水青树特殊管胞的分布位置及其形态特征的研究
2022-12-06李茜然郑欣欣石江涛
李茜然 勇 璐 潘 彪 郑欣欣 石江涛 李 婷
(南京林业大学材料科学与工程学院,南京 210037)
水青树(Tetracentron sinense)是一种濒危的落叶乔木,起源于白垩纪或三叠纪早期,隶属于昆栏树目(Trochodendrales),属于真双子叶植物中较早分化形成的四个支系之一。它分布于中国的陕西、甘肃、湖北、四川、贵州、云南等省及印度的北部、缅甸北部、尼泊尔和不丹。由于采伐破坏、性状变异等[1-3]原因,水青树数量骤减,被列为国家Ⅱ级重点保护濒危野生植物[4]。从叶子和树干的形态来看,水青树是一种阔叶树种,但其木质部缺乏导管分子。因此,水青树被描述为无导管分子的被子植物[5]。经后续的大量研究,不同的学者提出了不同的观点。水青树是否具有导管分子的问题一直备受关注。一部分学者认为水青树是不存在导管的。李坚等[6]、张萍[7]使用扫描电子显微镜观察了水青树三切面的木质部结构,观察结果表明水青树不具导管。Suzuki 等[8]通过解剖技术在水青树中发现了特殊管胞,他认为这些异常的细胞类似导管,但不属于导管分子。然而,一部分学者认为水青树中的特殊管胞为导管分子。任毅[9]和李红芳等[10]借助环境扫描电子显微镜对8~10 年生水青树的特殊管胞进行观察,表明特殊管胞的纹孔为梯状,且有残留的纹孔膜,他们将这种细胞称为原始导管。陈璐[11]、Carlquist 等[12-13]从导管分子的定义出发,认为水青树存在原始状态的导管分子,且穿孔板上的纹孔膜残留属于细胞进化的特征。Liu 等[14]对水青树特殊管胞的基因进行了探索,发现水青树具有导管分子的相关基因,但并未对特殊管胞的相关形态进行详细描述。
前人对特殊管胞的形态特征已经有了一定了解,也从基因水平对其进行了探究,但依旧对水青树木材中特殊管胞的具体形态、排列规律、细胞壁构造特征及其生理功能等方面没有全面的探索,研究这些有助于明确水青树演化位置,体现保护水青树的重要意义,同时对更好地理解管胞的进化过程及探究树木从针叶材到阔叶材的演化规律有重要指导意义。
1 材料与方法
1.1 试验材料与设备
试验样品取自南京林业大学标本室的34年生水青树径列中心条。试验所用的主要药剂为:无水乙醇(分析纯,南京化学试剂有限公司)、乙酸(冰醋酸,分析纯,南京化学试剂有限公司)、30%过氧化氢(分析纯,国药集团化学试剂有限公司)、二甲苯(分析纯,上海凌峰化学试剂有限公司)、番红(臧红T,阿拉丁试剂(上海)有限公司)、中性树胶(显微镜用,国药集团化学试剂有限公司)。试验所用的主要仪器设备有光学显微镜(Olympus BX51,日本)、体视显微镜(Leica S8AP0,德国)、滑走式切片机(Yamato TU-213,日本)、电子扫描显微镜(Phenom Scientific XL G2,中国)。
1.2 试验方法
1.2.1 制片观察
将木块放在60 ℃的水浴锅中进行水煮软化处理,直至木块完全沉底。采用划走式切片机将软化后的试样进行切片,取厚度为18 μm 的横切面、径切面、弦切面切片,经过1%番红水溶液染色、系列梯度(30%、50%、70%、80%、90%、100%)乙醇脱水、二甲苯∶乙醇=1∶1、二甲苯、中性树脂胶封固,制成永久切片。将三切面的封片置于光学显微镜下,进行显微观察。
1.2.2 管胞形态分析
木块经体视显微镜观察确定特殊管胞所在区域与正常管胞区域,分别切割成10.0 mm×0.5 mm×0.5 mm 的火柴棍大小的木条,放入具5mL 解离液(冰醋酸∶30%过氧化氢=1∶1)的试管中浸泡,并把试管放入水浴锅中70 ℃水浴加热7 h,直至解离液浸泡着的木材样品变白。然后倒掉解离液,并用蒸馏水洗净样品中残留的酸液至样品接近中性,滴10 mL 蒸馏水在试管中,然后振荡试管直至木条都松散成纤维状,再加入1 mL 的1%番红水染液后摇匀。取1滴溶液滴在载玻片上,盖上盖玻片并将其放在光学显微镜下观察。使用Motic Image Plus 2.0 软件对选取的50 个特殊管胞(special tra⁃cheids,记 为ST)和50 个 正 常 管 胞(normal tra⁃cheids,记为NT)的长度,弦向宽度及壁厚进行测量。
1.2.3 扫描电镜观察
取未染色的解离管胞溶液,滴1 滴在载玻片上,使管胞分布均匀后将载玻片放入烘箱烘至绝干,利用导电胶将其移至载物台。用切片机从软化后的试样上切下厚度为60 μm 的三切片,放置烘箱至绝干,后粘到载物台上。将解离管胞和切片喷金,后在扫描电子显微镜下进行显微观察。
1.3 数据处理
用Excel 2016对特殊管胞和正常管胞的长、宽和壁厚等进行统计整理后,求出相应平均值、标准偏差和变异系数。
2 结果与分析
2.1 水青树特殊管胞的分布位置
特殊管胞在水青树的分布如图1 所示。横切面上,特殊管胞紧邻木射线呈径向整齐排列贯穿年轮界限,多为一列,偶见两列。特殊管胞呈矩形,且自早材至晚材的弦向宽度变化不大,径向宽度逐渐变小(如图1:a~b)。弦切面上,特殊管胞两侧紧邻木射线或其中一侧紧邻木射线,另一侧靠近轴向管胞。纵向连接有两种排布形式:纺锤形特殊管胞单个分布;两个或以上的特殊管胞以端接的方式上下单列或多列连接(如图1:c)。
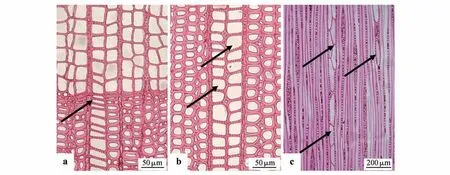
图1 水青树特殊管胞位置分布图a~b 为横切面;a 箭头所指为单列特殊管胞径向排列,且穿过年轮界限;b 箭头所指为2 列特殊管胞径向排列;c 为弦切面,特殊管胞紧邻木射线分布,箭头所指特殊管胞单独分布或上下端接相连Fig.1 Formation position of special tracheids in Tetracentraceae sinense a-b.Transverse sections;a.The single column of special tracheids were arranged in the horizontal direction of the tree,and ran through the growth ring boundary;b.Arrow referred to the radial arrangement of two special tracheids;c.The special tracheids were adjacent to the wood ray in the tan⁃gential section,the special tracheids referred to by the arrow was individually distributed or with several cells terminally
水青树特殊管胞的分布有别于正常管胞,仅一到两列径向分布,这可能会促进木质部的横向输导或对木质部与韧皮部的输导起到连接作用,在纵向上特殊管胞既有单独分布,也存在上下端接的情况下,这说明特殊管胞可能存在一定的进化方向,如多个管胞上下连通加快水分运输等。
2.2 水青树特殊管胞的形态特征
特殊管胞(ST)与正常管胞(NT)的相关尺寸如表1 所示。特殊管胞的平均长度为286.44 μm,平均弦向宽度为55.22 μm,平均壁厚为1.53 μm。水青树中的特殊管胞与正常管胞相比,平均长度约为正常管胞的1/9,平均宽度大15.83%。水青树特殊管胞壁厚变化趋势较正常管胞壁厚较小。
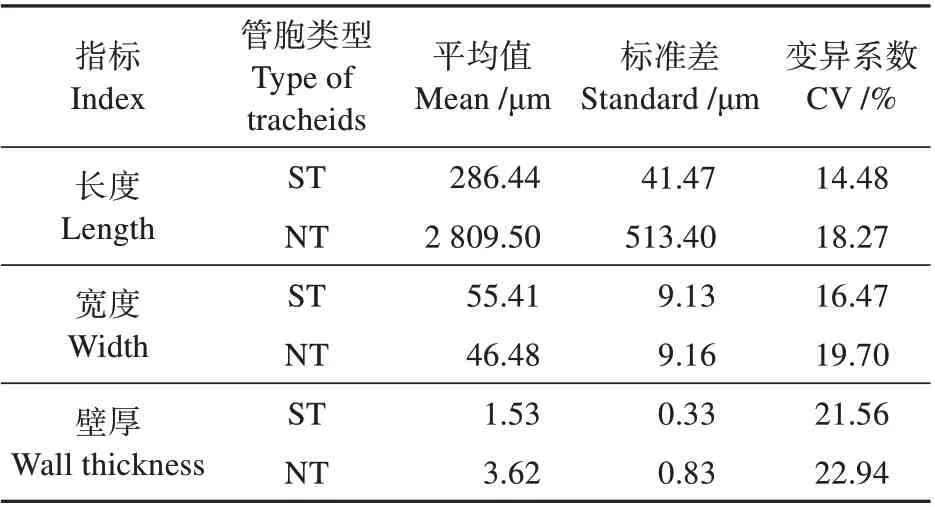
表1 特殊管胞和正常管胞的相关尺寸Table 1 Quantitative anatomical features related to spe‐cial tracheids and normal tracheids
水青树特殊管胞解离后的形态如图2 所示。根据特殊管胞在弦切面的分布,特殊管胞的形态可分为以下3 种:无端壁的纺锤形管胞(如图2:a),有两个倾斜端壁(如图2:b)和只有一个倾斜端壁的细胞(如图2:c)。特殊管胞两端封闭且端壁面光滑无穿孔。特殊管胞侧面壁纹孔丰富。其弦面壁上有大量具缘纹孔,呈梯状、对列、互列分布(如图2:a,c),经扫描电子显微镜放大观察纹孔内残留纹孔膜(如图2:d)。特殊管胞的径面壁上纹孔为具狭缘的单纹孔(如图2:b)。

图2 水青树特殊管胞形态及纹孔特征a~c.水青树不同形态特殊管胞(a.纵向单个排列的纺锤形特殊管胞,两端封闭无穿孔;b.纵向上多个细胞且多列分布的特殊管胞,有两个倾斜端壁;c.纵向上多个单列连接且位于端头的特殊管胞,有一个倾斜端壁);d.特殊管胞弦面壁上的具缘纹孔(箭头所指为纹孔膜残留)Fig.2 Microstructure of special tracheid and characteristics of pits in T.sinense a-c.Were special tracheids in different shape(sa.The spindle-shaped special tracheid arranged in a single longitudinal arrangement,closed at both ends without perforation;b.The special tracheid with multiple cells and multiple columns in longitudinal direction with two sloping end walls;c.The special tracheid was vertically connected by multiple single columns and was located at the end,with one sloping end wall);d.The bordered pits on the wall in tangential section,and the arrow referred to the pit membrane
在形态方面,特殊管胞与正常管胞较为接近,具尖削端头,呈纺锤状或近纺锤状,但长宽尺寸远不同于正常管胞,其长度远小于正常管胞,宽度略大于正常管胞,特殊管胞的径向变化趋势较小。
2.3 水青树特殊管胞侧面壁上纹孔分布
特殊管胞弦面壁上的大量纹孔为特殊管胞间的具缘纹孔,呈对列、互列偶见梯状排列(见图3:a~b)。特殊管胞径面壁上的纹孔为特殊管胞与射线细胞间的纹孔,为具狭缘的单纹孔,呈大圆形至椭圆形(见图3:c)。径面壁上的纹孔成组状分布,每个区域多为2~10个纹孔,呈1~4排横列(见图3:a~b)。特殊管胞的倾斜端壁无纹孔(见图3:b~c),这说明上下排列的特殊管胞间并未连通。特殊管胞与正常管胞间无纹孔(见图3:d)。

图3 水青树特殊管胞侧面壁上的纹孔分布a~b.水青树特殊管胞(a.纺锤形特殊管胞,左侧箭头所指为其径面壁与射线细胞间的具狭缘单纹孔,右侧箭头所指为具缘纹孔,呈互列、对列;b.具一个倾斜端壁的特殊管胞,其中左侧箭头所指为正常管胞和特殊管胞的接触面,无纹孔,右侧箭头所指为倾斜端壁,无纹孔);c.特殊管胞与射线之间的具狭缘的单纹孔,且特殊管胞间上下连接处无纹孔;d.水青树的弦切面(箭头所指表示特殊管胞与正常管胞间无纹孔)Fig.3 Distribution of pits on the wall of the special tracheids in T.sinense a-b.Were special tracheids of T. sinense(a.Showed the spindle-shaped special tracheid,the left arrow was referred to the single pits with narrow edge between the radial wall and the ray cells,the right arrow was referred to the bordered pits,which were arranged in opposite and alternate.;b.Showed a special tracheid with an inclined end wall,the left arrow referred to the contact surface without pits between normal tracheids and special tracheids,the right arrow referred to the inclined end wall without pits);c.Showed the single pits with narrow edge between a special tracheid and a ray,there were no pits at the junctions of the special tracheids;d.showed the tangential section(This were no pits indicated by the arrow between the special tracheids and the normal tracheids)
特殊管胞之间的纹孔位于弦面壁,正常管胞间的纹孔多位于径面壁,这导致了两种管胞的不同输导方向,即特殊管胞主要为径向输导,特殊管胞与木射线之间也存在纹孔,说明特殊管胞与木射线间存在一定输导关系。
3 讨论
3.1 管胞向导管分子进化的性状表现
导管分子由管胞逐渐演化而来[15]。管胞主要起到输导水分和机械支撑作用[16]。管胞呈纺锤形,无端壁,不存在穿孔,上下管胞间搭接,侧壁上管胞间搭接处常具纹孔,水分和无机物均需穿过纹孔膜实现细胞间的物质运输。随着针叶材向阔叶材的进化,相关基因表达的性状已经能产生特定的酶等物质,将初生壁纹孔膜等部分溶解掉,上下细胞间演化为端接,无纹孔膜阻碍,使得纵向连接的导管分子端壁上形成通透的穿孔,进化为专司输导功能的导管,这就使得水分和无机物的运输速度和强度明显提高[17]。所以,导管和管胞在形态上的最大区别为:端壁有无穿孔,一般认为穿孔与纹孔的区别主要是纹孔膜的消失与否[15]。
管胞—原始导管—进化导管的演化过程主要有以下几个特征:(1)细胞形态。细胞长度由长向短进化,横切面形态由四边形向多边形、卵圆形、圆形进化;(2)细胞间连接处由纹孔向穿孔(梯状穿孔-单穿孔)进化[18];(3)细胞端处由较长(两端尖削)向较短进化[15];(4)管间纹孔式。其进化速度较穿孔板快,依次为梯状、梯状对列、对列、互列[19]。水青树特殊管胞较正常管胞形态有较大差别,处于由管胞向导管进化过程中,其分布规律也明显区别于正常管胞,这表明了特殊管胞是由管胞向导管分子演化过程中的一种中间类型细胞。
3.2 水青树特殊管胞与一般导管的比较
将一般阔叶材的导管与笔者所观察到的特殊管胞进行比较,可以发现两者有一定相似之处:在排列分布上,阔叶材导管存在径列复管孔或径向排列分布[20],这与特殊管胞径向呈串排列较类似;在纹孔分布上,径向排列的导管间的弦面壁上存在大量管间纹孔,且导管与木纤维间几无纹孔[16],与射线间具较多纹孔,这与径向排列的特殊管胞弦面壁上具大量纹孔且与射线间具较多纹孔完全一致。两者的主要不同之处在于:导管分子纵向连通起到运输水分的作用,而特殊管胞两端封闭无法实现在纵向上的水分输导;从横切面上看导管分子呈径向排列、径列复管孔、散生或成团分布,而特殊管胞仅呈长串状径向分布。特殊管胞的相关形态处于进化过程中的阶段,其目的可能是更好地适应环境[21]。
近年来,相关研究人员发现了一些处于进化中的导管分子。黄玉源等[17,22-23]先后在银杏类植物、松柏纲、红豆杉纲和苏铁类植物中发现了导管,其中银杏(Ginkgo biloba)的茎组织中具导管,其端壁一部分呈倾斜状,部分呈近水平状,其端壁的穿孔明显可见且大小不一呈梯状或网状;在水杉(Metasequoia glyptostroboides)、南洋杉(Araucar⁃ia cunninghamii)等植物中发现的导管分子,其穿孔尺寸较大且数量较多,部分存在与侧壁呈水平状态的穿孔板,属于被子植物发达导管所具备的特征。李臻等[24]在木贼属的根状茎和气生茎的木质部中发现导管分子,次生壁加厚呈螺纹或梯状。这可能是树木长时间处于严峻环境如干旱缺水、土地贫瘠等,导致管胞出现变化[25]。朱俊义[26]对花楸(Sorbus pohuashanensis)的导管分子观察,重演了系统发育过程中导管网状穿孔板演化成单穿孔板的过程。上述细胞存在较大尺寸的穿孔,且形态远不同于水青树特殊管胞,较水青树特殊管胞更接近传统意义上的导管。
3.3 水青树特殊管胞在功能方面的意义
在针叶材向阔叶材进化过程中,特殊管胞的出现对提高树木的输导功能具有重要意义。作为高大阔叶乔木,水青树为满足自身快速生长和叶片大量的水分蒸腾需求,其木质部需要有强大的输导系统。在缺乏成熟导管的条件下,特殊管胞的存在至关重要。虽然纵向连接的特殊管胞并未连通,但其紧邻木射线径向呈串连接,扩大了木质部的径向输导渠道,且径向相连的特殊管胞弦面壁上存在大量具缘纹孔,拓宽了树木径向与轴向的运输路径。同时特殊管胞在树木的生长发育中也起到一定作用。每年形成层活跃初期水青树难以将水分从轴向管胞传到形成层细胞[8],但特殊管胞可保证水分在径向上通过弦面壁上的纹孔快速流动。正常管胞的弦面壁上纹孔较少,因而与形成层间水分传输受限,水分可以通过射线细胞从轴向管胞转移到特殊管胞而后进入形成层细胞或直接从射线细胞转移到形成层细胞。特殊管胞与射线细胞有直接联系这一事实可能具有一定的生理意义,它们可能有助于射线细胞中糖分的快速运输[8]。
4 结论
水青树的特殊管胞分布位置和形态特征存在一定规律。在分布位置方面,水青树特殊管胞沿径向连续排列并贯穿年轮界限,在纵向上紧靠木射线单个排列或多个细胞1~2 列上下端接;在形态特征方面,水青树特殊管胞接近纺锤形或有1~2个倾斜端壁,平均长度远小于正常管胞,平均宽度接近正常管胞;其特殊管胞两端封闭,不存在穿孔;在纹孔的分布方面,水青树特殊管胞弦面壁上的纹孔较为密集具纹孔膜,且分布与一般导管高度相似,但也不完全一致;在输导功能方面,水青树特殊管胞已经具有与一般管胞不同的输导功能。虽然水青树的特殊管胞已经被证实具导管分子相关基因,但其相关性状并未完全表达。综上,水青树的特殊管胞并不是真正意义上的导管。水青树正处于由针叶材向阔叶材进化阶段中的某一状态,而其特殊管胞的分布位置及形态特征标志着其处于管胞分子向导管分子演化的一定阶段。
