miR-338靶向CaMKⅡγ调控大鼠神经病理性疼痛的作用机制研究
2022-10-31马真宋海亮谢然尖措赵吉彪
马真 宋海亮 谢然尖措 赵吉彪
神经病理性疼痛是中枢神经系统发生损伤或疾病引起的一种慢性疾病,影响30%至50%的世界人口,是患者、社会和医疗领域的重要负担[1,2]。神经病理性疼痛发病机制复杂,有发病率高、现有治疗方案不足、临床干预效率较低等特点,是一件非常棘手的问题,因此需要进一步研究以确定和验证潜在的靶点[2,3]。多项研究表明神经病理性疼痛与炎性反应密切相关,机体炎性因子的大量释放严重损害神经细胞及非神经细胞的生长,影响神经系统功能的恢复,最终引起机体的痛觉异常[4,5],因此从降低机体炎性反应、缓解疼痛等方面寻找相关的治疗靶点是有效的。微小RNA(microRNA,miRNA)是一类内源性非编码单链RNA,长度约为22个核苷酸,参与调节神经系统中相关基因的表达,在维持神经系统功能等方面发挥重要作用,可通过调节神经炎症来影响神经病理性疼痛的发生发展过程[6]。miR-338-3p是miRNA家族成员之一,参与炎性反应等多种生物学进程的调控。钙调蛋白依赖性蛋白激酶Ⅱα(Ca2+/calmodulin-dependent protein kinase Ⅱα,CaMKⅡα)是细胞内传递Ca2+信号的关键分子,具有α、β、γ和δ四种异构体。Zhang等[7]研究发现miR-338-3p通过阻止炎症信号通路的异常激活而抑制促炎性细胞因子的表达,并认为可能是miR-338-3p的直接靶点,但关于miR-338-3p与CaMKⅡγ之间的研究较少,因此本研究通过构建神经病理性疼痛大鼠模型,探究miR-338-3p调控神经病理性疼痛的可能作用机制。
1 材料与方法
1.1 动物与细胞 55只SD雄性大鼠购于上海吉辉实验动物饲养有限公司,均为SPF级,体重约为240 g,生产许可证号:SCXK(沪)2017-0012。大鼠均在本院动物房内饲养,动物房通风、光照条件良好,温度约25℃,实验期间自由进食进水,每周清洁1次鼠笼并更换新鲜垫料,每天清洁1次饮水瓶。大鼠背根神经元细胞(货号:CP-R126)购于武汉普诺赛生命科技有限公司。
1.2 主要试剂与仪器 总RNA提取试剂盒、反转录试剂盒、肿瘤坏死因子α(tumor necrosis factor α,TNF-α)、干扰素γ(interferon γ,IFN-γ)、白介素1β(interleukin 1β,IL-1β)ELISA试剂盒、双荧光素酶报告基因检测试剂盒(货号:R1200、RP1105、SEKR-0009、SEKR-0008、SEKR-0002、D0010)购于北京索莱宝科技有限公司;miR-388-3p类似物(mimic)及其阴性对照物(mimic NC)均油广州锐博生物科技有限公司合成;CaMKⅡγ抗体、GAPDH抗体、羊抗兔IgG(货号:ab262701、ab181602、ab6721)购自美国Abcam;EnVision多功能酶标仪购于PerkinElmer公司;I-U-004爪触觉测试仪、38450电子触觉测量仪购于意大利UGO BASILE公司等。
1.3 方法
1.3.1 动物模型制备及分组处理:参考文献[8]制备神经病理性疼痛大鼠模型,将55只大鼠按照随机数字法分为假手术组(10只)和造模组(45只)。将大鼠麻醉后进行表皮常规消毒,纵向切开大鼠腰部5~6间隙后逐层分开,使L5白色椎板暴露,用针头刺破髂棘(L5~L6棘突间隙)部位,将PE-10导管从刺破处置入约2 cm,再将导管外端经皮下移至颈部并固定,大鼠出现甩尾、抖动反射、有清亮脑脊液流出等现象表明置管成功,置管成功后3 d剔除瘫痪、状态不佳或运动障碍的大鼠;再次将大鼠麻醉,于大鼠背部正中做切口,暴露L5和L6脊神经,分离并采用4-0缝线结扎L5、L6脊神经,当出现小腿肌肉轻度颤动时表明结扎强度适宜,随后缝合大鼠肌肉及皮肤,假手术组仅分离L5和L6脊神经,不做结扎处理。造模过程中观察并记录大鼠一般情况,当大鼠出现舔舐后爪、后爪收缩、跛行、足趾并拢等现象表明神经病理性疼痛模型构建成功。造模过程中,造模组出现3只大鼠置管后状态不佳,2只死亡,共造模成功40只。
1.3.2 将造模成功的40只大鼠按照随机数字法分为模型组、阴性对照组、miR-338-3p组、miR-338-3p+CaMKⅡγ组,每组10只。参考文献[9],miR-338-3p组在造模第3、5、7天鞘内注射4 mg/kg的miR-338-3p mimic,miR-338-3p+CaMKⅡγ组共注射miR-338-3p mimic、CaMKⅡγ mimic,阴性对照组鞘内注射等量mimic NC,模型组及假手术组注射等量0.9%氯化钠溶液。
1.4 大鼠行为学测定 分别在造模第0、2、6、10、14天测定大鼠机械痛觉阈值(mechnical withdrawal threshold,MWT)及热缩足潜伏期(thermal withdrawal latency,TWL)。提前将大鼠放入爪触觉测试仪中30 min,待其适应环境后用电子触觉测量针刺激大鼠的足底部,在大鼠缩足时记录仪器中电子触觉测量针刺激力度,即为大鼠MWT值。将大鼠置于足底热刺激仪透明塑料笼中适应30 min,用热辐射刺激仪透过玻璃板照射大鼠足底部,记录大鼠从照射开始到其缩足的时间,即为大鼠TWL值,为防止烫伤大鼠,将时间上限设置为30 s,每只大鼠重复测量5次,每次间隔5 min[10]。
1.5 大鼠背根神经节标本采集 行为学测定结束后,将大鼠麻醉,用含有多聚甲醛及戊二醛的磷酸缓冲液进行心脏灌注,灌注至大鼠四肢僵硬,无菌环境下取出大鼠L5~L6背根神经节,低温保存,用于后续实验测定。
1.6 大鼠背根神经节中miR-338-3p、CaMKⅡγ mRNA表达情况 取1.5中大鼠部分背根神经节组织,用总RNA提取试剂盒提取其中的总RNA,取1.5 μl背根神经节组织RNA提取液,用反转录试剂盒获得cDNA,对miR-338-3p、CaMKⅡγ进行扩增,用实时荧光定量PCR测定,反应体系共20 μl:10 μl 2×SYBR Mix,8 μl H2O,上下游引物各0.5 μl,1 μl 10×cDNA模板;反应条件:95℃(5 min)、95℃(1 min)、62℃(1 min)、72℃(2 min),40个循环。以U6为内参,根据2-ΔΔCT算法分析miR-338-3p、CaMKⅡγ的表达水平,U6:上游引物5’-CTCGCTTCGGCAGCACA-3’,下游引物5’-AACGCTTCACGAATTTGCGT-3’;miR-338-3p:上游引物5’-UUUGCGGAAUUUUAA AUGCUGGA-3’,下游引物5’-GUUGUUUUAGUGACUACGACCU-3’;CaMKⅡγ:上游引物5’-ATGGCCACCACCGCCACCT-3’,下游引物5’-CTGCAGCGGTGCGGCAGGG-3’。
1.7 大鼠背根神经节中CaMKⅡγ蛋白表达情况 取1.5中大鼠部分背根神经节组织,用RIPA裂解液提取其中的总蛋白,用BCA试剂盒对提取的总蛋白进行定量。取30 μg蛋白沸水浴变性后,上样经10%SDS-PAGE电泳后电转至PVDF膜,5%脱脂奶粉封闭2 h,加入CaMKⅡγ抗体(1∶1 500)、GAPDH抗体(1∶1 500)并4℃孵育过夜,加入二抗羊抗兔IgG(1∶2 000)于室温下孵育1 h,化学发光法显色显影,扫描图像,分析条带灰度值,计算CaMKⅡγ蛋白与GAPDH蛋白的灰度值比值。
1.8 大鼠背根神经节中炎性因子表达水平测定 取1.5中大鼠部分背根神经节组织,加入适量生理盐水,研磨成10%的组织匀浆,离心取上清液,根据肿瘤坏死因子-α(TNF-α)、干扰素-γ(IFN-γ)、白介素1β(IL-1β)相关试剂盒的操作步骤进行背根神经节组织中TNF-α、IFN-γ、IL-1β表达水平的检测。
1.9 荧光素酶报告实验检测 在TargetScan数据库中分析预测miR-338-3p与CaMKⅡγ的靶向作用位点。构建突变型(MUT)和野生型(WT)基因靶点CaMKⅡγ的3’-UTR荧光素酶表达载体,将购买的大鼠背根神经元细胞快速融化,经过消化、传代后在37℃、5%CO2培养箱中进行培养,采集处于对数生长期的细胞,将CaMKⅡγ-MUT、CaMKⅡγ-WT重组载体分别与miR-338-3p mimic及mimic NC共转染至大鼠背根神经元细胞中,转染24 h后收集细胞,测定荧光素酶活性,实验重复3次。
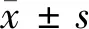
2 结果
2.1 大鼠一般情况观察 假手术组大鼠健康状况良好,行为学观察无明显变化;与假手术组比较,模型组大鼠出现甩足、舔足、足趾并拢等现象;与假手术组比较,miR-338-3p组大鼠行为学得到一定改善。
2.2 miR-338-3p与CaMKⅡγ靶向关系预测及验证 TargetScan数据库结果显示,miR-338-3p在CaMKⅡγ mRNA 3’UTR存在相应的结合位点;荧光素酶报告实验结果显示,与CaMKⅡγ WT+mimic NC组比较,CaMKⅡγ WT+miR-338-3p mimic组大鼠背根神经元细胞荧光素酶活性显著降低(P<0.05),CaMKⅡγ MUT+mimic NC组、CaMKⅡγ MUT+miR-338-3p mimic组比较差异无统计学意义(P>0.05)。见表1。
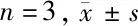
表1 荧光素酶报告实验检测miR-338-3p与CaMKⅡγ的靶向作用关系
2.3 5组大鼠MWT、TWL值比较 假手术组大鼠手术前后MWT、TWL值无明显变化,其余组大鼠造模后MWT、TWL值明显降低,之后随着时间的推移逐渐升高。造模第0天,5组大鼠MWT、TWL值比较差异无统计学意义(P>0.05)。造模第2天,与假手术组比较,模型组、阴性对照组、miR-338-3p组、miR-338-3p+CaMKⅡγ组大鼠MWT、TWL值均显著降低(P<0.05)。造模第6、10、14天,与假手术组比较,模型组大鼠MWT、TWL值均显著降低(P<0.05);与模型组相比,阴性对照组差异无统计学意义(P>0.05),miR-338-3p组及miR-338-3p+CaMKⅡγ组大鼠MWT、TWL值均显著升高(P<0.05);与miR-338-3p组比较,miR-338-3p+CaMKⅡγ组大鼠MWT、TWL值均显著降低(P<0.05)。见表2。
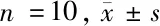
表2 5组大鼠MWT、TWL值比较
2.4 5组大鼠背根神经节组织中miR-338-3p、CaMKⅡγ mRNA及蛋白表达情况比较 与假手术组比较,模型组大鼠背根神经节组织中miR-338-3p表达水平显著降低(P<0.05),CaMKⅡγ mRNA及蛋白表达水平显著升高(P<0.05);与模型组比较,阴性对照组差异无统计学意义(P>0.05),miR-338-3p组及miR-338-3p+CaMKⅡγ组大鼠背根神经节组织中miR-338-3p表达水平显著升高(P<0.05),CaMKⅡγ mRNA及蛋白表达水平显著降低(P<0.05);与miR-338-3p组比较,miR-338-3p+CaMKⅡγ组大鼠背根神经节组织中CaMKⅡγ mRNA及蛋白表达水平显著升高(P<0.05)。见图1,表3。

图1 大鼠背根神经节组织中CaMKⅡγ表达情况
表3 5组大鼠背根神经节组织中miR-338-3p、CaMKⅡγ mRNA及蛋白表达情况比较
2.5 5组大鼠背根神经节组织中炎性因子表达水平比较 与假手术组比较,模型组大鼠背根神经节组织中TNF-α、IL-1β、IFN-γ表达水平显著升高(P<0.05);与模型组比较,阴性对照组差异无统计学意义(P>0.05),miR-338-3p组及miR-338-3p+CaMKⅡγ组大鼠背根神经节组织中TNF-α、IL-1β、IFN-γ表达水平显著降低(P<0.05);与miR-338-3p组比较,miR-338-3p+CaMKⅡγ组大鼠背根神经节组织中TNF-α、IL-1β、IFN-γ表达水平显著升高,差异均有统计学意义(P<0.05)。见表4。
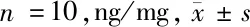
表4 5组大鼠背根神经节组织中TNF-α、IFN-γ、IL-1β表达水平比较
3 讨论
神经病理性疼痛是以中枢神经系统或周围神经系统病变或疾病引起的,有痛觉过敏、感受异常、自发性疼痛、异常疼痛等临床特征,自身免疫性疾病、物理性机械损伤、神经毒性药物等均是其常见诱因,由于神经病理性疼痛发病机制复杂,导致其临床治疗效果不佳,因此,找出神经病理性疼痛发病机制的具体分子机制对于治疗这种慢性疾病具有重要意义[11,12]。研究中常用脊神经结扎等方式构建神经病理性疼痛模型,王裕生等[13]通过此方法构建的神经病理性疼痛模型大鼠出现明显的痛觉过敏现象,且能更好的模拟神经病理性疼痛的发生发展过程。本研究显示,与假手术组比较,模型组大鼠出现甩足、舔足、足趾并拢等行为学异常现象,MWT、TWL值显著升高,提示神经病理性疼痛大鼠模型构建成功。
多项研究表明,神经病理性疼痛与机体炎性反应联系密切,当脊神经受到损害后,损害部位、背根神经节、脊髓等迅速产生较多的促炎因子,从而导致大量炎性因子的产生,参与多种伤害性信息的传递,神经生长因子、促炎性细胞因子等均可作用于神经元细胞,从而增强神经元兴奋性、进一步放大神经炎性反应[14,15]。Ghasemzadeh等[16]研究发现,神经病理性疼痛大鼠体内TNF-α等炎性因子水平显著升高,而迷迭香的抗炎作用能有效减轻模型大鼠的痛觉过敏现象,因此靶向神经炎性反应可能是神经病理性疼痛治疗的新方向。近年来的研究表明miRNA参与调控神经病理性疼痛的炎性反应,Bao等[17]研究发现miR-28-5p过表达能显著抑制神经病理性疼痛大鼠体内IL-1β、IL-6等炎性因子表达水平并缓解大鼠痛觉异常等不良表现。Lu等[18]研究发现,miRNA-146a-5p可通过抑制星形胶质细胞中的TNF受体相关因子6(TRAF6)的表达来减轻炎性反应,进而减轻脊神经结扎诱导的神经病理性疼痛。miR-338-3p是miRNA的一员,与机体炎性反应调节密切相关[19]研究发现低表达的miR-338-3p能激活内皮细胞炎症信号通路,而过表达miR-338-3p能显著抑制相关炎症通路的激活,可能与神经病理性疼痛疾病联系密切[20,21]。
本研究结果显示,与假手术组比较,模型组大鼠背根神经节组织中TNF-α、IL-1β、IFN-γ表达水平显著升高,提示神经病理性疼痛与神经炎性反应密切相关;与模型组比较,miR-338-3p组大鼠背根神经节组织中炎性因子表达水平、MWT、TWL值均显著降低,提示过表达miR-338-3p能显著降低神经病理性疼痛引起的神经炎性反应,并具有一定的缓解神经病理性疼痛的作用。
采用TargetScan数据库结果对miR-338-3p靶基因进行预测,结果显示miR-338-3p在CaMKⅡγ mRNA 3’UTR存在相应的结合位点,CaMKⅡγ是CaMKⅡ家族成员之一,与神经元损伤、中枢神经痛觉过敏等方面联系密切[22,23],Pan等[24]研究发现完全弗氏佐剂诱导的慢性炎症痛小鼠脊髓中CaMKⅡγ表达水平显著升高,miR-219过表达可显著降低CaMKⅡγ的表达,并显著减轻小鼠的痛反应异常等行为。本研究结果显示,与模型组比较,miR-338-3p组及miR-338-3p+CaMKⅡγ组大鼠CaMKⅡγ mRNA及蛋白表达水平、背根神经节组织中TNF-α、IL-1β、IFN-γ表达水平均显著降低,荧光素酶实验结果显示miR-338-3p过表达能显著下调野生型CaMKⅡγ-3’UTR荧光素酶活性,表明在脊根神经节中miR-338-3p可直接作用于CaMKⅡγ的3’UTR区,进而调控CaMKⅡγ的基因表达。与miR-338-3p组比较,miR-338-3p+CaMKⅡγ组大鼠CaMKⅡγ mRNA及蛋白表达水平、背根神经节组织中TNF-α、IL-1β、IFN-γ表达水平均显著升高,表明miR-338-3p可能通过靶向抑制CaMKⅡγ表达,进而降低神经病理性疼痛引起的神经炎性反应。
综上所述,miR-338-3p可能通过抑制CaMKⅡγ的表达缓解神经病理性疼痛大鼠的痛觉过敏及神经炎性反应,这可能为临床治疗神经病理性疼痛提供新思路,但本研究并未采用CaMKⅡγ相关干扰技术进行对比实验,因此仍需深入研究。
