合浦珠母贝PfCLEC17A 基因的克隆及表达分析
2022-11-03郑玉斯白丽蓉喻达辉
郑玉斯,王 培,郭 颖,白丽蓉,喻达辉,赵 森
(北部湾大学 广西北部湾海洋生物多样性养护重点实验室,广西 钦州 535011)
海水珍珠养殖产业是北部湾海域的特色经济支柱产业之一,合浦珠母贝(Pinctadafucata)作为培育海水珍珠的主要贝种,近年来由于养殖环境恶化和病害增多等因素,使得合浦珠母贝在人工养殖过程中出现大量死亡现象,造成珍珠的产量和质量下降,这一情况严重制约了合浦珠母贝养殖产业的发展[1]。养殖环境的恶化会造成贝类免疫力降低,同时解决病害问题也需要激活合浦珠母贝自身的免疫功能来发挥作用,所以对合浦珠母贝机体的免疫机制研究就显得特别重要。贝类属于无脊椎动物中的软体动物门,与脊椎动物不同,由于其缺乏适应性免疫,主要依靠先天免疫来防御各种病原体[2]。其中,识别抗原分子是机体先天免疫发生起始的关键,之后才能将识别后的信号传递到下游通路从而激活免疫应答[3]。这种识别是由一种或多种病原谱模式识别受体(Pattern recognition receptors,PRRs)来完成,它们可以识别并结合那些微生物表面保守而在宿主中不存在的特定的病原相关分子模式(Pathogen associated molecular patterns,PAMPs)[4],例如细菌脂多糖(LPS)、肽聚糖(PGN)、脂蛋白、鞭毛蛋白和核酸结构等,即PRRs与PAMPs结合后能够检测非自身分子并触发机体免疫防御[5]。到目前为止,已经有不少学者对PRRs家族进行了鉴别和研究。其中,C型凝集素是PRRs家族中的研究热点,它在软体动物先天免疫反应中参与非自身分子识别过程[6-9]。
C型凝集素是一种钙离子依赖的糖结合蛋白,参与多种免疫相关和其他生理功能。它通过与靶细胞中的糖基化分子表面的多糖进行糖特异性结合,在病原体识别和细胞间的相互作用中发挥重要作用[10]。C型凝集素有一个典型的结构域,命名为C型凝集素结构域(CTLD)或碳水化合物识别结构域(Carbonhydrate-recognition domain,CRD),它与微生物中存在的碳水化合物有很强的亲和力[11]。通过这种特异性结合,C型凝集素能够介导多种重要的细胞过程,包括白细胞黏附和快速刺激防御机制对抗有害微生物等[12]。到目前为止,已经在软体动物中鉴定了多种C型凝集素基因[13-15]。这些基因参与各种生物免疫过程,包括非自我识别、微生物凝集、吞噬作用和包裹作用的诱导以及抗菌特性[13]。例如长牡蛎(Crassostreagigas)的CgCLec-2在免疫识别中作为模式识别受体发挥作用,在病原消除后作为调节因子,并在补体系统的激活中发挥潜在作用[15]。此外,在栉孔扇贝(Chlamysfarreri)中鉴别了5个C型凝集素基因(CfLec-1~CfLec-5),在海湾扇贝(Argopectenirradians)上发现了7个C型凝集素基因(AiCTL-1~AiCTL-7),其中大部分凝集素只对某些特定的革兰氏阳性或革兰氏阴性菌起作用[16]。最近,在菲律宾蛤仔(Ruditapesphilippinarum)中也鉴定出2种C型凝集素(Vpclec-3和Vpclec-4),它们作为模式识别受体具有明显的识别谱,并可能参与菲律宾蛤仔对弧菌的天然免疫应答[17]。以上研究结果显示,双壳类动物体内的C型凝集素与病原体表面的碳水化合物有较强的亲和力,在各种微生物感染的早期阶段发挥着重要作用[18-19]。目前,在合浦珠母贝中已报道了PoLEC1[20]、PoLEC2[21]和PmLec-8[22]等C型凝集素的相关研究。虽然对C型凝集素基因家族中的部分成员进行了相关研究,由于海洋环境的复杂性,海洋生物在面对不同的病原微生物时的识别机制可能有所不同。因此,有必要进一步鉴定C型凝集素基因家族中其他成员并研究它们在抵抗病原微生物侵害方面所发挥的功能,旨在为更深入理解无脊椎动物的免疫机制奠定基础,同时为我国水产贝类的绿色养殖提供理论指导。
1 材料和方法
1.1 试验材料
试验用合浦珠母贝购自广西北海市某珍珠养殖场,其壳长(47±5)mm、壳宽(15±3)mm、壳高(52±4)mm、体质量(20±4)g。选取健康有活力的合浦珠母贝个体,分别采集鳃、外套膜、闭壳肌、足和肝胰腺等组织于液氮中冷冻后放至-80 ℃ 冰箱保存备用。
使用的试剂TRIzol、TransScript®Ⅱ All-in-One First-Strand cDNA Synthesis SuperMix for PCR、TransStart®Green qPCR SuperMix、pEASY®-T1 Cloning Kit、Trans5α Chemically Competent Cell购自北京全式金公司,SMARTer RACE 5′/3′Kit购自宝日医生物技术(北京)有限公司。
1.2 试验方法
1.2.1 RNA提取和第一链cDNA合成 采用 TRIzol 法提取各组织RNA,经1%琼脂糖凝胶电泳检测RNA完整性,使用Little Lunatic高通量微流控全光谱分光光度计(美国Unchained Labs)检测RNA浓度。 按照 TransScript®Ⅱ All-in-One First-Strand cDNA Synthesis SuperMix for PCR反转录试剂盒的操作说明合成 cDNA,保存于-20 ℃冰箱备用。
1.2.2 全长cDNA克隆 从合浦珠母贝转录组数据中筛选出C型凝集素基因的同源序列,利用Oligo软件设计引物PfCLEC17A-F和PfCLEC17A-R(表1),以合浦珠母贝转录组数据中表达量最高的肝胰腺cDNA作为模板进行PCR扩增。反应条件:94 ℃ 5 min;94 ℃ 30 s,55 ℃ 30 s 72 ℃ 45 s,30个循环;72 ℃延伸8 min;反应结束后所得PCR产物在1%琼脂糖凝胶上进行电泳检测,并进行回收纯化PCR产物。将回收后的目的DNA片段与pMD18-T质粒连接后导入大肠杆菌DH5α感受态细胞。经菌落PCR确认的阳性克隆菌落送至广州生工测序部进行测序验证,测序结果使用软件DNAMAN 8.0(Lynnon Biosoft,美国)进行拼接和序列比对。针对拼接正确的序列设计特异性引物,用SMARTer RACE 5′/3′Kit(TaKaRa)进行RACE 扩增。首先,按照SMARTer RACE 5′/3′ Kit试剂盒说明书合成3′RACE-cDNA和5′RACE-cDNA,以此cDNA为模板进行巢式PCR扩增,反应体系为25 μL,反应条件:94 ℃ 5 min;94 ℃ 30 s,60 ℃ 30 s,72 ℃ 1 min,30个循环;72 ℃延伸10 min;反应结束后PCR产物经琼脂糖凝胶电泳确认后,进行第2轮PCR,对目的条带进行胶回收纯化、连接转化、挑选阳性菌并测序。

表1 PfCLEC17A扩增及表达所用引物Tab.1 Primers used in amplification and expression of PfCLEC17A
1.2.3 生物信息学分析 测序结果经Chromas软件分析后,在DNAMAN 8.0软件上进行序列拼接,获得合浦珠母贝PfCLEC17A基因的全长序列。利用NCBI ORF Finder在线工具查找开放阅读框和预测编码蛋白序列。采用ProtParam tool在线工具预测编码蛋白的理化特性;使用 SignalP 3.0(https://services.healthtech.dtu.dk/service.php?SignalP-3.0)预测蛋白信号肽序列;采用 NetNGIys 1.0 预测糖基化位点;采用NetPhos 3.1预测蛋白磷酸化位点;采用 SMART 4.0 进行蛋白质结构域分析;使用SWISS-MODEL在线软件进行同源建模,预测蛋白质的三级结构;利用Expasy在线预测蛋白的亲水性/疏水性;利用MegAlign分析序列同源性,在NCBI数据库中搜索不同物种的同源序列,使用Clustal 2.1和GENEDOC软件进行多重序列比对;挑选出最具代表性物种的C型凝集素序列,使用MEGA 7.0 软件的 Neighbor-joining法构建进化树。
1.2.4PfCLEC17A基因的荧光定量PCR分析 选取6只健康的合浦珠母贝并对其进行解剖,取鳃、外套膜、闭壳肌、性腺、血液、足和肝胰腺等7个组织用于分析PfCLEC17A基因在不同组织中的表达情况。以麻醉后注射闭壳肌的方式对合浦珠母贝进行溶藻弧菌攻毒感染,在感染后的第0,1,3,6,12,24,48,72,96小时随机采集6个合浦珠母贝的肝胰腺组织来分析PfCLEC17A基因的表达量变化,以注射0.1 mL PBS的个体作为空白对照组。所有样品取下后立刻放于液氮中速冻,保存于-80 ℃冰箱备用。所有样品的RNA提取和cDNA合成步骤如前所述。cDNA按照 1∶8稀释后作为试验模板,参照全式金公司的TransScript®Green qPCR SuperMix试剂盒说明书进行荧光定量PCR分析,全程操作在冰上进行。每次试验3个样品,每个样品进行3次重复。
1.2.5 荧光定量PCR数据分析 荧光定量PCR结果使用 2-ΔΔCt法进行计算,分析PfCLEC17A基因在合浦珠母贝各组织以及溶藻弧菌刺激后各时间段内的相对表达情况。计算结果使用 IBM SPSS Statistics 22 软件中的LSD(最小显著性差异)法进行统计学分析,P<0.05表示差异显著。
2 结果与分析
2.1 PfCLEC17A基因的克隆与序列分析
通过分析合浦珠母贝的转录组数据,设计PfCLEC17A-F和PfCLEC17A-R引物,扩增获得Ctypelectin17A基因的中间片段,经过测序验证,该片段长度为534 bp(图1-A)。采用5′-RACE技术进行巢式PCR得到RACE产物长度为150 bp(图1-B),经测序验证,除去和ORF重叠的碱基后得到5′ 非编码区(Untranslation region,UTR) 为117 bp。用同样的方法进行巢式PCR,得到3′-RACE产物长311 bp(图1-C),测序验证后3′-UTR为48 bp。将所有测序序列进行拼接,得到PfCLEC17A基因全长为699 bp(图2)。
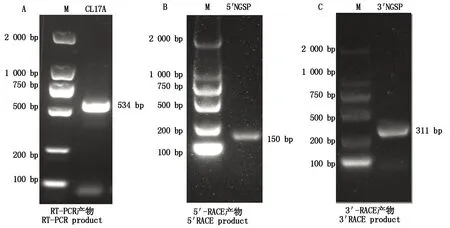
DNA Marker为DL2000。The DNA Marker is DL2000.

双下划线.CLECT结构域;阴影部分.信号肽区域;黄色阴影部分.二硫键;蓝色阴影部分.糖基化位点。
2.2 PfCLEC17A的分子特征鉴定
生物信息学分析显示,PfCLEC17A蛋白理论分子质量为20.326 ku,理论等电点pI为6.11,预测前24个氨基酸为信号肽(图2、图3-A),PfCLEC17A蛋白含有一个经典的CLECT结构域(图3-B),含有17个磷酸化位点,其中2个为Tyr、7个为Ser、8个为Thr位点,还有4个糖基化位点,分别位于N20、N85、N140和N142(图2)。同时还预测到2对保守的二硫键(C16-C119和C93-C111)(图2)。二级结构预测(准确度高于70%)显示,PfCLEC17A蛋白由2个α-螺旋和10个β-折叠组成。三级结构预测的3D模型如图3-C所示。蛋白质亲水/疏水性分析结果显示(图3-D),大部分氨基酸残基的分值为负值(正值代表疏水性,负值代表亲水性),所以PfCLEC17A蛋白为亲水性蛋白。
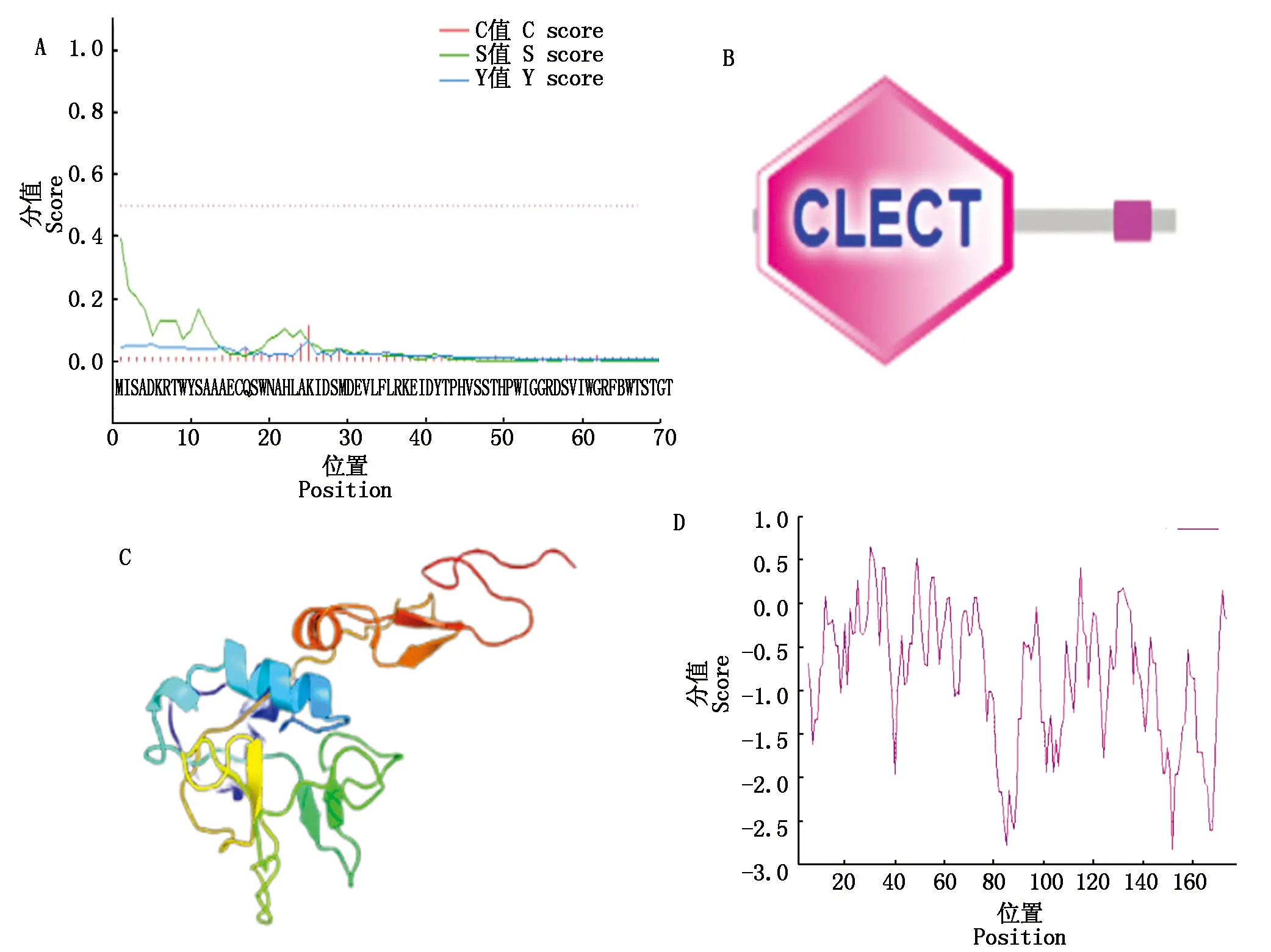
A.蛋白质信号肽预测;B.蛋白质结构域预测;C.蛋白质三级结构预测;D.蛋白质亲水性/疏水性预测。A.Protein signal peptide prediction;B.Protein domain prediction;C.Protein tertiary structure prediction;D.Protein hydrophilicity/hydrophobicity prediction.
2.3 合浦珠母贝PfCLEC17A氨基酸序列比对及系统进化树构建
氨基酸序列比对结果显示,PfCLEC17A与其他物种的C型凝集素氨基酸序列的同源性在14.7%~87.9%。其中,与栉孔扇贝的相似性最高,为87.9%,与紫贻贝的相似性次之,为61.3%,与高原鼠兔的相似性最低,为14.7%。利用ClustalW 2.1和GENEDOC软件将PfCLEC17A蛋白与其他几种贝类蛋白序列进行比对,结果显示(图4),PfCLEC17A的氨基酸序列与其他物种的氨基酸序列具有部分相似的结构,都含有形成二硫键的4个保守的半胱氨酸,糖结合位点几乎都具有独特的“EPN”结构,但是在“WND”位点表现不一致,合浦珠母贝和栉孔扇贝此位点为“WSD”基序,紫贻贝的则为“WDD”基序。

图4 PfCLEC17A氨基酸序列比对Fig.4 PfCLEC17A amino acid sequence alignment
在MEGA 7.0中,以PfCLEC17A蛋白和其他物种的凝集素序列通过邻近法构建的系统发育树显示(图5),白头海雕等几种鸟类聚在一起,智人和高原鼠兔2种哺乳动物聚为一支,合浦珠母贝先与栉孔扇贝聚在同一分支上,然后与其他软体动物聚在一起。以上聚类结果与传统的分类方法相一致,也体现了PfCLEC17A在进化上的保守性。
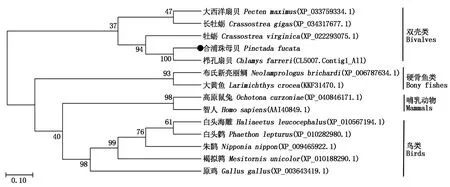
图5 CLEC17A氨基酸序列系统进化树Fig.5 Phylogenetic tree analysis of CLEC17A
2.4 合浦珠母贝PfCLEC17A组织特异性表达分析
采用RT-qPCR方法检测PfCLEC17A基因在合浦珠母贝鳃、外套膜、闭壳肌、肝胰腺、性腺、血细胞和足等不同组织中的表达情况,结果显示(图6),PfCLEC17A在所检测的各组织中均有表达,但相对表达量存在显著差异。在不同组织中的相对表达量大小表现为肝胰腺>鳃>闭壳肌>足>性腺>外套膜>血细胞(P<0.05)。
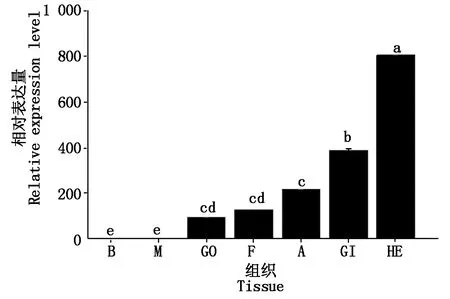
M.外套膜;B.血细胞;GO.性腺;F.足;A.闭壳肌;GI.鳃;HE.肝胰腺。不同字母表示在不同组织中的相对表达量差异性显著(P<0.05)。
2.5 溶藻弧菌感染后合浦珠母贝PfCLEC17A的表达量变化
荧光定量PCR分析结果显示(图7),溶藻弧菌感染1 h后,PfCLEC17A表达水平较对照组显著升高并达到最大值(图7),随后表达量出现回落。除了1 h显著上升(P<0.05)和第12,72小时外,在其他时间,溶藻弧菌感染后PfCLEC17A的相对表达量均低于注射PBS的对照组。由此可见,合浦珠母贝在受到病原微生物侵害初期,肝胰腺中PfCLEC17A基因会被迅速激活,但感染后期,其表达会受到一定程度的抑制。这可能与C型凝集素蛋白家族成员参与免疫激活的早期阶段有关。
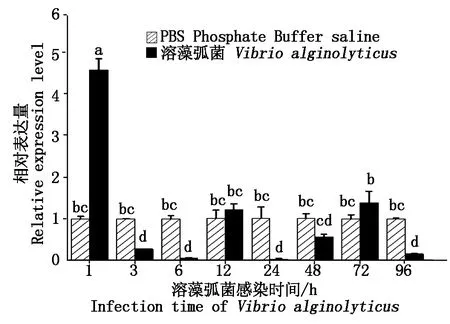
不同字母表示溶藻弧菌不同感染时间下的相对表达量有差异性显著(P<0.05)。Different letters indicate significant differences in relative expression at different time after challenged with V.alginolyticus(P<0.05).
3 结论与讨论
C型凝集素是一类含有钙离子依赖糖识别域的蛋白超家族,根据其结构和功能可以分为17个亚家族,每个成员都有其特有的结构和功能[23-25],除了一些C型凝集素(NK细胞受体家族)没有经典的CRD结构域以外,大多数的C型凝集素受体都是聚糖结合受体,都含有一个或多个CRD识别域[26-27]。本研究获得C型凝集素第17个亚家族成员A的序列,其开放阅读框长534 bp,编码178个氨基酸残基,含有一个经典CRD碳水化合物识别域,有17个磷酸化位点,其中2个Tyr、7个Ser和8个Thr位点。C型凝集素根据参与聚糖识别和钙离子配位的氨基酸基序不同可分为甘露糖基序(EPN,Glu-Pro-Asn)和半乳糖基序(QPN,Glu-Pro-Asp)两类[27]。CLEC17A蛋白中存在EPN和WND 2个糖结合基序,其对甘露糖和Ca2+具有高度特异性[28]。本研究从合浦珠母贝中获得的PfCLEC17A氨基酸序列中含有EPN和在WSD这2种基序。因为EPN基序仅存在于胶原凝集素和选择凝素中[29-30],推测PfCLEC17A可能是一种胶原凝集素或选择凝集素。据报道,第2个基序在脊椎动物中一直是Trp-Asn-Asp(WND),WND也在部分无脊椎动物C型凝集素中存在。第2个基序在软体动物中表示出多样性,目前已有10多个基序被报道,例如WND、Trp-Ile-Asp(WID)、Trp-Ser-Asp(WSD)、Trp-His-Asp(WHD)、Phe-Ser-Asp(FSD)和Leu-Ser-Asp(LSD)等[31-32]。其中EPN基序被认为是与碳水化合物特异性结合的关键开关,第2个基序(WSD)会增加这种结合的亲和力和特异性[31-32]。此外,用软件分析得到序列中含有4个糖基化位点,分别是N20、N85、N140和N142,N-连接糖基化位点可能在CLEC17A向细胞表面的运输和定位中发挥生理作用[20]。
将合浦珠母PfCLEC17A氨基酸序列与其他物种进行同源比对分析并构建进化树,比对结果显示,合浦珠母PfCLEC17A序列与栉孔扇贝的序列相似性最高,为87.9%,与其他贝类相似性大部分在50%~60%,与其他物种序列同源性在10%~30%,与鼠兔的相似性最低仅为14.7%。进化树分析结果显示,合浦珠母PfCLEC17A与其他软体动物亲缘关系较近,与鱼类鸟类的关系较远。将合浦珠母与其他物种的CTL所编码的氨基酸序列进行同源性和相似性比对,发现它们的同源性与相似性不高,仅凝集素特有的基序及功能位点基本保持一致。在其他贝类C型凝集素同源比较研究也发现类似结论[33-35],由此可见,C型凝集素是一类进化较快但功能位点保守的蛋白质。
本研究检测了PfCLEC17A基因在不同组织中的表达情况,在所检测组织中均有表达,其中在肝胰腺中表达最高。目前,关于贝类CLEC17A基因的研究较少,但是关于贝类C型凝集素家族其他成员的研究已有相关报道,如马氏珠母贝PmLec-8基因在各组织中均有表达,在肝胰腺中表达量最高,其次是鳃[22];在海湾扇贝中,C型凝集素在肝胰腺中表达量最高,血淋巴表达量最低,在其他组织中均有所表达[36];在关于菲律宾蛤仔[37]的研究中,6个C型凝集素基因分别在鳃和肝胰腺中显著高表达。最近在菲律宾蛤蜊中鉴定了VpClec-3和VpClec-4共2种C型凝集素基因,它们在肝胰腺和鳃组织中均高水平表达[17]。以上研究显示,C型凝集素在多种组织中表达,说明它在多种病原免疫反应中扮演重要角色。但在不同物种的不同组织中表达量不尽相同,可能是由不同物种C型凝集素的分布及功能不同造成的。肝胰腺不仅是软体动物的主要免疫器官,也是消化代谢最旺盛的器官,大多数免疫相关基因在肝胰腺中都有表达[37-38]。其次,鳃是软体动物免疫防御的第一道防线,与外界广泛接触,对环境中的有害物质起到一定过滤和抵御作用[39],PfCLEC17A在合浦珠母贝的肝胰腺和腮中的相对表达量都很高,可能在机体抵御外界病原微生物入侵过程中发挥重要作用。
在溶藻弧菌刺激下,合浦珠母贝PfCLEC17A基因在肝胰腺中的表达量迅速升高,这与魁蚶Sb-Lec1基因[40]和文蛤Mm-CTL基因[41]的表达模式相同,说明C型凝集素基因的表达受微生物的诱导,合浦珠母贝PfCLEC17A基因参与机体的前期免疫应答。但是,PfCLEC17A基因在接收病原菌刺激后如何将信号传递给下游分子并改变基因表达,这方面的工作还需进一步研究。本研究初步对PfCLEC17A基因功能进行了探索,为后续深入了解凝集素在合浦珠母贝免疫防御系统中的作用奠定基础。
本研究通过RACE技术获得了PfCLEC17A基因的全长序列,并对其进行生物信息学分析,通过RT-qPCR技术检测该基因在不同组织中的表达情况,结果显示,该基因在肝胰腺中表达量较高。进一步检测溶藻弧菌刺激后合浦珠母贝肝胰腺中PfCLEC17A基因在不同时间段的变化情况,发现其在弧菌感染后表达量迅速上升并达显著水平,以上结果表明,该基因分布广泛,并在宿主和病原体相互作用的前期发挥重要作用。
