基于单拷贝核基因的褐沙蒿遗传结构和谱系地理学分析
2022-12-06唐思如王铁娟刘梦瑶
唐思如 王铁娟 秀 芳 刘梦瑶 张 慧
(内蒙古师范大学生命科学与技术学院,内蒙古自治区高等学校生物多样性保护与可持续利用重点实验室,呼和浩特 010022)
谱系地理学(phylogeography)最早由Avise 提出,是通过重建基因谱系,揭示物种形成现有地理分布格局的成因,以及近缘种间或种群间的历史进化关系[1-3]。对某个物种的谱系地理学研究,更多关注该物种冰期避难所及在气候变化过程中物种范围的变化等进化历史问题[4]。在我国,谱系地理学的研究以横断山区和毗邻的青藏高原地区为热点,对干旱区的研究也有一定报道,见于我国西北部荒漠区的濒危植物或一些特有植物的研究[5-9]。草原区沙地的相关研究少见,如Xu 等[10]对科尔沁沙地分布的东北木蓼(Atraphaxis manshurica)进行谱系地理学研究,提出该种的遗传分化主要发生在更新世中晚期,与周期性的冰期和季风变化相一致。
褐 沙 蒿(Artemisia intramongolica)为 菊 科(Compositae)蒿属(Artemisia)龙蒿组(Sect.Dracun⁃culus)半灌木植物,该种集中分布于内蒙古典型草原带的浑善达克沙地,是该区域固定沙地和半固定沙地的主要建群植物,也可见于覆沙高平原上[11]。褐沙蒿与分布于科尔沁沙地与呼伦贝尔沙地的差不嘎蒿(A.halodendron)和分布于鄂尔多斯及其周边的黑沙蒿(A. ordosica)构成典型的沙生地理替代分布[12],为我国北方重要的防风固沙、水土保持植物,其嫩枝与叶为家畜喜食,具有一定的饲用价值。对于褐沙蒿的遗传分化与多样性研究有过一定报道,集中在片段分析方面,包括RAPD[13]、ISSR[14]、SSR[15]3 种分析方法,而基于序列分析的谱系地理学研究还未见报道。作为浑善达克沙地的特有植物和最为重要的固沙植物之一,其形成与分化的时空格局及相关的地球环境背景尚不清楚,值得深入探讨。本研究基于转录组开发得到的褐沙蒿及其近缘种的2 个直系同源单拷贝核基因片段,分析褐沙蒿的遗传结构以及谱系分布格局,探讨褐沙蒿的种群扩张历史,为褐沙蒿以及其近缘种间的进化迁移提供重要资料,也为浑善达克沙地形成对其现有物种的地理分布格局的影响研究提供参考。
1 材料与方法
1.1 试验材料
在褐沙蒿自然分布区范围内共选取8 个地理种群,地点与生境见表1 和图1。在每个种群随机选取20 个个体,个体之间间隔10 m 以上。每株植物采集生长良好的嫩叶约10 g,放入已编号密封袋中加变色硅胶快速干燥保存,带回实验室后置于冰箱中-20 ℃存放。8个种群的凭证标本均存放在内蒙古师范大学生命科学与技术学院标本馆。

图1 褐沙蒿采样分布图地图信息来自国家基础地理信息中心(https://www.webmap.cn)Fig.1 Sampling distribution map of A.intramongolica The map information is from the National Geomatics Center of China(https://www.webmap.cn)

表1 采样地点与生境Table 1 Sampling sites and habitats
1.2 研究方法
1.2.1 同源单拷贝核基因开发
利用Illumina HiSeq 4000 测序平台(北京百迈克公司)对蒿属龙蒿组10 种植物样品进行转录组测序,得到Clean Data,利用Trinity 软件[16]组装得到大量Unigene,借助OrthoMCL[17]软件根据Blast的结果,通过mcl算法分别对样品进行基因家族聚类分析,得到5 000 多条直系同源单拷贝核基因。由于褐沙蒿与黑沙蒿、差不嘎蒿为近缘种[18],筛选在3 种植物间具有2 个以上变异位点的基因,由北京擎科生物科技有限公司进行引物合成。
1.2.2 DNA提取
采用北京天根生化科技公司的DNAsecure 新型植物基因组DNA 提取试剂盒提取样品总DNA,然后利用0.7%琼脂糖凝胶电泳(1xTBE)分离并检测条带,用超微量紫外分光光度计检测浓度后,将DNA 产物统一稀释至40 ng·μL-1,于-20 ℃保存备用。
1.2.3 引物筛选与PCR扩增
通过NCBI网站中的Primer-Blast合成的引物,然后在褐沙蒿种群内进行预试验,选择扩增效果好、多态性较高的基因c9065和c7847,c9065引物序列是F:5'-TTCATTGACATGGGAAAGAAATCA-3'和R:5'-TGGTTGCTTCAAATCGTCCG-3',c7847为F:5'-ATGTTCATGCTGACCCCAAT-3'和R:5'-CCGGATCCTCCATCGATAAC-3'。对 褐 沙 蒿8 个种群个体进行PCR 扩增,反应体系为25 μL:2TaqMasterMix 12.5 μL,DNA模板2.0 μL(80 ng),ddH2O 8.5 μL,引物各1.0 μL(10 μmol·L-1)。PCR 扩增程序:70 ℃预变性1 min,94 ℃变性1 min,55 ℃退火20 s,72 ℃延伸50 s,循环2次;接着94 ℃变性20 s,55 ℃退火20 s,72 ℃延伸50 s,循环38 次;最后72 ℃延伸5 min,4 ℃保存。扩增产物由北京擎科生物科技有限公司完成测序。分析所有个体序列的双峰位点,按照双峰位点组成类型,选择相应个体克隆测序,以对软件拆分的单倍型序列进行矫正。
1.3 数据分析
通 过Chromas 软 件(http://technelysium.com.au/)查看测序峰图,对于杂合位点按照IUPAC 编码标准(R=A/G;Y=C/T;M=A/C;S=C/G;W=A/T)录入;利用DNASTAR Lasergene v7.1 软件[19]进行序列比对;用MEGA7 软件[20]将序列前后切齐,并进行手工校正,序列文件以Fasta 格式保存;由于是核基因,具有重组,使用DnaSP 5.10 软件[21]中的UNPHASE/Genotype 功能将序列拆分,生成单倍型,并通过克隆测序结果对单倍型进行验证,统计单倍型数量(h)、计算单倍型多样性(Hd)、核苷酸多态性(π)与基因流(Nm)等指标;通过Arleqiun3.5软件[22]进行分子变异方差分析(AMOVA),统计种群内和种群间的遗传变异(1 000 次置换检验);由PERMUT 软件[23]计算种群遗传分化系数Gst、Nst值(1 000 次置换检验);利用Arleqiun3.5 软件计算中性检验的Tajima’sD、Fu’Fs、SSD、Raggedness index值,通过DNAsp 5.10 绘制错配图;利用软件PO⁃PART[24]中的Median Joining Network 模型构建单倍型网络关系图。
2 结果与分析
2.1 遗传多样性分析
褐沙蒿c9065片段长度为485 bp,116 个个体共获得14 个变异位点和48 种单倍型(GenBank 注册序列号:OL632106-OL632153);c7847片段长457 bp,147 个个体共获得19 个变异位点和40 种单倍型(GenBank 注册序列号:OL632154-OL632193)。褐沙蒿在种的水平上,c9065的Hd为0.871 6,π为0.008 2,c7847的2个值分别为0.934 8和0.005 9;在种群水平上c9065的Hd为0.808 5~0.931 2,π为0.007 2~0.009 0,c7847的Hd为0.884 1~0.9524,π为0.005 3~0.006 6。按照Grant等[25]提出的标准,Hd>0.5、π>0.005,多样性程度高(见表2)。因此,褐沙蒿不论是物种水平还是各个种群均具有较高的多样性。
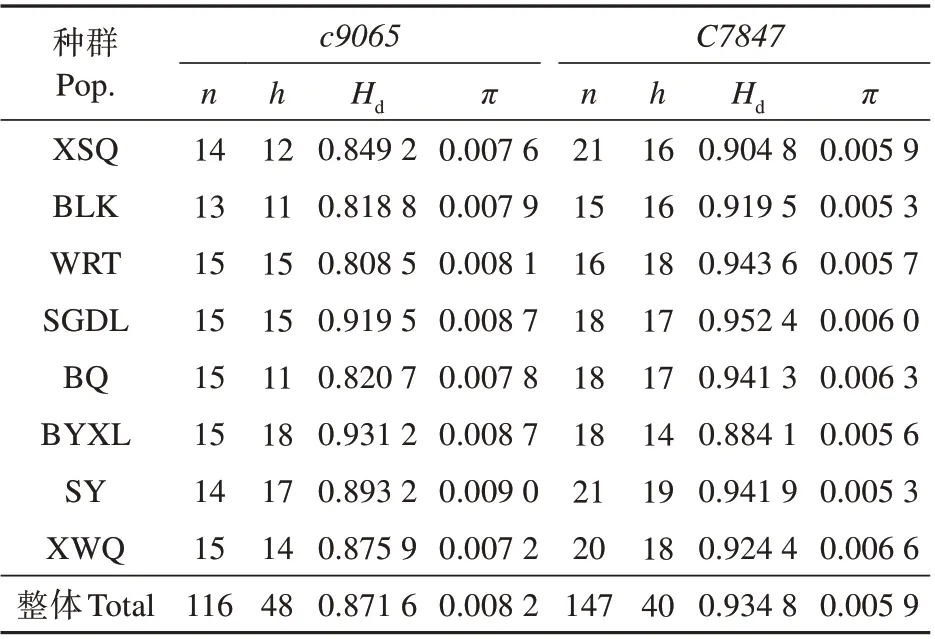
表2 褐沙蒿8 个种群的单倍型多样性(Hd)与核苷酸多样性(π)Table 2 Haplotype diversity(Hd)and nucleotide diversi‐ty(π)of eight populations of A.intramongolica
2.2 遗传结构与单倍型网络关系
分子方差分析结果表明,基于c9065和c7847的褐沙蒿种群内变异分别占99.398%和98.908%,表明绝大多数遗传变异来源于种群内(见表3),也因而有着很小的遗传分化,Fst分别为0.006(c9065)和0.011(c7847),处于Hartl 提出的分化很弱范围(Fst介于0~0.05)[26]。基因流Nm能反映种群间基因交流的情况,Nm≥1 为高水平[27],褐沙蒿c9065和c7847检测的基因流分别为6.810 和7.270(见表4),远远大于1,说明其种群间基因交流非常广泛,减少了种群间的遗传分化,与Fst的结果相互印证。从谱系地理结构来讲,c9065的分析结果Nst(0.007)

表3 基于c9065和c7847的褐沙蒿分子方差(AMOVA)分析Table 3 Analysis of molecular variance(AMOVA)of A.intramongolica based on c9065 and c7847

表4 褐沙蒿种群间遗传分化系数与基因流Table 4 Genetic differentiation coefficient and gene flow of A.intramongolica
图2为褐沙蒿的单倍型网络图,其中每个圆代表一种单倍型,圆的半径与该单倍型的数量呈正比,每种颜色分别代表1 个种群。2 个核基因的结果均表现为褐沙蒿网络图较为复杂,交错连接,主要分为两大分支A 和B。c9065结果中,Hap1 在各种群均出现,且出现的次数最多,其次为Hap2,也在各种群均出现;而分布少的单倍型多,有17种在2~4 个种群中出现,24 种为种群特有单倍型(见图2A)。c7847结果中,Hap2、Hap3、Hap4、Hap5、Hap7、Hap9及Hap12为8个种群共有单倍型,其中Hap5 出现次数最多,有16种单倍型在种群中出现的次数较少,只在2~4 个种群中出现,13 个为各种群特有单倍型(见图2B)。从各单倍型的分布情况看,均未与种群的地理分布相关。
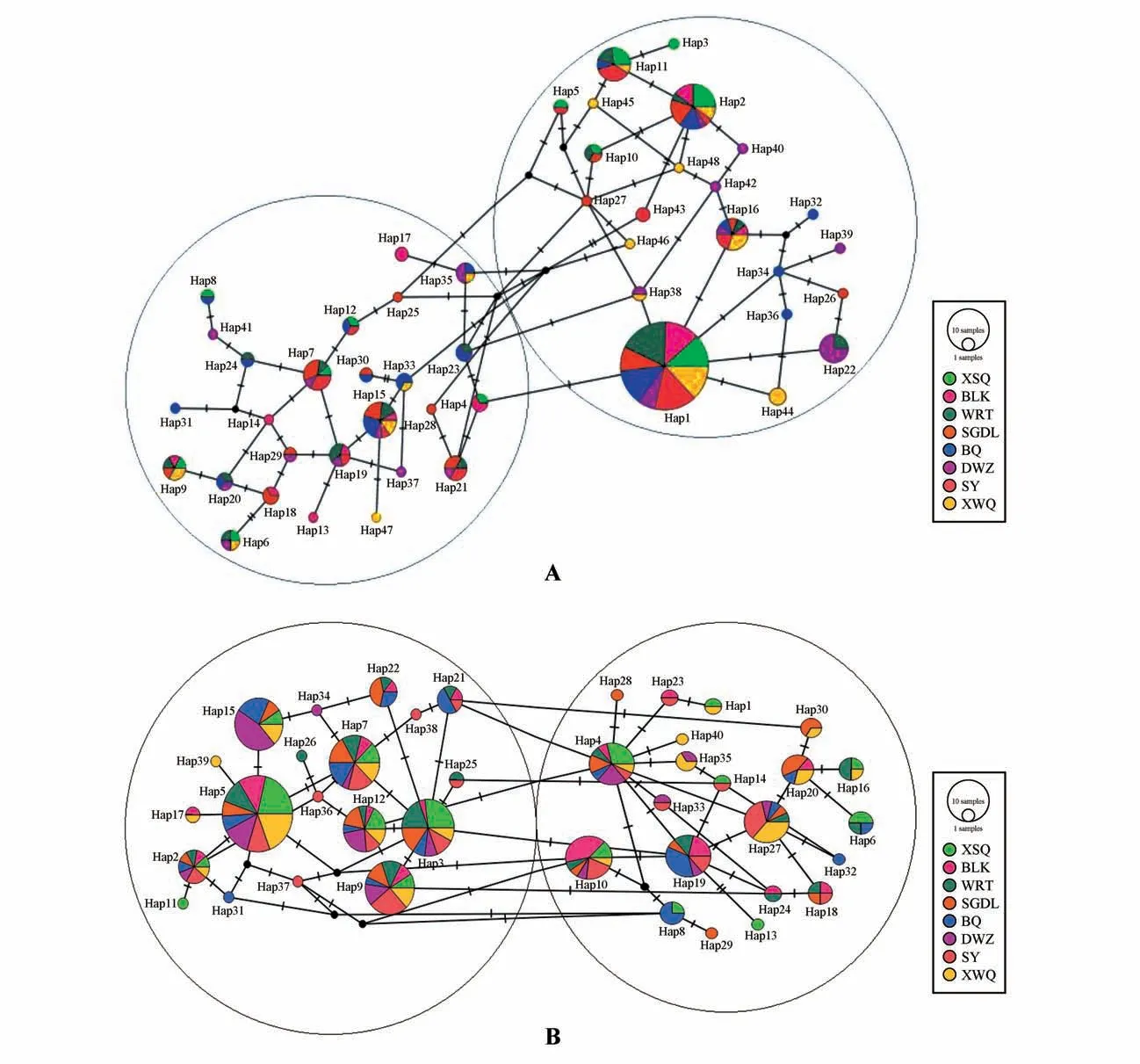
图2 基于单拷贝核基因c9065(A)与c7847(B)的单倍型网络Fig.2 Haplotype network based on the single-copy nuclear gene c9065(A)and c7847(B)
2.3 褐沙蒿地理种群中性检验及种群扩张分析
对褐沙蒿所有个体和种群进行中性检验表明,2 个基因的Tajima’sD值无论是总的还是种群水平均不显著(P>0.05),不拒绝中性进化假设。两个基因Fu’sFs总的值均显著(P<0.05),显示存在种群扩张(见表5)。从错配分析图来看,c7847呈单峰,支持Fu’sFs的结果,但c9065呈双峰(见图3)。综合各指标结果,表明褐沙蒿不存在明显种群扩张。歧点分析结果显示,c9065在总体和种群水平上SSD 值与Raggedness index 值均不显著(P>0.05),c7847两个值在总体水平上均显著,种群水平上多数显著(P<0.05)(见表5)。综合分析各种群的Fu’sFs值、SSD 和Raggedness index 值,位于分布区东北部的种群BYXL 与XWQ 在2 个基因上均显示Fu’sFs值显著(P<0.05)、SSD 和Rag⁃gedness index 值不显著(P>0.05),说明存在明显的种群扩张,位于西北部的XSQ 种群多数指标也显示存在种群扩张。综上所述,褐沙蒿在中性模式下经历了有限的扩张。

表5 褐沙蒿种群扩张概率Table 5 Probabilities statistics for population expansion of A.intramongolica

图3 基于单拷贝核基因c9065(A)与c7847(B)褐沙蒿种群错配分析Fig.3 Mismatch analysis diagram for populations of A.intramongolica based on the single-copy nuclear gene c9065(A)and c7847(B)
3 讨论
3.1 种群遗传多样性与遗传结构
以往对褐沙蒿种群遗传多样性的研究,无论是RAPD[13]、ISSR[14]还是SSR[15]有着一致的结果,均显示褐沙蒿有着高的遗传多样性。本研究通过转录组开发得到变异适中的直系同源单拷贝核基因c9065和c7847,分别对褐沙蒿自然分布范围内8 个地理种群116 个、147 个个体进行单倍型分析,结果显示,2个基因的单倍型多样性不论是总的还是种群均大于0.5;总的核苷酸多样性均在0.005以上,表明不论是物种还是种群均有着高的遗传多样性。这与之前基于各种片段分析的研究结果具有一致性,反映出虽然褐沙蒿分布范围不大,但作为蒿属这一进化类群中的一员对现有生存环境有着良好的适应性。
就种群遗传结构而言,基于c9065和c7847的AMOVA 分析显示,褐沙蒿种群内变异分别为占99.398%和98.908%。与以往的研究相比,RAPD分析显示褐沙蒿种群内变异占87.96%[13]、ISSR 为92.43%[14]、SSR 为86.02%[15],结果具有一致性,即褐沙蒿的变异主要由种群内的遗传变异导致。只是核基因显示出种群间变异更小,这与核基因得到的变异位点不如片段分析(RAPD、ISSR、SSR)得到的位点丰富有关。褐沙蒿分布于浑善达克沙地,该沙地地势较为平坦,没有明显的地理阻隔,加之褐沙蒿为风媒传粉,而花粉扩散正是基因流动的主要形式[28],因而得到褐沙蒿种群间的基因流很高,分别为6.810(c9065)和7.270(c7847),远远大于1,也正是广泛的基因交流导致褐沙蒿的种群间存在很小的遗传分化。
3.2 谱系地理结构与种群历史动态分析
植物谱系地理结构分析中常用的是叶绿体基因片段(cpDNA)和核基因中的ITS[5,29],但这些常用的片段在蒿属尤其是近缘种间分辨率不够,如我们曾经做过核基因ITS[30]、叶绿体基因片段trnL-F[31],以及未发表的多个叶绿体基因片段,结果显示在蒿属近缘种间具有高度的一致性,在种内个体间基本一致,因而进行谱系地理学研究具有一定的局限性。本研究以褐沙蒿及其近缘种为对象,开发得到大量的直系同源单拷贝核基因片段,选择变异适中的基因c9065和c7847,基因片段长485、457 bp,易于扩增,分别获得14、19 个变异位点,较丰富,不仅适合褐沙蒿研究,也可用于蒿属其他物种研究。本研究通过2 个核基因计算遗传分化系数Gst和Nst,并进行1 000次的置换检验,显示c9065的Nst
蒿属的化石孢粉最早见于亚洲北部第三纪渐新世的地层中[32],林有润提出蒿属植物在北半球形成分化后,沿着西、东及南三方向迁移而扩展分布区[33],在我国形成“三斜带密集带谱”,褐沙蒿所属的龙蒿组植物处于第三条带谱,即从大兴安岭西部至西藏西北部以北的干旱或半干旱地区[32]。据孢粉相关报道,我国北方蒿属植物主要出现在中新世时期[34-36],与前述相吻合。褐沙蒿分布于浑善达克沙地,浑善达克沙地是在晚新生代青藏高原隆升的大背景下形成演变的,第四纪冰期时,该区域受东亚季风控制,伴随着气候的巨变,出现沙丘活化、扩展与沙丘固定、收缩的波动过程[37],这必将对植被产生的影响。不过从褐沙蒿的研究结果来看,总体水平上,2 个核基因的Tajima’sD值均不显著(P>0.05),Fu’sFs值显著(P<0.05),呈现出在中性模式下经历了有限的种群扩张。从各种群来看,位于分布区东北部的西乌旗(XWQ)和锡林浩特东的BYXL种群存在明显的种群扩张、西北部的西苏旗(XSQ)种群也存在较明显的种群扩张,反映出分布区北部种群具有更大的波动性。Xu 等[10]对科尔沁沙地(与浑善达克沙地相邻)分布的东北木蓼进行种群历史动态研究,发现有其中2个种群的遗传多样性水平明显高出其他种群,因而推测这2 个种群的所在地可能是该种在冰期时的避难所,但本研究中褐沙蒿的各种群多样性水平差异不大,以往的研究也是如此,因而无法推断小型避难所的区域,尚需浑善达克沙地其他物种的研究。
