小孢子发生及雌雄配子体发育鲜见报道,鉴于此,本文利用石蜡切片方法,观察三叶木通的大、小孢子发生及雌、雄配子体发育过程中胚胎学特征,明确雌花中雄蕊和雄花中心皮发生退化的关键时期,以期了解三叶木通有性生殖过程,分析该物种自然条件下结实率低的生殖原因,并为后续进行杂交育种和新品种培育提供理论基础。
1 材料与方法
野生三叶木通采自湖北省神农架国家公园木鱼镇官门山,地理坐标为31°26'56″N,110°23'59″E。于2020 年1 月花芽分化初期至5 月初开花期间不同时期取样,将不同发育时期的三叶木通早期花芽和雌雄花材料于FAA 固定液(70%乙醇90 mL、冰醋酸5 mL、38%甲醛溶液5 mL)中固定,带回实验室置于冰箱中4 ℃保存备用,采用石蜡切片方法,将固定好的材料放在解剖镜下用解剖针小心分离出雌雄蕊,依次通过(80%、85%、90%、95%、100%、100%)乙醇梯度脱水,每级2 h;依次经过1/2 二甲苯+1/2 乙醇、纯二甲苯、纯二甲苯透明,每级2 h,60 ℃下用液体纯蜡浸蜡12 h,每4 h 换1 次新蜡后,进行包埋,用Leica RM2125RT 切片机对包埋块进行连续切片,切片厚度8 μm;依次经展片、脱蜡、复水后,采用番红-固绿(Safranin-Fast Green)对染法进行染色,通过(95%、95%、100%、100%)乙醇脱水,二甲苯透明,中性树胶封片;用Leica DMR 多功能显微镜及测量系统观察拍照。
取散粉期花粉,经乙醇复水处理,用10%无水亚硫酸钠在沸水中软化1 h、水洗、苯胺蓝染色制片,用舜宇RX50 荧光显微镜下观察花粉的形态特征。
2 结果与分析
三叶木通为雌雄同株,单性花,总状花序,长10~12 cm,基部为1~2 朵雌花,上部为30~50 朵雄花(图版Ⅰ:A)。雄花具苞片4 枚,雄蕊6 枚,花托上呈单轮排列,成熟花药具2条纵沟,纵向开裂,雄花中退化心皮3~5 枚,位于花托中央,小而细长(图版Ⅰ:B)。雌花2 朵,位于花序轴基部,苞片4枚,每朵雌花中有离生雌蕊4~6 枚,柱头、花柱和子房发育正常,花柱极短,雌花中退化的雄蕊位于子房基部,极小(图版Ⅰ:C)。

图版Ⅰ 三叶木通的花序、雄花和雌花特征A.总状花序,示基部为2朵雌花(黑箭头所示),上部雄花(红箭头所示);B.雄花的形态,示雄蕊(st)与败育的心皮(ca);C.雌花的形态,示雌蕊(pi)与退化的雄蕊(se)PlateⅠ Floral morphology of A.trifoliata A.Raceme,showing 2 female flowers at the base and male flowers at the upper part;B.Male flower morphology,showing stamens and aborted carpel;C.Female flower morphology,showing pistils and staminode
2.1 小孢子发生和雄配子体发育
2.1.1 花药壁发育
三叶木通雄花具有雄蕊6 枚,每枚雄蕊具4 个小孢子囊,两侧小孢子囊由药隔相连,药隔由薄壁细胞和维管束组成(图版Ⅱ:A)。花药原基最外层有一层表皮细胞,表皮以内4个角隅处形成孢原细胞(图版Ⅱ:B)。表皮下的一个孢原细胞经过1 次平周分裂,形成内外2层子细胞,向外为周缘细胞,即初生壁细胞,向内生成体积较大的初生造孢细胞,初生壁细胞再进行平周和垂周分裂,由外向内产生4~5层细胞,逐渐形成1层药室内壁、2~3层中层和1层绒毡层,此时花药壁各层细胞区别并不明显(图版Ⅱ:C)。花药壁细胞继续生长,早期的绒毡层细胞多为单核,细胞质浓厚。初生造孢细胞经过数次有丝分裂,产生多边体形排列的次生造孢细胞(图版Ⅱ:D)。随着花药壁的发育,当次生造孢细胞进一步发育成小孢子母细胞时,花药壁分化完全,即表皮下的4~5层细胞逐渐分化为1层药室内壁,2~3 层扁平状的中层细胞,最内层为多面体型的绒毡层细胞,细胞体积大,原生质浓厚,往往是双核,甚至多核。发育完全的花药壁内是小孢子母细胞(图版Ⅱ:E),由此可以看出,三叶木通花药壁的发育为基本型。小孢子四分体时期,药室内壁沿径向壁和切向壁增厚,并产生不均的条状或略微木质化结构,中层细胞体积开始解体,绒毡层细胞发达,开始出现解体的迹象(图版Ⅱ:F);随着小孢子进一步发育,药室内壁细胞进一步径向延长,最内一层中层细胞首先被吸收,绒毡层不断为小孢子提供充足的营养,细胞壁变得模糊不清,细胞核消失,逐渐原位解体被吸收,属于分泌型绒毡层(图版Ⅱ:G);随着花药发育成熟,花药壁仅剩下纤维状加厚的药室内壁及残留的表皮细胞,绒毡层和中层细胞逐渐完全退化(图版Ⅱ:H~I)。

图版Ⅱ 三叶木通花药壁发育A.花药横切面,箭头所示4个小孢子囊;B.孢原细胞平周分裂;C.未分化的4~5层花药壁细胞和次生造孢细胞形成;D.发育中的花药壁和早期小孢子母细胞;E.双核或多核绒毡层细胞;F.花药壁和小孢子四分体;G.绒毡层细胞开始退化;H~I.纤维状加厚的药室内壁和成熟花粉粒;an.花药;ac.孢原细胞;sc.次生造孢细胞;aw.花药壁;ep.表皮;en.药室内壁;ml.中层;ta.绒毡层;fw.纤维状加厚的药室内壁;pg.花粉粒PlateⅡ Anther wall development of A.trifoliata A.Anther transverse section,arrow showing four sporangiate;B.Archesporial cell periclinal division;C.Undifferentiated 4-5 layers of anther parietal cells and secondary sporogenous cells;D.Developing anther wall and early microspore mother cells;E.Binucleated or multinucleated tapetum cells;F.Anther wall and microspore tetrad;G.Tapetum cells begin to degeneration;H-I.Fibrous endothecium and mature pollen;an.Anther;ac.Archespo⁃rial cell;sc.secondary sporogenous cells;aw.Anther wall;ep.Epidermis;en.Endothecium;ml.Middle layer;ta.Tapetum;fw.Fibrous endothecium;pg.The pollen grains
2.1.2 小孢子发生及雄配子体发育
三叶木通小孢子发生是由小孢子母细胞经过减数分裂产生,小孢子母细胞体积大,细胞核大,核仁明显,细胞质浓厚(图版Ⅲ:A)。随着周围胼胝质的形成,小孢子母细胞经过第1次减数分裂后形成双核细胞,不形成二分体时期,完成第2次减数分裂后形成四分体,四分体大多为四面体型排列,三叶木通小孢子母细胞胞质分裂为同时型(图版Ⅲ:B~C)。
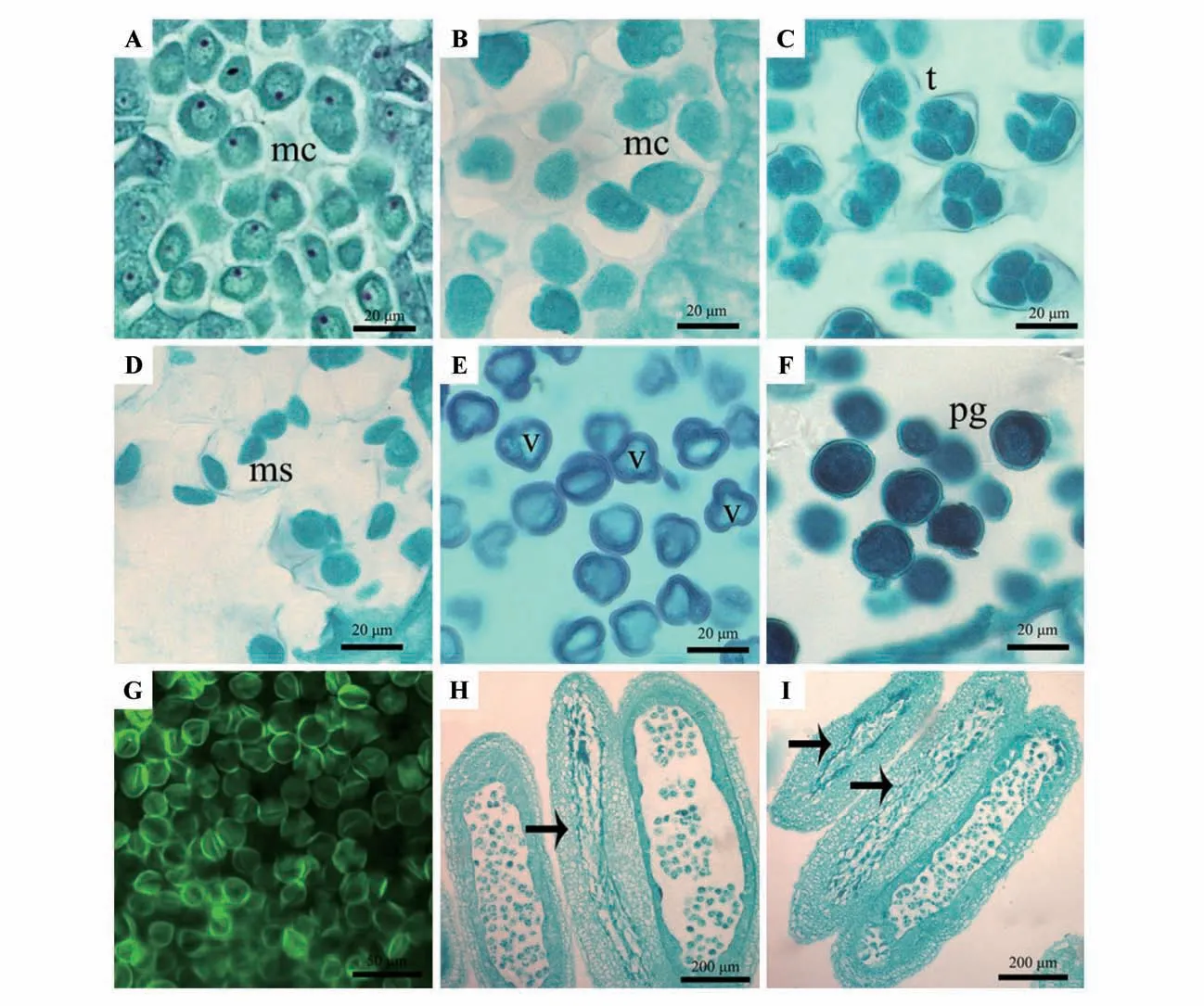
图版Ⅲ 三叶木通小孢子发生及雄配子体发育A.小孢子母细胞;B.小孢子母细胞减数分裂Ⅱ后期;C.四分体;D.胼胝质溶解;E.单核靠边花粉;F.成熟花粉粒;G.荧光显微镜下的花粉形态;H.3 个花粉囊,示中间一个有花粉败育现象;I.3 个花粉囊,示左上方2 个有花粉败育现象;mc.小孢子母细胞;t.四分体;ms.小孢子;v.液泡;pg.花粉粒Plate Ⅲ Microsporogenesis and male gametophyte development of A.trifoliata A.Microspore mother cells;B.Anaphase of meiosis Ⅱof the microspore mother cells;C.The tetrad;D.Callose dissolution;E.Uninucleate pollen;F.Mature pollen grains;G.Pollen morphology in the fluorescence microscope;H.Three pollen sasc,showing the middle one has pollen abortion;I.Three pollen sacs,showing two on the upper left have pollen abortion;mc.Microspore mother cells;t.The tetrads;ms.Microspores;v.Vacuole;pg.The pollen grains
三叶木通形成的小孢子四分体外部包裹着1 层厚厚的胼胝质壁,随着胼胝质壁溶解消失,小孢子释放到药室中,形成4 个单核的小孢子(图版Ⅲ:D)。接着小孢子体积逐渐增大,细胞质发生液泡化,并形成一个大液泡,将小孢子中的细胞核挤向一边,形成单核靠边期(图版Ⅲ:E)。单核小孢子细胞壁加厚,较厚的内外壁结构依稀可见,核仁增大而显著,经过有丝分裂形成一个大的营养核和一个小的生殖核,最后发育形成2-核成熟花粉粒即雄配子体(图版Ⅲ:F)。荧光显微观察下,成熟花粉壁显绿色荧光,赤面观呈椭圆、圆形,极面观呈6个明暗相间的三角形(图版Ⅲ:G)。
同时可以观察到三叶木通花药发育过程中有少量败育现象,自小孢子母细胞减数分裂后,花粉囊各层花药壁细胞排列层次不清、紊乱,绒毡层细胞发育不正常,并延时降解,小孢子四分体因得不到营养而收缩降解(图版Ⅲ:H~I),成熟花粉圆球形染色较深的为可育花粉,花粉皱缩空壳为败育花粉。
2.2 大孢子发生和雌配子体发育
从子房纵切面来看,三叶木通具1 室子房,为侧膜胎座,胚珠多枚(图版Ⅳ:A)。子房发育早期,位于腹缝线处出现分裂能力极强、呈团状整体突起的一群细胞,称为胚珠原基。其顶端为珠心组织,基部发育成株柄,并连接着胎座。在胚心组织表皮下分化出一个细胞体积增大,细胞核大,细胞质浓厚的特殊细胞,称为雌性孢原细胞(图版Ⅳ:B),孢原细胞继续生长并分裂形成一个周缘细胞和一个造孢细胞(图版Ⅳ:C)。周缘细胞经过平周和垂周分裂形成珠心组织细胞,造孢细胞进一步发育为大孢子母细胞,其细胞体积大,细胞核显著,明显区别于其他薄壁细胞,大孢子母细胞发育过程中,靠珠心基部发生突起,形成内外2层珠被,内珠被发生先于外珠被,在胚珠顶端不愈合留下一个开口,称为珠孔(图版Ⅳ:D)。在此过程中,胚珠由最初的直立生长慢慢向一侧发生弯曲生长,大孢子母细胞减数分裂后形成线型排列的大孢子四分体,与小孢子发育不同的是,大孢子四分体中,靠近珠孔端的3 个大孢子逐渐变小退化,靠近合点端的1 个大孢子最先发育,体积变大,慢慢移向珠孔端,成为功能大孢子(图版Ⅳ:E)。功能大孢子被包围在数层珠心组织细胞之中,所以三叶木通胚珠发育为厚珠心类型。

图版Ⅳ 三叶木通大孢子发生及雌配子体发育A.子房纵切;B.雌性孢原细胞;C.周缘细胞产生1层珠心细胞时,示造孢细胞及内珠被原基;D.大孢子母细胞时期,示厚珠心胚珠;E.珠孔端的3 个大孢子相继退化,合点端功能大孢子的形成;F.二核胚囊;G.八核胚囊;H-I.卵器,示1 个卵细胞及2 个助细胞;J.上下极核;K.合并中的上下极核;L.受精极核;缩写:asc.孢原细胞;mmc.大孢子母细胞;ii.内珠被;oi.外珠被;dm.退化大孢子;fm.功能大孢子;bes.二核胚囊;ac.反足细胞;cc.中央细胞;ec.卵细胞;sy.助细胞;pn.极核;fn.受精极核Plate Ⅳ Megasporogenesis and female gametogenesis of A.trifoliata A.Ovary longitudinal section;B.The archesporial cell differentiated immediately below the nucellarepidermis;C.The Peripheral cells produce a layer of nucellus cells,showing sporogenous cells and inner integument primordia;D.Megaspore mother cell stage,showing thick nucellus ovule;E.The 3 megaspores at the end of the nucleus degenerate one after another,and the formation of functional megaspores at the chalazal end;F.Binucle⁃ate embryo sac;G.Eight nuclear embryo sac;H-I.Egg apparatus,showing an egg cell and two synergia;J.The polar nuclei which is in the process of fusion;K.The second nucleus of central cell;L.Fertilization of polar nuclei;asc.Archesporial cell;mmc.The mother megaspore cell;ii.The inner in⁃tegument;oi.The outer integument;dm.The degenrated megaspores;fm.The functional megaspore;bes.Binucleate embryonary sac;ac.Antipodal cell;cc.Central cells;ec.Egg cell;sy.Synergids;pn.Polar nucleus;fn.Fertilization of polar nuclei
三叶木通功能大孢子发育形成单核胚囊,随即经过第1 次有丝分裂,形成2 个游离核,即进入二核胚囊期(图版Ⅳ:F),继续经过2 次有丝分裂,最终形成七细胞八核胚囊期(图版Ⅳ:G)。珠孔端分化出1 个卵细胞和2 个助细胞,构成卵器,呈“品”字型排列(图版Ⅳ:H~I),2 个上下极核互相融合生成次生核的中央细胞(图版Ⅳ:J~K),靠近合点端的为3 个反足细胞充当吸收,转运和分泌营养物质到胚囊的作用。受精后,三叶木通成熟胚囊中2 个助细胞逐渐退化,合点端的3 个反足细胞解体,中央细胞受精形成受精极核(图版Ⅳ:L)。因此,三叶木通经3 次连续的有丝分裂并进一步发育为七细胞八核的蓼型胚囊,在雌配子体发育过程中,胚珠生长继续弯曲,珠心和珠被的纵轴与珠柄近成直角,最后成熟胚珠为横生胚珠(图版Ⅳ:G)。
2.3 雌花中的雄蕊和雄花中的心皮发育
三叶木通雌花中雄蕊的早期发育与正常雄花中的相似,小孢子母细胞能进行减数分裂,但小孢子进一步发育过程中出现异常现象,具体表现为大部分小孢子发育停留在单核靠边期(图版Ⅴ:A),大部分小孢子发生败育,药室内壁也发生纤维状加厚,但不如正常雄花中的显著,中层细胞始终不退化,绒毡层细胞退化不完全,可见残余痕迹。即使有少部分花粉发育成熟,但花药壁不开裂(图版Ⅴ:B)。其外形与雄花中的雄蕊相比,颜色为嫩绿色,发育不良,被雌蕊挤压在基部,形成功能性雌花(图版Ⅰ:C)。

图版Ⅴ 三叶木通雌花中雄蕊和雄花中心皮的发育A.单核小孢子;B.雌花中的雄蕊花药壁不开裂,示中层不退化,绒毡层退化不完全;C.雄花中4枚退化心皮;ms.小孢子;ca.心皮PlateⅤ Development of stamens in female flowers and carpel in male flowers of A.trifolia A.Uninucleate microspore;B.Stamens in female flowers anther wall does not crack,showing the middle layer is not degraded,the tapetum layer is not completely degraded;C.4 degenerated carpels in the male flowe;ms.Microspores;ca.Carpel
三叶木通雄花在雄蕊从花药造孢细胞、小孢子四分体以及此后花粉成熟的过程中,雄花中的心皮愈合不完全,心皮不产生胚珠,体积虽有增大,心皮早期未充分发育而造成退化(图版Ⅴ:C)。其外形与雌花中雌蕊相比,花柱颜色为嫩绿色,柱头未膨大,形成功能性雄花(图版Ⅰ:B)。
3 讨论
三叶木通雄蕊每枚花药具4个小孢子囊,小孢子母细胞胞质分裂属于同时型,小孢子四分体呈“品”字四面体型排列,四分体解体后,形成单核花粉。中层和绒毡层在小孢子的发育过程中最后解体,成熟花粉为2 核细胞。三叶木通子房1 室,侧膜胎座,胚珠横生,多枚,双珠被,厚珠心,胚囊为单孢子发生七细胞八核蓼型胚囊。这些发育特征与木通科其他属植物发育方式既有同一性又存在差异性[9-12],胚珠发生方式与之前报道的弯生和倒生类型不同[23],本次试验观察发现,三叶木属于典型的横生胚珠类型。三叶木通具有一些较为原始的胚胎学特征,2-核成熟花粉、单孢子发生的蓼形胚囊等,同时,三叶木通也具有较为进化的胚胎学特征,横生胚珠,绒毡层为较进化的分泌绒毡层,表明三叶木通的胚胎学特征可能处于由原始到进化的过渡地位。
木通科植物花性复杂,雌花的花药退化和雄花的心皮退化是常见的现象[24],其退化的时期及功能性有所不同,八月瓜(Holboellia latifolia)退化雄蕊发育到单核花粉阶段,导致花粉败育[25]。在单叶血藤(Sargentodoxa simplicifolia)两性花中的退化雄蕊,大部分小孢子发育到单核花粉期,少数能产生2 或3 细胞花粉,花药壁并不开裂[10]。串果藤(Sinofranchetia chinensis)退化雄蕊小孢子发育到四分体时期退化,少数能产生2 或3 细胞花粉,退化雄蕊在花部结构、开裂方式及花粉形态上与正常雄蕊无明显差别[12]。通过本次显微结构观察发现,三叶木通雌花中雄蕊退化,大部分小孢子发育停留在单核靠边期,即使有少部分发育成2-核成熟花粉,但花药壁不开裂,雄花中心皮退化是由于早期未发育所致。因此,三叶木通为一种功能上的单性花,并且认为此花比猫儿屎属、串果藤属和大血藤属植物中的进化,三叶木通胚胎学支持木通属和木通科各属具有相似的系统发育过程,其进化方向和水平具有各自不同的特点[8]。这些细胞学研究结果不仅反映了三叶木通的祖先繁育状态,而且为研究三叶木通单性花形成的分子机制提供理论证据,也为三叶木通种群进化趋势提供了线索。
三叶木通大、小孢子发生和雌、雄配子体发育大多基本正常,小孢子发育有少量败育现象,但发生的概率极低,大多能够正常发育成熟。前期授粉试验证明,三叶木通不仅可作为授粉树种,也可作为杂交母本的优良选择。木通科其他属植物的大、小孢子发生和雌、雄配子体发育过程中未见胚珠、卵器和小孢子发育异常的报道,可见三叶木通结实率低的胚胎学原因并非大、小孢子发生和雌、雄配子体发育异常。在花药间及不同药室内小孢子和同一子房中的胚珠的发育进程出现不同步的现象,大约相差1~2 个时期,这种发育不同步现象对增加花粉活性、延长授粉时间及促进异交有一定的帮助,但对于野生三叶木通分布零散且不集中情况下,可能会导致结实率降低。传统观念认为植物雌雄性功能在时间上或空间上的分离是广泛阻止花内自花授粉的有效机制,退化雄蕊是避免自交的适应机制[26],日本木通的传粉生物学研究表明木通具有自交不亲和性,即自花传粉或相同基因型的异花传粉而导致不育或结实率较低[27-28]。此外,影响结实率包括自然和人为等外部因素,野生三叶木通多生长在海拔1 200~1 800 m的公路旁、河边、山坡或山沟林中,常以单株或几株攀爬于乔木和灌木混杂的植物群落里,花期3~5月,神农架地区昼夜温差大且多雨,这些自然因素对于三叶木通传粉,结实也有影响,此时也正值旅游旺季,人为影响同样也不可避免。因此,三叶木通雌雄同株异花,是一种较为严格的异花授粉植物,在自然条件下难以完成传粉,其结实率低可能与影响传粉的外部因素有关,对三叶木通自交不亲合性的类型和特点有待进一步研究,将有助于深入解答其“花多果少”的原因,从而为三叶木通开发利用提供科学依据。
