基于谷子种质资源表型性状构建骨干种质库
2022-11-04臧贺藏李国强解慧芳王素英刘金荣郑国清
宋 慧 臧贺藏 李国强 解慧芳 邢 璐 李 龙 王素英 刘金荣* 郑国清*
(1.河南省农业科学院 农业经济与信息研究所/河南省智慧农业工程技术研究中心,郑州 450002;2.安阳市农业科学院 谷子研究所/河南省谷子育种工程技术研究中心,河南 安阳 455000)
谷子起源于中国,早在公元前5000—6000年二倍体谷子[Setariaitalica(L.)Beauv.]栽培种已在黄河流域广泛种植[1],谷子籽粒营养丰富、耐旱性强,是北方旱作地区重要的特色杂粮作物[2]。种质资源是新品种选育和遗传研究的基础,我国拥有最丰富的谷子种质资源,占世界总资源的70%左右[3],在谷子育种领域两次跨越式发展都与种质资源的定向改良有关,具有里程碑意义的谷子常规品种‘豫谷1号’引领谷子由农家种向育种家品种的跨越,改变了谷子作为低产作物的历史,其亲本是由日本引进的 ‘日本60日’和‘土龙’;杂交谷子育种带动谷子由中高产作物迈向高产作物行列,其中,不育系‘A2’的亲本来源于澳大利亚。当前,在丰富的种质资源基础上提高资源利用效率和拓宽种质遗传基础是培育谷子突破性新品种的核心,也是实现谷子种业创新的关键问题之一。
构建骨干种质能有效解决庞大种质资源群体研究与高效利用之间的矛盾,国内外学者分别基于表型性状和分子标记构建了花生[4]、水稻[5]、大豆[6]、小麦[7]和芝麻[8]等作物的骨干种质,以最小的资源量覆盖最全的资源群体遗传多样性,既保证了种质资源的遗传多样性,又减少了资源样本的数量,有利于资源研究。谷子种质具有丰富的遗传多样性,王海岗等[9-10]对不同来源的878份谷子核心种质进行了综合鉴定和遗传多样性评估,利用SSR标记分析了山西省谷子资源的群体遗传结构多样性。杨慧卿等[11]选用了来自国外及国内的68份分蘖型谷子进行遗传多样性分析,其数量性状遗传变异丰富。丁银灯等[12]对国内外124份谷子种质资源进行表型及SSR分析发现谷子单株生物产量、单穗重的数量性状变异系数较大,遗传多样性指数较高。贾冠清等[13]以SSR标记为基础,构建了核心种质,绘制了SNP标记的谷子单倍型图谱,发掘出控制多个谷子主要农艺性状和植物学性状的QTL位点,形成了一个基于关联分析的谷子功能基因发掘平台,为后续分子标记辅助育种提供支撑。针对当地育种现状重新构建和利用资源骨干种质是拓宽谷子育种遗传基础、解决遗传变异冗余和样本利用效率低等问题的重要途经。目前,以地方农家品种为主构建骨干种质的研究鲜见报道。本研究以1 237份谷子种质资源(包括野生种、农家种、育成品种和国外引进品种)为材料,以19个数量和质量表型性状数据为基础,构建谷子种质资源的骨干种质,利用极差、表型保留比例、表型相关性等对谷子骨干种质的代表性、主成分分析、直方图进行评价和确认,旨在高效发掘和利用以地方农家品种为主的谷子种质资源,以期为谷子种质资源的高效创新利用提供技术支撑。
1 材料与方法
1.1 试验材料及地点
由中国农业科学院作物所提供1 237 份地理来源明确的种质资源,包括引进品种、育成品种、地方品种、野生品种。试验于2020年在河南省北关区柏庄镇安阳市农业科学院试验基地进行,于5月20日播种,10月3日收获。
1.2 试验设计
小区面积2 m2,种植密度60万株/hm2,行长1 m,行距40 cm,2行区种植。田间管理参照全国谷子联合鉴定试验的方法管理。
1.3 调查内容及方法
根据谷子种质资源描述规范和数据标准对生育期、主茎节数、株高、穗下节间长度、主穗长度、叶面积、草重、刺毛颜色、幼苗叶色、幼苗叶姿、穗形、穗姿11个田间农艺性状和主穗长度、单株穗重、穗粗、主穗码数、主茎单穗重、主茎单穗粒重、单株粒重、千粒重8个产量性状,共19个谷子表型性状进行采集。质量性状根据其表型进行赋值(表1),数量性状数据根据0.5个标准差为间距进行标准化,分为10级(1级≤X-2δ,10级>X+2δ,中间每级间差0.5δ,X为性状平均值,δ为标准差)。

表1 谷子质量性状赋值Table 1 Quantified values of the qualitative traits of foxtail millet
1.4 分组及取样比例
根据品种来源分为引进品种、农家品种、野生品种和育成品种4个类型,按植物学性状中穗形分成7组。分组取样量指全资源份数的平方根值占小组平方根之和的比例(平方根取样法)。
式中:Ni代表第i组的取样份数,ni代表第i组的品种数,nj代表第j组的品种数,m代表总分组数,N代表总资源份数。
1.5 聚类分析
采用SPSS 22.0软件以类平均数法(UPGMA)进行表型性状的系统聚类分析,类内采用Microsoft Excel中的Rand between函数随机取样。
1.6 检测骨干种质的代表性
利用t测验、F测验分别评价全部种质和骨干种质表型性状的均值、方差、变异系数及Shannon-Weaver指数是否存在显著差异。
变异系数(CV)=(标准偏差SD/平均值Mean)×100%,代表全部种质和骨干种质的离散程度。
(1)
式中:H′为Shannon-Weaver 多样性指数,Pi为某性状第i级的分布频率,n为总分级数。
表型保留比例(the ratio of phenotypic retention,RPR)用于评价骨干种质中覆盖资源的变异程度。
(2)
式中:Pio为全部种质中某性状的表型变异数,Pi为骨干种质中该性状的表型变异数,n为表型性状总数。
1.7 评价骨干种质
利用主成分分析和直方图比较全部种质与骨干种质基于主成分的样品分布图及表型性状的分布频率,对构建的骨干种质的有效性进行确认。
2 结果与分析
2.1 谷子骨干种质的筛选构建
由表2可知,按照不同穗形将谷子全部种质分7个小组,UPGMA法聚类分析后随机取样,每组取样量为1~17份,得到113份种质资源,经初评后,将野生种质中增补5份入选,共确定出118份骨干种质,占全部种质资源的9.9%。此骨干种质中按地理生态区包括国外品种20份(17.09%)、农家种56份(47.86%)、野生种5份(4.28%)、育成品种36份(30.77%),代表了谷子资源的地理来源(P=0.579),构建的骨干种质与全部种质之间差异水平不显著,即骨干种质能够代表全部种质。按不同穗形可分为圆锥型39份(33.32%)、纺锤型23份(19.66%)、圆筒型21份(17.95%)、棍棒型16份(13.68%)、鸭嘴型4份(3.42%)、猫爪型8份(6.84%)、佛手型6份(5.13%),方差分析表明,骨干种质穗形代表了全部种质的穗形组成(P=0.637),差异水平不显著,骨干种质能代表全部种质的类型。
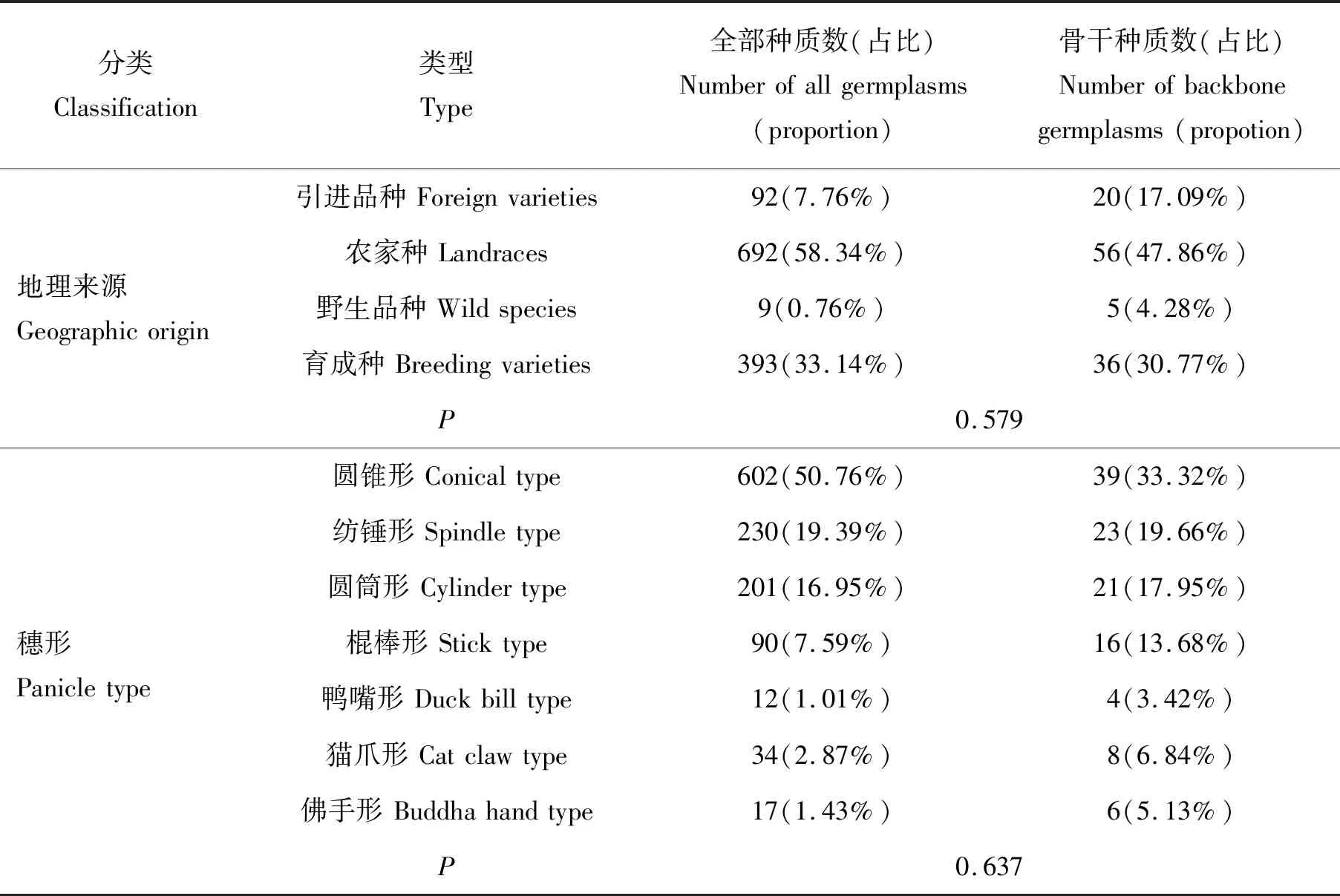
表2 骨干种质和全部种质的植物学类型组成和地理来源的显著性测验Table 2 Significance tests for botanical type composition and geographical origin of backbone germplasm and all germplasm
2.2 谷子全部种质与骨干种质的平均值、方差和极差比较评价
由表3可知,谷子骨干种质的19个表型性状的均值皆大于全部种质,且骨干种质中穗形、主穗码数和千粒重与全部种质之间差异不显著。F测验表明,骨干种质大部分性状的方差大于全部种质,说明构建的谷子骨干种质遗传冗余度明显减小,变异率更高,全部种质与骨干种质的差异未达到显著水平,说明构建的骨干种质能代表全部种质。
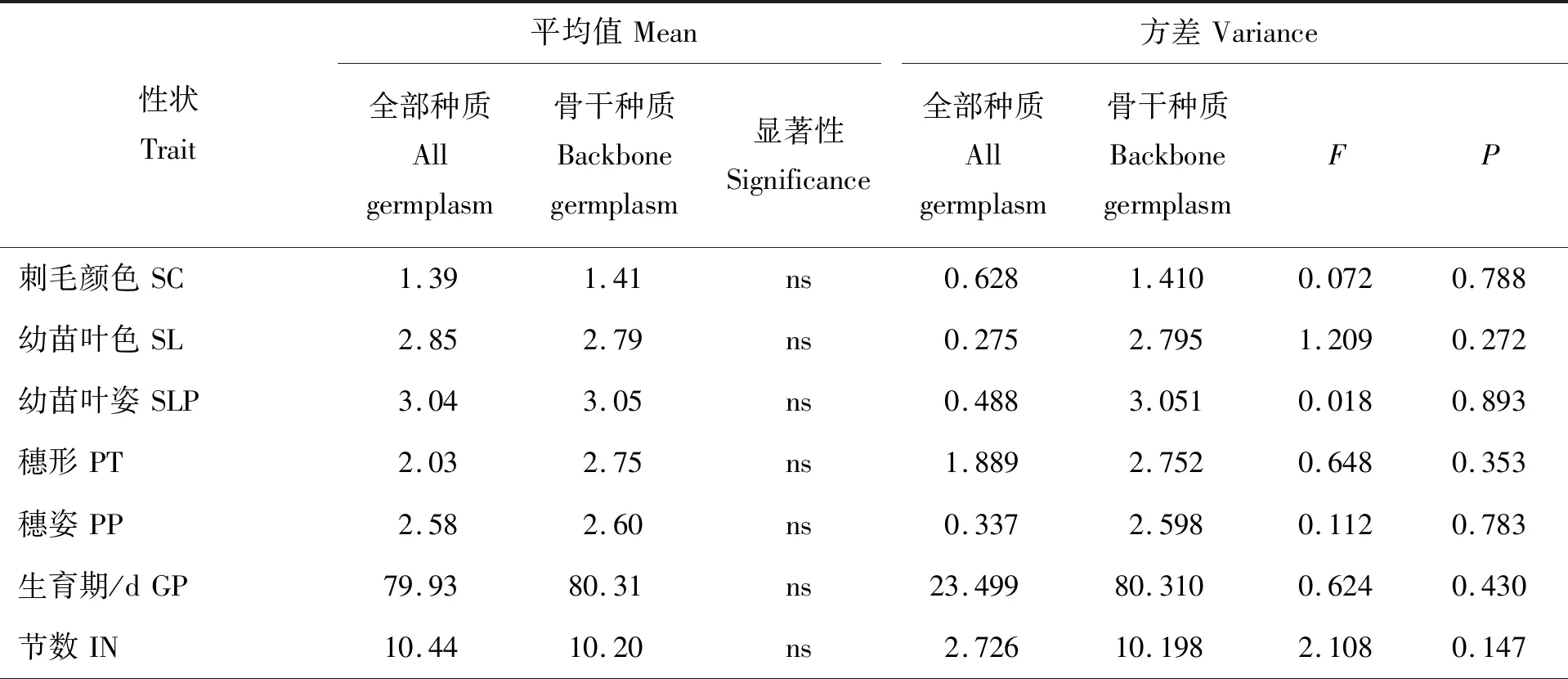
表3 全部种质与骨干种质19个表型性状平均值和方差的比较Table 3 Comparison of means and variances for ninteen traits in all germplasm and backbone germplasm
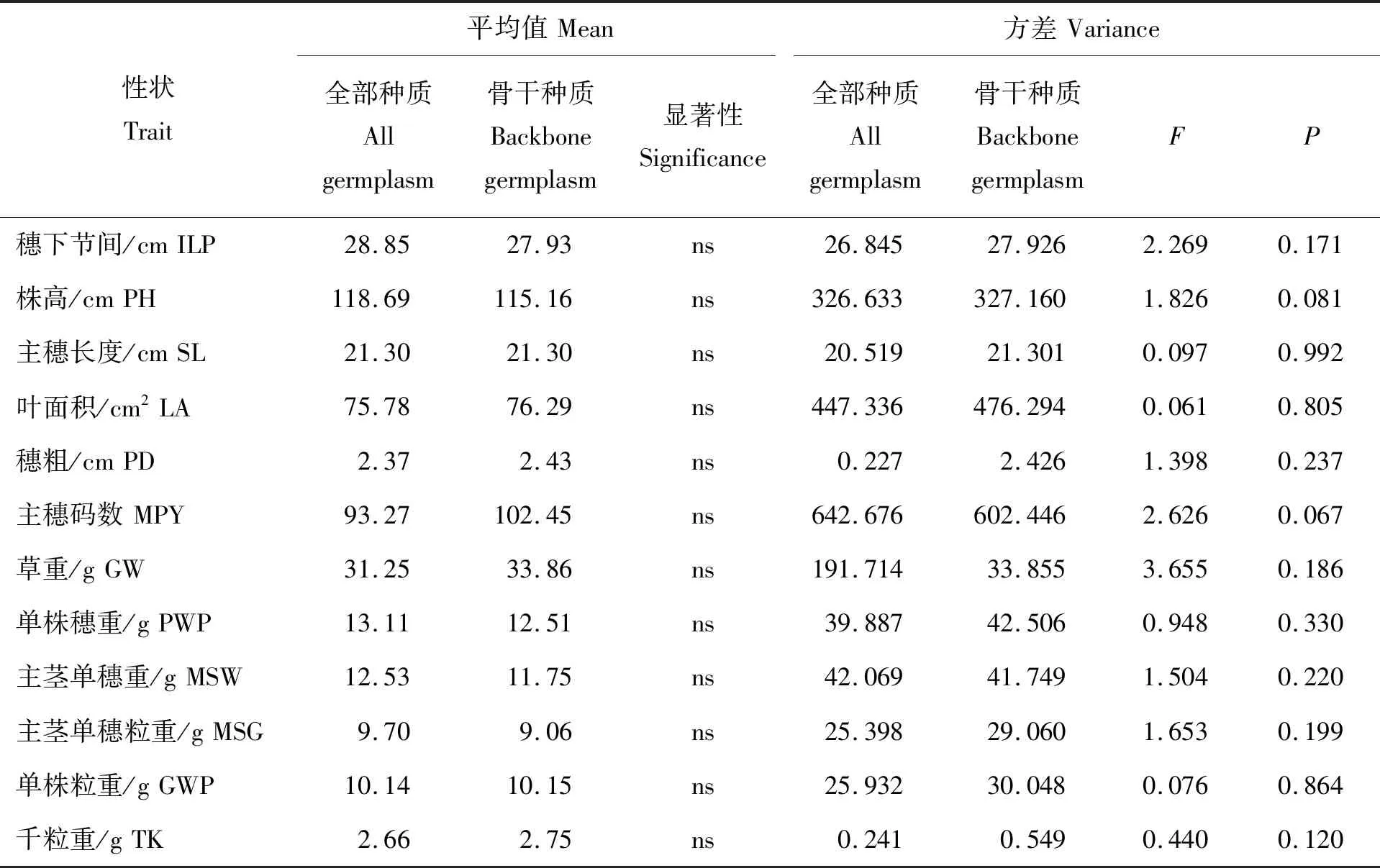
表3(续)
2.3 极差、Shannon-Weaver多样性指数及变异系数的比较评价
由表4可知,谷子全部种质质量性状的极差变异范围全部保留在骨干种质中;数量性状变异范围除单穗重(67.77%)、穗粗(77.09%)、千粒重(74.68%)外,其余性状变异范围84.66%~99.96%保留在了骨干种质中。骨干种质平均H′和CV分别为1.76±0.154和37.52±5.765,H′显著高于全部种质(P=0.004),变异系数差异不显著(P=0.356)。
2.4 分布频率及表型保留比例的比较评价
由表5可知,骨干种质覆盖了全部种质的表型分级。卡方测验结果表明,表型差异均未达显著水平,说明骨干种质代表了全部种质的变异,穗形的表型保留比例较大,可能是由于补充了一些特殊种质和极端材料所致;其余性状的表型保留比例均比较合适,表明骨干种质保留了全部种质丰富的变异,且丰度更高。
2.5 谷子骨干种质的确认
由图1可知,谷子全部种质中存在大量样本重叠和遗传背景相似性,群体的遗传冗余程度高,尤其坐标轴原点(0,0)交叉处样本密度大,重叠程度高,经筛选后谷子骨干种质样品主成分分布图保留了谷子全部种质的外部轮廓特征,降低了样本之间交叉重叠(图1(b)),样本的分布和密度明显减低,且较多外围的个体入选到骨干种质中,构建的谷子骨干种质既去除了全部种质的大部分遗传冗余,又确保了骨干种质的代表性。因此,所构建的骨干种质能够覆盖全部种质的遗传多样性和群体结构,表明骨干种质的构建是有效的。
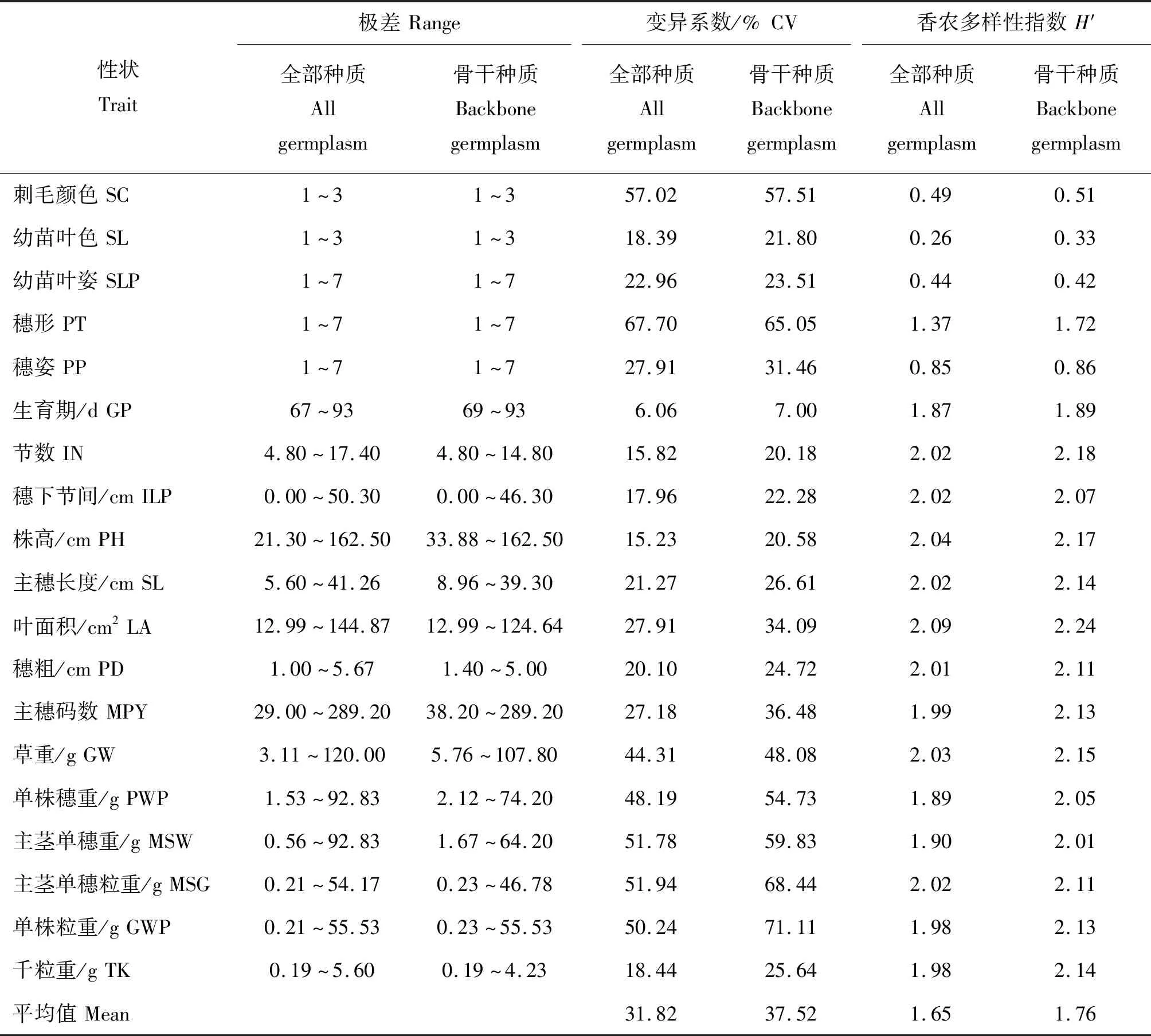
表4 全部种质与骨干种质极差、变异系数和遗传多样性指数的比较Table 4 Comparison of range,coefficient of variation and genetic diversity index of all germplasm and backbone germplasm
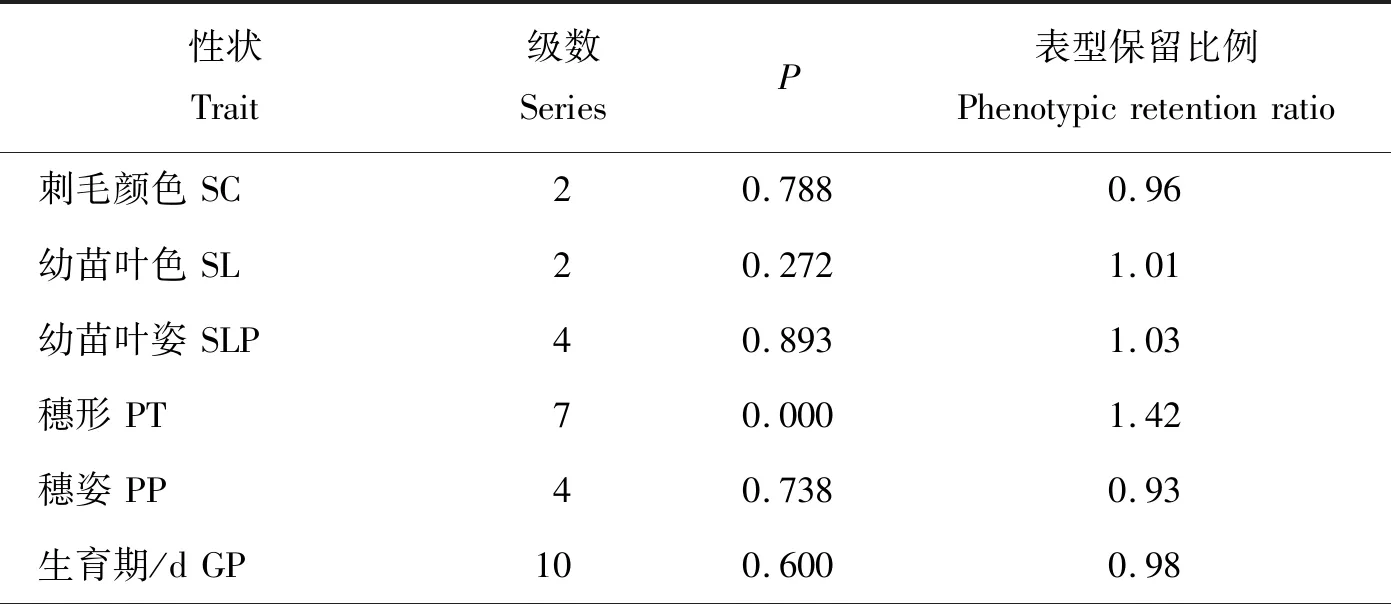
表5 全部种质与骨干种质表型性状的分布频率和表型保留比例的比较Table 5 Comparison of the distribution frequency and phenotypic retention ratio of traits between all germplasm and backbone germplasm
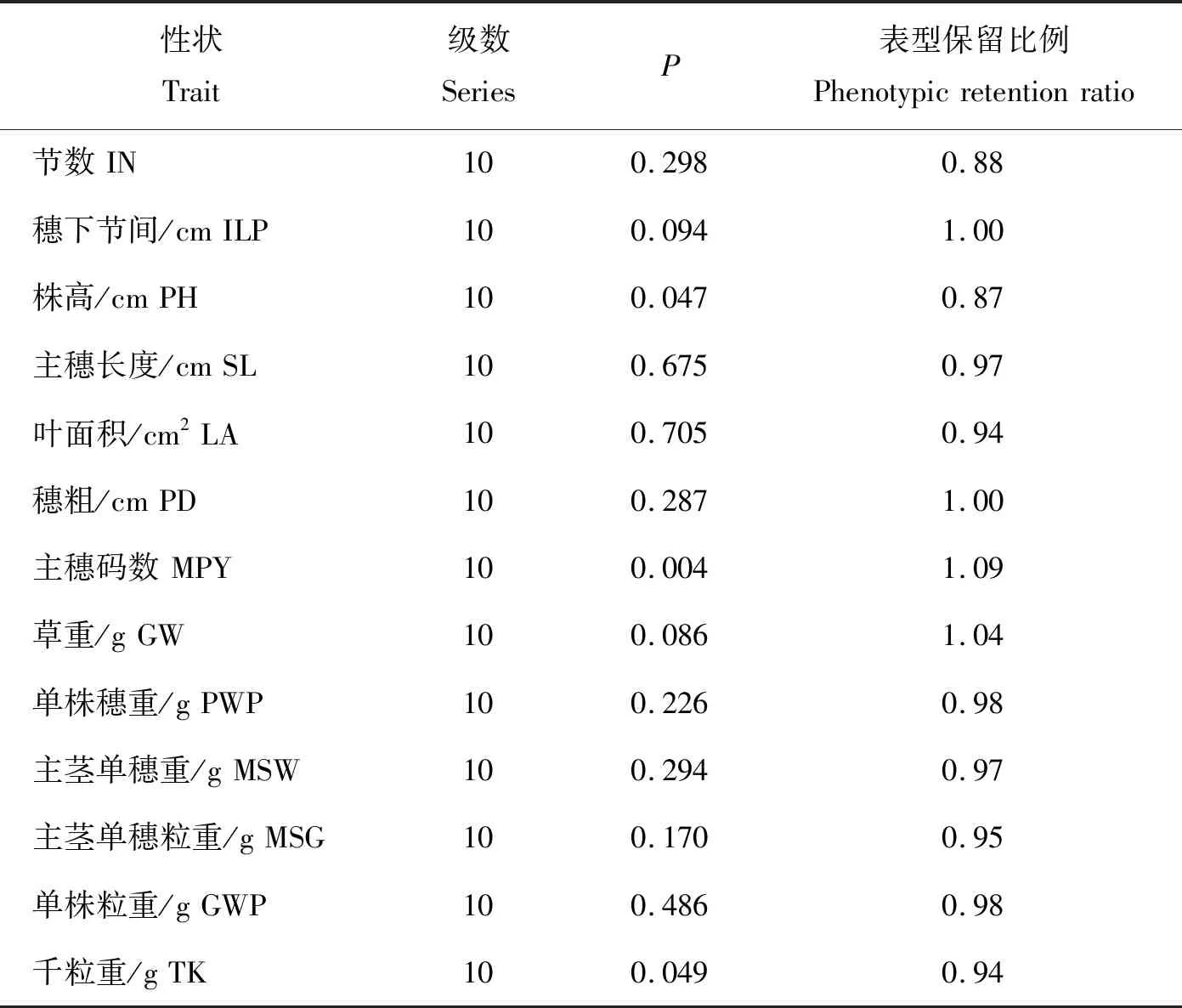
表5(续)
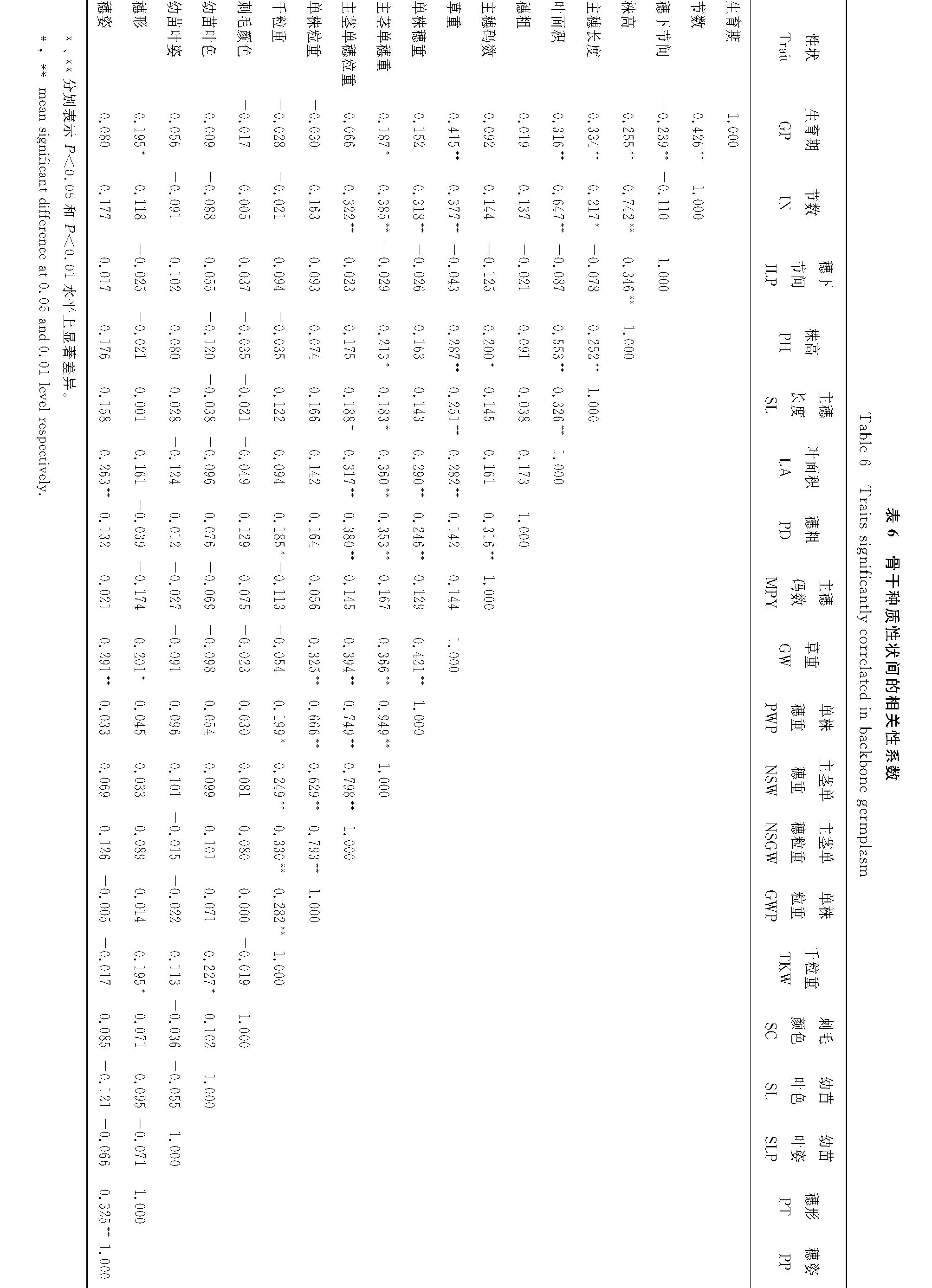
图1 全部(a)与骨干种质(b)的样品主成分分布图Fig.1 Principal component plots for the all (a) and backbone germplasm (b)
由图2可知,在8个产量性状表型数据中,全部种质和骨干种质的主穗长度、主茎单穗重、主茎单穗粒重、穗粗、千粒重、单株穗重、单株粒重、主穗码数分布均呈正态分布,且构建的谷子骨干种质与全部种质分布趋势一致。因此,构建的骨干种质很好地保留了全部种质的群体结构和遗传多样性,确保了骨干种质的有效性。
由图3可知,谷子田间农艺性状包括幼苗叶色、叶面积、穗姿、穗形、穗下节间长度、生育期、刺毛颜色、草重、株高、幼苗叶姿、主茎节数在全部种质和骨干种质中都表现出广泛的变异,且大部分变异的分布频率较为一致,均基本符合正态分布,因此,构建的118份谷子骨干种质能够有效覆盖全部种质的遗传多样性和全部群体结构。
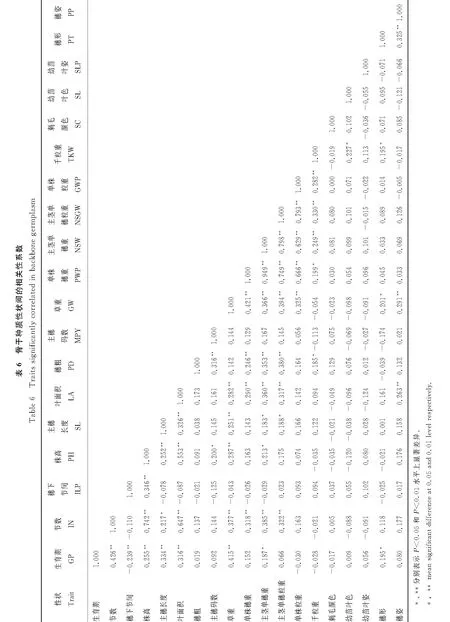
2.6 不同表型性状间相关分析
由表6和表7可知,构建的谷子骨干种质与全部种质表型性状之间相关性一致。全部种质与骨干种质在叶面积、草重、单株穗重、主茎单穗重、主茎单穗粒重、单株粒重性状都表现为极显著正相关,均可作为高产育种的决策指标。

(a)、(c)、(e)、(g)、(i)、(k)、(m)、(o)分别为谷子全部种质的主穗长、主茎单穗重、主茎单穗粒重、穗粗、千粒重、单株穗重、单株粒重和主穗码数;(b)、(d)、(f)、(h)、(j)、(l)、(n)、(p)分别为谷子骨干种质的主穗长、主茎单穗重、主茎单穗粒重、穗粗、千粒重、单株穗重、单株粒重和主穗码数。(a),(c),(e),(g),(i),(k),(m),(o) are main spike length,spike weight of main stem,grain weight per spike of main stem,spike diameter,1 000-grain weight,spike weight per plant,grain weight per plant,yard number of main spike in all resources of foxtail millet,respectively.(b),(d),(f),(h),(j),(l),(n),(p) are main spike length,spike weight of main stem,grain weight per spike of main stem,spike diameter,1 000-grain weight,spike weight per plant,grain weight per plant,yard number of main spike in backbone resources of foxtail millet,respectively.图2 全部资源与骨干种质的8个谷子产量性状表型频率直方图Fig.2 Phenotypic frequency histograms of 8 millet yield traits in all resources and backbone germplasm
3 讨 论
3.1 构建谷子骨干种质对取样方法的选择
确定合适的取样方法是构建骨干种质的基础,常见的取样方法有优先取样[14-15]、偏离度取样[16-17]和随机取样[18-19]。优先取样能保留原种质的表型性状极值及遗传变异结构;偏离度取样可以最大范围覆盖原种质资源的遗传变异;随机取样策略则维持原种质的遗传多样性形式[20]。刘娟等[21]使用随机取样和位点优先取样策略构建了新疆野杏的骨干种质资源,比较发现位点优先取样的资源多态性位点数小于随机取样法。郑福顺等[22]分别选用随机取样法、优先取样法、偏离度取样法进行取样,利用UPGMA进行聚类对480份番茄种质资源构建了宁夏回族自治区番茄骨干种质资源。刘方等[23]采用UPGMA聚类分析对不同地域的103份亚洲棉进行了表型遗传分析,将其聚类为8个组群。李洪果等[24]利用多次聚类+位点优先取样法+马氏距离+最短距离法的取样最佳策略,从306份杜仲雄性种质中构建了包含33份材料的杜仲雄性资源的骨干种质。潘英华等[25]先根据地理分布将4 137份普通野生水稻进行分组,再结合SSR标记分析,进行多次UPGMA聚类,构建了包含351份种质的广西普通野生水稻资源的骨干种质。本研究选取的1 237份谷子种质资源是在已筛选916份核心种质[13]的基础上,又从全国补充321份种质资源,扩充资源样本群体的遗传多样性,然后对谷子资源根据地理来源和穗形分成7个小组,每组种质从9到692份不等。每组的取样量为1~17份;UPGMA法聚类分析后随机取样,得到113份种质,初步评价后补充极值材料和特殊性状种质5份,提取118份(9.9%)的谷子种质资源构建骨干种质。本研究采用均值、方差、极差、变异系数、Shannon-Weaver多样性指数、分布频率、表型保留比例等方法对构建的骨干种质进行评价,构建的骨干种质与全部种质间的差异不显著,最大限度的覆盖了谷子资源的遗传变异,进一步验证了聚类分析和随机取样谷子骨干种质的构建的有效性。
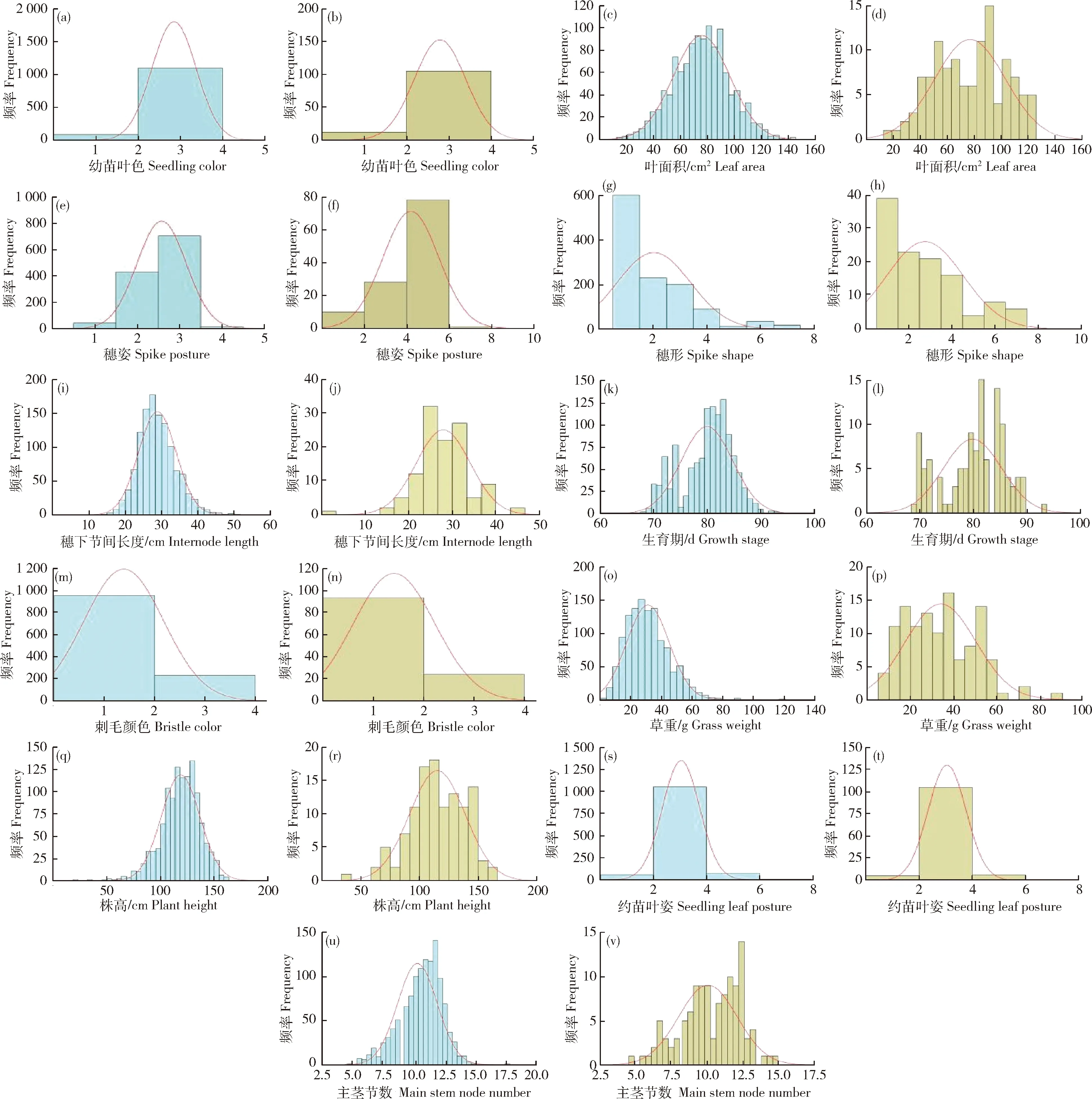
(a)、(c)、(e)、(g)、(i)、(k)、(m)、(o)、(q)、(s)、(u)分别为谷子全部种质的幼苗叶色、叶面积、穗姿、穗形、穗下节间长、生育期、刺毛颜色、草重、株高、幼苗叶姿、主茎节数;(b)、(d)、(f)、(h)、(j)、(l)、(n)、(p)、(r)、(t)、(v)分别为谷子骨干种质幼苗叶色s、叶面积、穗姿、穗形、穗下节间长、生育期、刺毛颜色、草重、株高、幼苗叶姿、主茎节数。(a),(c),(e),(g),(i),(k),(m),(o),(q),(s),(u) seedling color,leaf area,spike posture,spike shape,internode length,growth stage,bristle color,grass weigh,plant height,seedling leaf posture,main stem node number in all resources of foxtail millet,respectively;(b),(d),(f),(h),(j),(l),(n),(p),(r),(t),(v) seedling color,leaf area,spike posture,spike shape,internode length,growth stage,bristle color,grass weigh,plant height,seedling leaf posture,main stem node number in all backbone of foxtail millet,respectively.图3 全部种质与骨干种质的11个谷子农艺性状表型频率直方图Fig.3 Phenotypic frequency histogram of 11 foxtail millet agronomic traits in all resources and backbone germplasm

3.2 评价及确认谷子骨干种质
遗传多样性指数、表型方差、表型分布频率、变异系数、表型保留比率等是衡量骨干种质的重要参考[26]。Hu等[27]采用UPGMA聚类+马氏距离分析,通过t检验、同质性检验对168份棉花种质资源和21份骨干种质的11个表型性状进行比较,证明了构建的骨干种质的可靠性。本研究通过t检验、F测验、对骨干种质与全部种质进行比较分析,t测验表明,骨干种质H′显著高于核心种质(P=0.004),变异系数在骨干种质和全部种质中差异不显著(P=0.356),说明骨干种质的样本足够大,且有效去除了谷子种质资源中的冗余,保留了骨干种质的遗传多样性,变异均匀度显著提高。Shannon-Weaver多样性指数和变异系数表明全部种质和骨干种质的19个表型性状的H′和变异系数是非常相似的,大部分性状的变异范围均保留在骨干种质中。卡方测验结果表明,骨干种质的19个表型性状与全部种质之间无显著差异,表明骨干种质保留了全部种质丰富的变异,且丰度更高。另外,二者在穗形性状组成(P=0.637)和地理来源(P=0.579)上也有很好的一致性。利用主成分分析和直方图对骨干种质进一步确认,骨干种质的遗传冗余较小,离散程度和分布特点与全部种质一致;骨干种质的性状变异广泛,分布频率与全部种质资源较符合,进一步验证了骨干种质良好的代表性和有效性。表型性状相关性分析表明,产量性状表型中叶面积、草重、单株穗重、主茎单穗重、主茎单穗粒重、单株粒重等性状在骨干种质和全部种质呈极显著正相关,骨干种质和全部种质的叶面积与草重(r=0.282**、r=0.349**)、单株穗重(r=0.290**、r=0.289**)、单株穗粒重(r=0.339**、r=0.317**)均呈极显著正相关,在育种过程中,优先选择时叶面积和生物量大的单株,5~6代稳定后测量产量。优先筛选与其产量极显著正相关的易测量田间性状,关联出复杂数量性状,从而便捷的实现育种目标。
4 结 论
基于地理来源和穗形分组,平方根法确定取样量,组内按表型数据进行UPGMA聚类分析,类内随机取样,构建了中国谷子地方品种的118份骨干种质,占全部资源的9.9%,其性状的均值、方差、极差、变异系数、Shannon-Weaver多样性指数、表型保留比例等与全部种质的差异不显著,地理生态区和穗形分布一致,保持了中国谷子种质资源丰富的表型分布频率和表型相关性。主成分分析和直方图结果也进一步确认了骨干种质的遗传多样性和群体结构与全部种质的一致。因此,本研究构建的骨干种质具有很好的代表性。
