抗(感)谷瘟病谷子品种内生菌多样性分析
2022-11-03刘佳悦贾丽霞王苗苗孙东辉孙贺贺郝志敏李志勇
刘佳悦,贾丽霞,王苗苗,孙东辉,孙贺贺,郝志敏,李志勇
(1.河北省农林科学院 谷子研究所,河北 石家庄 053000;2.河北农业大学 生命科学学院,河北 保定 071001;3.河北省农林科学院 农业信息与经济研究所,河北 石家庄 050051;4.邢台现代职业学校,河北 邢台 054000)
内生菌是生活史的全部或部分阶段存在于健康植物组织和器官,且不引起寄主植物损伤的微生物[1-2]。研究表明,健康植物中存在多种多样的微生物群落,它们与宿主共同进化,同时伴随种子萌发的过程,可迁移到不同组织及器官中,对植物的有益性状具有调节作用。其中的有益细菌和真菌,如拮抗细菌、菌根真菌及固氮菌等,可以通过促进植物营养获取、提高植物抗逆性以及对病原菌的抵抗力,影响植物的生长和健康[3-5]。
近年来,随着人们对农作物品质要求的提高,能够促进作物生长及提高其抗逆性的新型微生物资源受到了越来越多的关注,这些微生物与植物之间的相互作用已成为植物学、微生物学、生态学等多个领域的研究热点。相关研究证实,植物内生菌可在种子中占据多个生态位[6],在种子萌发过程中,首先定殖在根部,还可伴随着细胞的移动扩散到包括茎、叶的其他植物组织[7]。从整体角度来看,不同部位具有不同的菌群结构。而不同微生物群体与植物各个组织在长期进化的过程中形成了互惠共生的关系,在植物抵御不良环境及病原菌的侵染过程中均可发挥巨大的作用[8-10]。同时,这些结果也可表明内生菌在时间、空间上会产生系统性变化,进一步将不同生态位微生物群体变化与宿主感病性及抗病性联系在一起,有助于揭示不同内生菌种类在寄主感病或抗病过程中发挥的作用。
谷子(Setariaitalica)属禾本科狗尾草属,一年生草本植物,具耐旱、适应性强、抗逆范围广等特征,在我国北方广泛种植。因其具有较高的营养价值,常作为北方人民的主要杂粮作物之一[11-12]。谷瘟病是由灰梨孢菌(Magnaportheoryzae)引起的病害,谷子各生育阶段都可发生[13]。近年来,由于生产中缺乏抗病品种,单一品种大面积的种植,谷瘟病已经成为影响我国谷子生产的重要病害之一。而且,田间灰梨孢菌有多个生理小种,容易发生生理小种变异,导致谷子抗病性丧失。而化学农药的大量使用,容易造成环境污染和农药残留,并且容易使病原菌出现抗药性,因此,急需新的防治策略来防控谷瘟病[14-15]。自然条件下,谷子及其内生菌和谐共生,形成原始微生态环境,病害发生后导致谷子原始微生态平衡破坏,究竟哪些内生菌种群发生变化及不同部位谷子内生菌群动态有何变化,目前尚未知晓,而这些内生菌种群的明确是研究谷子与相关菌群互作的重要内容,对于生物防治制品[16]的研发、抗谷瘟病品种的选育及谷子产量的提高具有重要意义。
为了更深入地了解谷子不同品种与品种及同一品种不同器官内生细菌的群落结构和多样性特征,本研究以4个不同品种的谷子为研究对象,分别收集、提取其茎、叶片、叶鞘3个部位DNA,通过Illumina Miseq 高通量测序对内生菌群的结构组成进行分析,以期解析不同品种谷子及其不同组织内生菌群落结构组成的差异性,揭示植物内生菌群落结构特征,进一步筛选谷子不同品种及部位功能微生物,为内生菌资源的挖掘与利用奠定基础。
1 材料和方法
1.1 试验材料
本试验选取感谷瘟病的沙湾谷子、冀谷22和抗谷瘟病的小青谷、石榴子共4个不同品种的谷子为研究对象(表1)。谷子播种于营养土中,在人工气候箱种植,10 h光照 26 ℃/14 h黑暗 22 ℃,在抽穗期时,接种谷瘟病菌强毒菌株10-52,待感病品种发病后,采集各品种的叶片、茎、叶鞘共计12种样品,-80 ℃保存,备用。

表1 不同品种谷子取样信息Tab.1 Sampling information of different foxtail millet varieties
1.2 16S rDNA PCR扩增及高通量测序
用FastDNA SPIN Kit for Soil提取植物样品基因组DNA,利用1%琼脂糖凝胶电泳检测抽提的基因组DNA。按16S rDNA V3—V4区,合成带有barcode的特异引物,进行PCR扩增。每个样本3个重复,将同一样本的PCR产物混合后用2%琼脂糖凝胶电泳检测,使用AxyPrepDNA凝胶回收试剂盒(AXYGEN公司)切胶回收PCR产物,Tris-HCl洗脱;2%琼脂糖电泳检测。构建Miseq文库,通过诺禾致源novaseq6000平台进行高通量测序,上海美吉生物云平台QiiME1(http://qiime.org/install/index.html)进行数据分析。
1.3 数据处理
Miseq测序得到的PE首先根据overlap关系进行拼接区分样本后经Uparse(http://www.drive5.com/uparse/)及Usearch数据库进行OTU聚类分析和物种分类学分析,Silva(Release138http://www.arb-silva.de)数据库进行16S细菌和古菌核糖体比对分析,Unite(Release 8.0 http://unite.ut.ee/index.php)数据库进行真菌多样性比对。为了得到每个OTU对应的物种分类信息,采用RDP classifier(Version 2.11http://sourceforge.net/projects/rdp-classifier/)贝叶斯算法对97%相似水平的OTU代表序列进行分类学分析,并分别在各个分类学水平统计各样本的群落物种组成。基于OTU聚类分析结果,对OTU进行alpha多样性分析(https://www.mothur.org/wiki/Down),PICRUSt(http://picrust.github.io/picrust/)数据库对16S序列进行KEGG、COG、Pfam功能预测;基于分类学信息,在各个分类水平上进行群落结构的统计分析。经美吉生物云平台对原始数据进行处理。为保证在后续分析中数据的均一性,将有效序列按不同分组最小样本序列数目进行抽平。
2 结果与分析
2.1 谷子内生细菌群落结构分析
为了得到每个OTU对应的物种分类信息,采用RDP classifier贝叶斯算法对97%相似水平的OTU代表序列进行分类学分析,并分别在各个分类学平:Domain(域)、Kingdom(界)、Phylum(门)、Class(纲)、Order(目)、Family(科)、Genus(属)、Species(种)统计各样本的群落物种组成。经物种组成分析(图1),门水平上,供试样本内生菌优势种类均为革兰氏阳性菌变形菌门及革兰氏阴性菌放线菌门(Actinobacteriota)。拟杆菌门(Bacteroidota)、绿弯菌门(Choroflexi)、粘菌门(Myxococcota)、厚壁菌门(Firmicutes)相对丰度次之。

G1-1.沙湾谷子叶片;G3-1.沙湾谷子茎;G4-1.沙湾谷子叶鞘;G1-2.冀谷22叶片;G3-2.冀谷22茎;G4-2.冀谷22叶鞘;G1-4.小青谷叶片;G3-4.小青谷茎;G4-4.小青谷叶鞘;G1-5.石榴子叶片;G3-5.石榴子茎;G4-5.石榴子叶鞘。
2.2 谷子内生菌Alpha多样性分析
以抽取的数据量为横坐标,以Alpha多样性指数值为纵坐标绘制曲线,根据曲线是否达到平缓来判断本次测序数据量是否足够。对沙湾谷子、冀谷22号、小青谷、石榴子样品进行Alpha多样性分析,绘制稀释曲线,曲线趋于平缓,说明本次测序数据量充足(图2)。
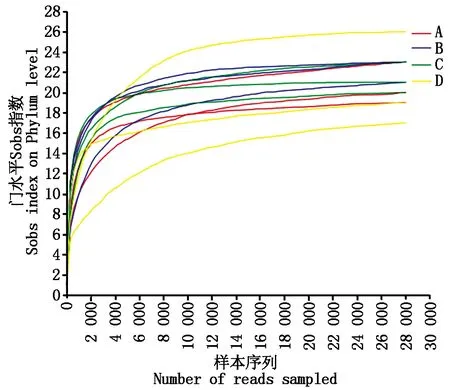
A~D.沙湾谷子、冀谷22号、小青谷、石榴子。图4~5同。A—D.Shawan millet,Jigu 22,Xiaoqinggu,Shiliuzi.The same asFig.4—5.
如图3所示,Sobs指数反映出感病品种(沙湾谷子、冀谷22)叶片群落丰富度较高,石榴子(抗谷瘟病)在叶鞘时期丰富度较沙湾谷子(感谷瘟病)占有优势。冀谷22(感谷瘟病)这一品种茎的Shannon指数较高,群落多样性及特有OTU数目较高;沙湾谷子叶片群落多样性高于其他几个品种;小青谷(抗谷瘟病)叶鞘物种多样性较高。说明不同品种谷子内生菌在物种组成上具有较大差异,感病品种叶片、茎部微生物多样性较高;抗病品种叶鞘内生菌多样性相对丰富。

A~C.叶片、茎、叶鞘;G-1.沙湾谷子;G-2.冀谷22号;G-4.小青谷;G-5.石榴子。A—C.Leaves,stems,leaf sheath;G-1.Shawan millet;G-2.Jigu 22;G-4.Xiaoqinggu;G-5.Shiliuzi.
2.3 谷子内生菌Beta多样性分析
通过主坐标分析(Principal co-ordinates analysis,PCoA分析),评估不同品种及不同谷子内生菌群落结构的Beta多样性,用来研究样本群落组成的相似性或差异性,与PCA分析类似;图4结果显示,谷子不同部位对内生菌群落结构影响更大(76.53%,P<0.001);从谷子不同部位分析菌群群落组成可看出,不同品种谷子同一部位菌群含量有一定的差异性,沙湾谷子及冀谷22这2个感病品种含有与小青谷、石榴子(抗谷瘟病)差异较大的内生菌群,推测其可能与谷子谷瘟病抗性相关;而不同部位之间菌群含量又具有很大的相似性,说明部分内生菌在植株生长过程中会产生迁移。冀谷22的茎在PC轴与其他3个品种距离较远,4个品种的叶鞘在PC轴上距离相对叶片与茎较近,不同品种的叶片在PC轴上距离相对均一,2个感病品种(沙湾谷子、冀谷22)叶片内生菌群距抗病品种(小青谷、石榴子)较远,同Alpha多样性分析结果一致,进一步验证不同品种谷子及同一品种谷子不同部位内生菌群落组成差异较大,叶片较茎、叶鞘菌群差异性高。同时部分菌群可在不同生长时期迁移到不同部位。说明器官类型对内生菌群结构的影响大于对品种的影响。
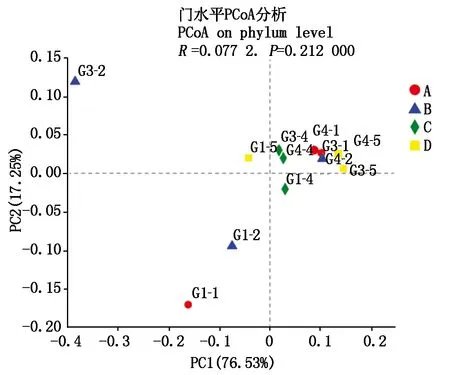
G1.叶片;G3.茎;G4.叶鞘。G1.Leaves;G3.Stems;G4.Leaf sheath.
2.4 同一部位不同品种谷子内生菌菌群物种组成分析
通过对内生菌群分布情况进行分析发现,4个品种谷子叶片共有内生菌属16个门,占比最高的门为变形菌门、放线菌门、拟杆菌门、厚壁菌门,其中,冀谷22优势菌所属门为弯曲菌门,沙湾谷子优势菌门为肠杆菌门(Entotheonellaeota),石榴子独有内生菌属梭杆菌门(Fusobacteriota);茎部优势菌门同叶片一致,沙湾谷子独有内生菌属螺旋体门(Spirochaetota),冀谷22在茎部独有内生菌属脱硫菌门(Desulfobacterota)及肠杆菌门,小青谷优势菌门为Latescibacterota;4个品种叶鞘共有内生菌同属16个门,其中沙湾谷子优势菌门为Armatimonadota,冀谷22(感谷瘟病)独有内生菌所属门为肠杆菌门,石榴子独有内生菌属Methylomirabilota(图5)。
综上,感病品种及抗病品种谷子在同一部位的内生菌群分布差异较明显,同时,感病品种都具有肠杆菌门这一菌门。此外,冀谷22(感谷瘟病)在叶片、茎及叶鞘内生菌群结构组成丰富度为4个品种中最高,石榴子(抗谷瘟病)在叶鞘中独有内生菌多样性较高。
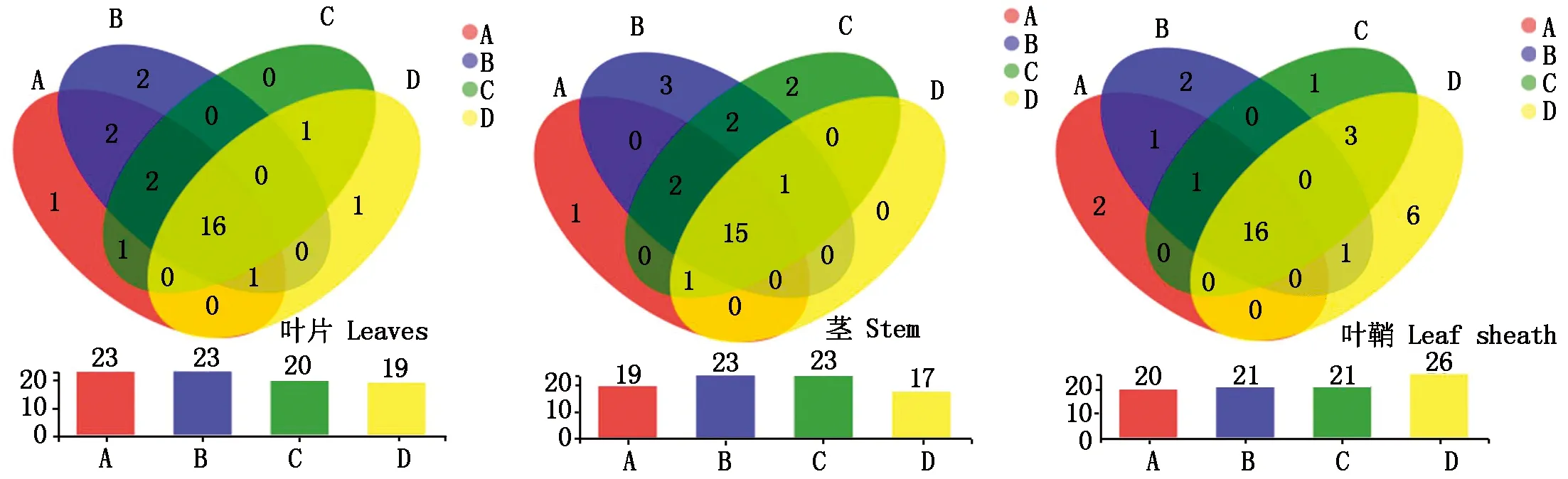
图5 OTU分布的Veen图分析Fig.5 Veen plot analysis of the OTU distribution
2.5 同一品种不同部位谷子内生菌菌群物种组成分析
通过对谷子不同部位内生菌菌群物种组成分析发现,同一品种不同部位谷子内生菌菌群组成有较大差异,沙湾谷子的茎、叶片、叶鞘独有内生菌为变形菌门、放线菌门、拟杆菌门、厚壁菌门,其中叶片独有内生菌数目属4个门,分别为NB1-j、Methylomirabilota、脱硫菌门、肠杆菌门,茎优势菌属Spirochaetota,叶鞘独有内生菌属Latescibacterota、Sumerlaeota,沙湾谷子叶片独有内生菌门丰富度较高;冀谷22号叶片独有内生菌所属门为MBNT15、Campilobacterota,茎特有内生菌属3个门,分别为梭杆菌门、脱硫菌门、Sumerlaeota,叶鞘特有内生菌属Latescibacterota;小青谷4个部位特有内生菌属15个门,其中叶片独有内生菌属脱硫菌门,茎NB1-j、Methylomirabilota,叶鞘SAR324_cladeMarine_group_B、Hydrogenedentes;石榴子叶片、茎、叶鞘共有内生菌属16个门,叶片独有内生菌属Armatimonadota,Methylomirabilota、Elusimicrobiota、Hydrogenedentes、Dependentiae、脱硫菌门、Synergistota、Dadabacteria为石榴子叶鞘特有内生菌所属门。结果与上述分析一致,4个品种不同部位优势内生菌所属门相同,2个感病品种在叶片、茎、叶鞘3个部位共有OTU数目较抗病品种多,推测其中部分菌属可能与感病性相关(图6)。
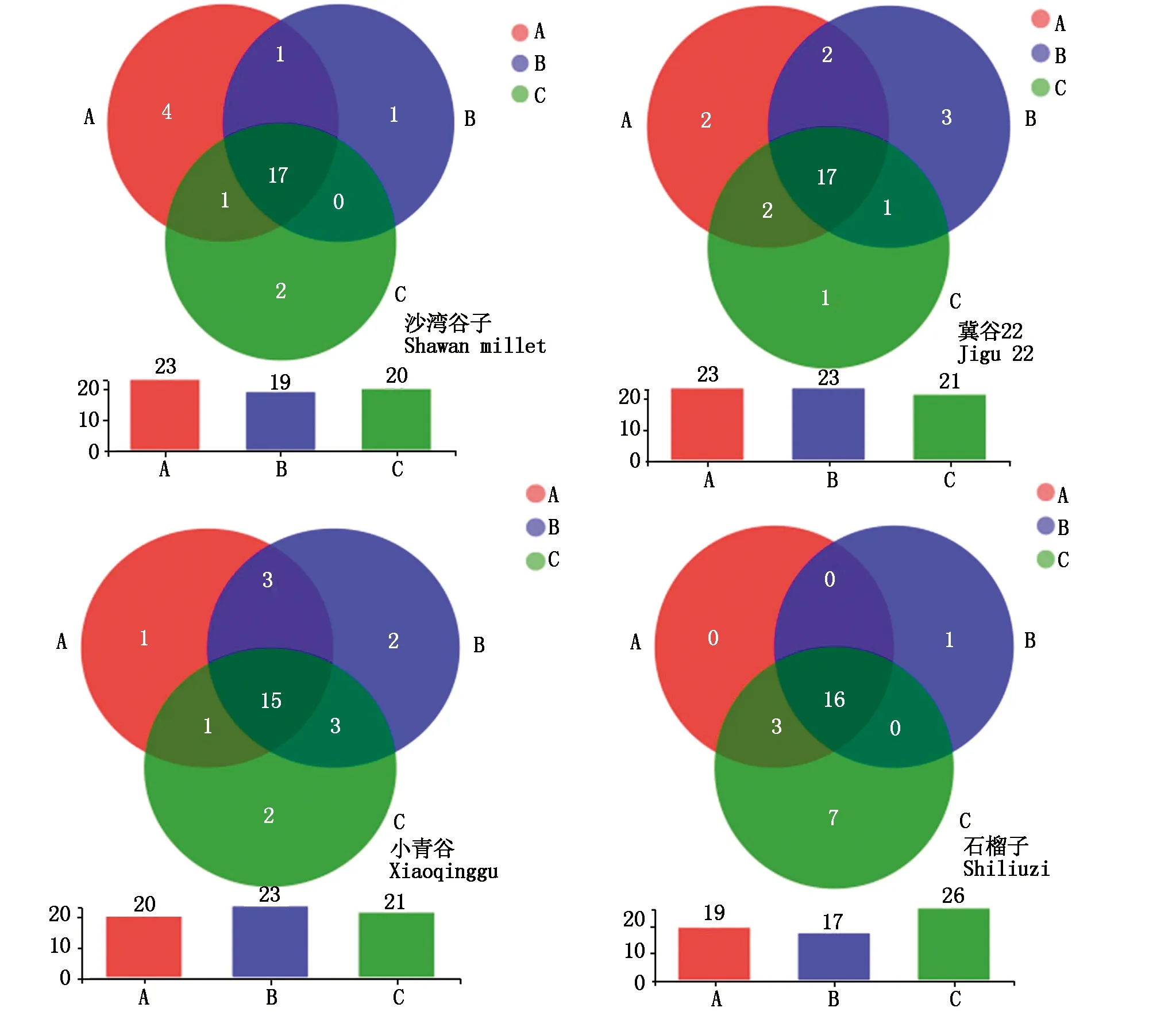
A~C.谷子的叶片、茎、叶鞘。A—C.Leaves,stem and leaf sheath of foxtail millet.
每一品种谷子在叶片、茎、叶鞘这3个部位所含内生菌种类不一,推测叶片接触外界环境范围较广,菌群可通过外界环境进入植物组织内部,造成内生菌多样性存在一定差异性。
3 讨论
由于新型病虫害的不断出现、化肥大量使用以及全球气候的剧烈变化,造成环境恶化严重威胁农业生产的稳定性。植物内生菌的分离鉴定及利用将为可持续农业发展奠定一定的基础。环境中,植物可与多种内生菌共存,部分内生菌可增加宿主植物固氮、产铁载体、获取养分、产生长激素的能力[17-18]。利用微生物提高植物抵御生物胁迫的能力迫在眉睫。
核心微生物是形成植物相关微生物组的关键,不仅可促进植物自身生长,还可招募有益微生物帮助其定殖调节植物-微生物或微生物-微生物之间的相互作用[19-20],从而达到抑制某些病害发生的目的。本研究首次采用Illumina Miseq高通量测序法对4个品种谷子及其3个不同部位16S rDNA的V3—V4区进行内生菌群落结构与多样性分析,研究表明,感(抗)谷瘟病品种内生菌群落核心微生物均为变形菌门、放线菌门、拟杆菌门、厚壁菌门,这与Jin等[21]对谷子根部微生物关联分析结果一致。其中,拟杆菌门具有降解纤维素的能力;绿弯菌门为兼性厌氧生物,在光合作用中不产生氧气、不能固氮,利用3-羟基丙酸途径固定二氧化碳;粘菌门由于原生质体裸露,繁殖迅速并较易培养,可作为研究原生质和生殖生理的一种材料。邹媛媛等[9]对多个水稻品种种子内生菌进行比较发现,优势菌属为泛菌属(Pantoea)和芽孢杆菌属(Bacillus)。经COG功能分析,各个品种谷子内生菌功能丰富,包括参与RNA加工和修饰;能量产生和转换、细胞周期控制、细胞分裂、染色体分裂、氨基酸转运和代谢;核苷酸转运和代谢;碳水化合物转运和代谢;辅酶转运和代谢;脂质转运和代谢;翻译、核糖体结构和生物发生;转录、复制、重组和修复;次生代谢物的生物合成、转运和分解代谢等。
通过对不同品种谷子内生菌群落结构进行分析发现,不同品种谷子内生菌物种组成具有一定的差异性,经Alpha多样性分析发现,感病品种(沙湾谷子、冀谷22)在叶片及茎部群落多样性及丰富度较高,可对其特有内生菌群进行致病性分析,研究谷瘟病发病机制,降低谷瘟病发病率;在叶鞘时期,石榴子这一抗病品种丰富度较沙湾谷子占有优势,对抗病品种特有菌群进一步分析,对抗病品种选育具有重要意义。小青谷叶鞘物种多样性较高。结果表明,不同品种谷子在物种组成上具有较大差异。从谷子不同部位分析菌群群落组成,发现不同部位菌群差异性较品种间差异对内生菌群落结构影响更大,而Jin 等[21]通过对谷子根微生物群的分类结构和功能分析发现,微生境和地理位置也是影响谷子根系微生物区系变化的主要因素,其中,微生境对β多样性变化的解释比例最大,其次是地理位置。不同品种的叶片在PC轴上距离相对均一,2个感病品种(沙湾谷子、冀谷22)叶片内生菌群距抗病品种(小青谷、石榴子)较远,进一步验证Alpha多样性及Beta多样性分析结果:谷子部分内生菌群落在植株生长过程中会产生迁移,迁移过程中,由茎至叶鞘再到叶片,而叶片在结构与功能方面均与叶鞘及茎有较大差异,且叶片受外界环境影响较大,菌群演变更为丰富,因此形成了明显不同的内生菌群。物种分析中发现,感病品种微生物多样性高于抗病品种,尤其发现了大量感病品种或抗病品种中特有的内生菌类群,这些内生菌对菌群结构的形成及其对寄主谷瘟病抗感特性的影响有待于进一步试验探索。
综上,本研究初步揭示了谷子不同品种及器官的内生菌群落结构特征,为相关功能菌株的筛选、应用及抗病品种的选育奠定了理论基础。
