腌制风干过程中卵形鲳鲹鱼肉性质、蛋白质氧化及游离氨基酸的变化
2022-10-09张进伟陈胜军赵永强吴燕燕王悦齐
张进伟,胡 晓,陈胜军,3,4,*,赵永强,4,吴燕燕,王悦齐,3,潘 创,王 迪
(1.中国水产科学研究院南海水产研究所,农业农村部水产品加工重点实验室,国家水产品加工技术研发中心,广东 广州 510300;2.上海海洋大学食品学院,上海 201306;3.三亚热带水产研究院,海南 三亚 572000;4.大连工业大学 海洋食品精深加工关键技术省部共建协同创新中心,辽宁 大连 116034)
卵形鲳鲹(),俗称金鲳、黄腊鲳,其肉质鲜嫩且无肌间刺,富含蛋白质、脂肪、多不饱和脂肪酸等营养物质,是海水养殖经济鱼种。2020年我国卵形鲳鲹养殖产量达到10.1万 t。在我国南方地区,人们常通过腌制、风干工艺制作腌腊鱼、风干鱼等,从而提高原料的附加值,延长产品保质期,提高产品风味。蛋白质是水产品的主要成分之一,加工过程中的氧化水解影响产品品质。多肽和游离氨基酸是在内源蛋白酶的作用下降解蛋白质产生,是风味物质的前体物质。游离氨基酸可以与还原糖发生美拉德反应,生成挥发性气味物质,对产品风味形成具有重要作用。此外,有研究表明肌原纤维蛋白降解会影响鱼肉的硬度和弹性。因此,有必要对腌制风干过程中鱼肉的蛋白质氧化及游离氨基酸变化规律进行探讨。
目前已有一些国内外学者对水产品腌制风干加工过程中蛋白质氧化和游离氨基酸的变化进行报道。Zhang Jinjie等研究发现腊鱼在腌制风干过程中游离氨基酸含量显著上升。吴燕燕等研究发现带鱼在腌干过程中蛋白质在蛋白酶、氨肽酶的作用下发生水解,蛋白水解指数呈上升趋势;而非蛋白含量在腌制后下降,在风干过程中增加。张会丽等在风干鲈鱼过程中发现随着风干温度的升高,蛋白水解指数增加,而游离氨基酸总量呈先增加后减少的趋势。本研究主要对卵形鲳鲹在腌制风干加工过程中鱼肉含水率、盐含量、质构特性、肌原纤维蛋白二级结构、羰基含量、巯基含量、蛋白质水解指数、十二烷基硫酸钠-聚丙烯酰胺凝胶电泳(sodium dodecyl sulfate-polyacrylamide gel electrophoresis,SDS-PAGE)图谱和游离氨基酸等进行测定分析,初步揭示卵形鲳鲹鱼肉在腌制风干过程中蛋白质氧化及游离氨基酸变化规律,为腌制水产品加工过程中品质控制提供一定理论参考。
1 材料与方法
1.1 材料与试剂
卵形鲳鲹长(31.32±0.88)cm、体质量(537.80±52.38)g,2021年5月购于广东广州市新港西路华润万家超市,置于-20 ℃冰箱中保存。
甲醇(色谱纯)、乙腈(色谱纯)、硝酸银标准滴定溶液(AgNO浓度0.1 mol/L) 上海安谱实验科技股份有限公司;17 种氨基酸混合标准品(2.5 μmol/mL)、天冬酰胺、谷氨酰胺、瓜氨酸、-正缬氨酸、色氨酸、羟脯氨酸、肌氨酸标准品 西格玛奥德里奇(上海)贸易有限公司;考马斯亮蓝法蛋白检测试剂盒、蛋白质羰基检测试剂盒、总巯基检测试剂盒 南京建成海浩生物科技有限公司;NuPAGE12% Bis-Tris预制胶英潍捷基(上海)贸易有限公司;苯甲基磺酰氟(phenylmethanesulfonyl fluoride,PMSF)、BeyoBlue™考马斯亮蓝超快染色液、SDS-PAGE蛋白上样缓冲液(5×)、BeyoColor彩色预染蛋白分子质量标准 上海碧云天生物技术有限公司;三氯乙酸、溴化钾(均为分析纯) 广州化学试剂厂;实验用其他试剂均为分析纯。
1.2 仪器与设备
RCX-1500-1540热泵除湿干燥机 佛山市顺德区勒流镇显高冷冻设备厂;H1850R台式高速冷冻离心机湘仪离心机仪器有限公司;809 Titrando自动电位滴定仪瑞士万通公司;T50均质机 德国IKA公司;Kjeltec2300全自动凯式定氮仪 丹麦福斯分析仪器公司;Brookfiled CT3型质构仪 美国博勒飞公司;IRAffinity-1傅里叶变换红外光谱仪 日本岛津公司;Mini Gel Tank PAGE系统 美国赛默飞科技公司;1100液相色谱仪(配有可变波长紫外检测器) 安捷伦科技(中国)有限公司。
1.3 方法
1.3.1 样品制备
卵形鲳鲹流水解冻后,剖开鱼腹,除去鱼鳃、内脏、黑膜等,用流动水清洗鱼体内残血等脏污,室温下沥干水分,以备后续腌制使用。称取鱼体质量10%的食盐,将其均匀涂抹在鱼体表面及内部,然后将鱼整齐堆放于腌制容器中室温下腌制2 d,每12 h将鱼翻动一次。腌制2 d后将鱼取出,除去鱼体表面的盐分,将鱼置于热泵除湿干燥机中干制4 d。参考任中阳等的工艺参数和预实验设置热泵干燥机工艺参数:温度(30±2) ℃、相对湿度(30±5)%。
1.3.2 含水率测定
按照GB 5009.3—2016《食品中水分的测定》中直接干燥法进行测定。
1.3.3 盐含量测定
按照GB 5009.44—2016《食品中氯化物的测定》中电位滴定法进行测定。
1.3.4 质构特性测定
参考Lerfall等的方法并稍作修改。取鱼背部肌肉将其修整为2 cm×2 cm×1 cm(长×宽×高),使用Brookfiled CT3型质构仪及TA 19平底圆柱形探头进行测定,测定模式为质地多面剖析(TPA)模式,测试速度1 mm/s,触发点负载5.0 g,下压距离5 mm,每组实验重复8 次。
1.3.5 肌原纤维蛋白提取
参考Pazos等的方法并稍作修改。精确称取2 g绞碎鱼肉样品于50 mL离心管中,加入10 mL预冷却的匀浆缓冲溶液(pH 7.2,5 mmol/L PMSF和10 mmol/L Tris-HCl缓冲液),冰浴条件下以12 000 r/min均质1 min(均质10 s,间隔10 s,防止过热蛋白质变性)。均质后在4 ℃、12 000 r/min离心15 min,弃去上清液,向沉淀中加入10 倍体积的盐溶液(pH 7.2,0.6 mol/L NaCl、5 mmol/L PMSF和10 mmol/L Tris-HCl缓冲液),均质混匀。冰水浴静置25 min,再以4 ℃、12 000 r/min离心15 min,所得上清液为肌原纤维蛋白溶液,测定肌原纤维蛋白含量,并置于-80 ℃冰箱中保存。
1.3.6 巯基与羰基含量测定
按照蛋白巯基试剂盒与蛋白羰基试剂盒的说明书对提取的肌原纤维蛋白中巯基与羰基含量进行测定。
1.3.7 傅里叶变换红外光谱测定
肌原纤维蛋白溶液置于-80 ℃冰箱冷冻4 h后取出,经真空冷冻干燥72 h后,得到肌原纤维蛋白冻干样品以备后续使用。参考李锐等的方法并略作修改,将肌原纤维蛋白冻干样品与溴化钾按1∶100的质量比混合,充分研磨,压制成薄片。使用傅里叶变换红外光谱全波段扫描(4 000~400 cm),分辨率4.0 cm,扫描次数32 次。使用Peakfit v4.12软件处理数据,分析蛋白质二级结构及其相对含量。
1.3.8 SDS-PAGE测定
参照Pan Chuang等的方法,使用Mini Gel Tank小型胶电泳槽对12%预制胶进行SDS-PAGE分析。用盐溶液将肌原纤维蛋白溶液稀释到1 mg/mL,取30 μL 1 mg/mL肌原纤维蛋白溶液与10 μL上样缓冲液混合,沸水浴5 min,上样量10 μL,电泳电压设置为120 V。电泳结束时后取下凝胶,使用考马斯亮蓝超快染色液染色,脱色至背景无色,用凝胶成像系统扫描并分析条带。
1.3.9 总氮与非蛋白氮含量测定
总氮测定:参考GB 5009.5—2016《食品中蛋白质的测定》中自动凯式定氮仪法进行测定。
非蛋白氮测定:参考吴燕燕等方法并稍作修改。准确称取5 g鱼肉(精确至0.001 g),加入5 倍体积预冷的质量分数10%的三氯乙酸溶液 ,22 000 r/min高速匀浆(3×10 s),4 ℃条件下放置12 h后取出,4 000 r/min离心5 min。用中速滤纸过滤上清液,用少量质量分数10%三氯乙酸洗涤沉淀,过滤后合并上清液并定容至100 mL。取10 mL滤液消化,用自动凯式定氮仪测定。
按下式计算蛋白水解指数:
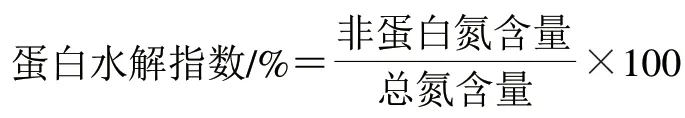
1.3.10 游离氨基酸含量测定
1.3.10.1 样品前处理
称取1 g绞碎鱼肉样品于10 mL离心管中,与5 mL 0.01 mol/L盐酸溶液混匀,沸水浴30 min后,10 000 r/min离心10 min,取上清液。向沉淀中加入2 mL 0.01 mol/L盐酸溶液,悬浮超声5 min,10 000 r/min离心10 min,合并上清液并定容至10 mL,过0.22 μm滤膜后上机测定。
1.3.10.2 液相色谱条件
ZORBAX Eclipse AAA氨基酸分析色谱柱(4.6 mm×150 mm,3.5 μm);流速:1 mL/min;流动相:A为40 mmol/L磷酸二氢钠(pH 7.8),B为乙腈-甲醇-水(45∶45∶10,/);梯度洗脱:0~1 min,100% A、0% B;1~23 min,43% A、57%B;23~34 min,0% A、100% B;34~41 min,100%A、0% B;紫外检测器波长:338 nm(0~19 min),266 nm(19.01~25 min)。
1.4 数据处理
每个指标设置3 个平行。采用Excel进行数据处理和作图,采用SPSS Statistics 23.0软件进行统计分析,Duncan法进行差异显著性分析。<0.05,差异显著。
2 结果与分析
2.1 腌制风干对卵形鲳鲹含水率和盐含量的影响
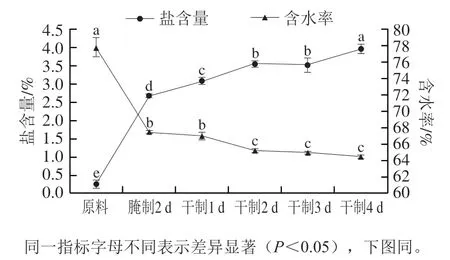
图1 卵形鲳鲹在腌制风干过程中盐含量和含水率的变化Fig. 1 Change in salt content and water content of T. ovatus during curing and air-drying
由图1可知,在腌制、风干过程中卵形鲳鲹的盐含量呈上升趋势,其中腌制2 d后卵形鲳鲹的盐含量急剧增加(<0.05);而在此过程中含水率呈下降趋势,分别在干制1、3、4 d后含水率变化差异不显著(>0.05)。腌制是渗透扩散的过程,干腌是将鱼体表面及内部均匀涂抹食盐,使其内外形成高渗透压,从而使盐含量显著增加,水分含量下降。在风干过程中盐含量逐渐增加,但干制2 d与3 d后卵形鲳鲹盐含量差异不显著(>0.05),这可能是由于卵形鲳鲹表皮水分快速脱除,内部水分缓慢迁移,形成硬壳,使含水率变化缓慢,盐含量变化不显著。此外,在干制4 d后,卵形鲳鲹的盐含量达到最高值,为(3.96±0.13)%。
2.2 腌制风干对卵形鲳鲹质构特性的影响
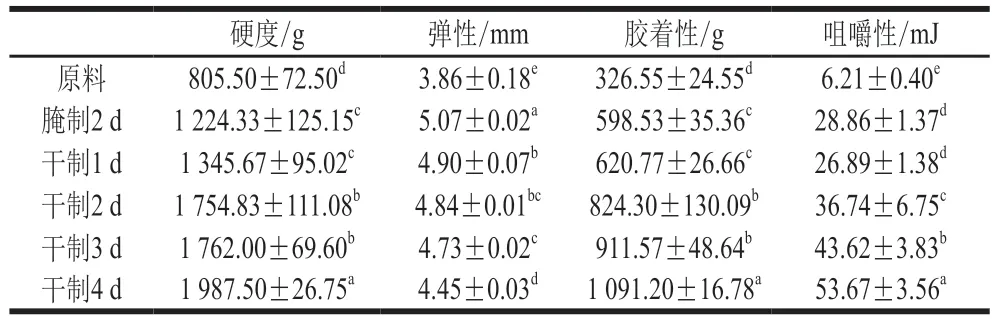
表1 卵形鲳鲹在腌制风干过程中质构特性的变化Table 1 Change in texture characteristics of T. ovatus during curing and air-drying processes
质构特性主要由硬度、弹性、胶着性、咀嚼性等性质组成,可以反映鱼肉的组织状态,是衡量其食用价值的重要指标。鱼肉质构特性的变化与其水分、pH值、细胞外基质(胶原蛋白)结构、蛋白质水解等因素有关。由表1可知,随着腌制风干的进行,卵形鲳鲹背部肌肉硬度增加,造成该现象的原因是鱼肉经腌制、风干后水分含量减少,盐含量升高,肌纤维收缩,肉质紧密,从而使鱼肉硬度增加。Yang Wenxian等研究发现草鱼在腌制2 d后,鱼肉硬度显著增加,与本研究结果相似。由表1可知,鱼肉在腌制2 d后弹性增加,并随着风干的进行鱼肉弹性降低。通常鱼肉的盐含量较低时,其弹性会有所增加,而在风干过程中鱼肉的盐含量明显增加,水分含量减少,弹性随含水率降低而降低,同时随着鱼肉中内部水分向外扩散,肌纤维直径及其横截面积减小,肌纤维收缩,导致鱼肉弹性下降。由表1可知,在腌制风干加工过程中,鱼肉的胶着性呈上升趋势。有研究表明脂肪含量对胶着性具有一定影响,而在腌制风干过程中鱼肉水分含量减少,脂肪相对含量增加,所以可能导致鱼肉胶着性的变化。由表1可知,卵形鲳鲹鱼肉咀嚼性整体呈上升趋势;其中,在腌制2 d后,鱼肉咀嚼性显著增加,而在风干1 d后略有下降但并不显著,之后呈显著增加趋势(<0.05)。
2.3 腌制风干对卵形鲳鲹肌原纤维蛋白含量的影响
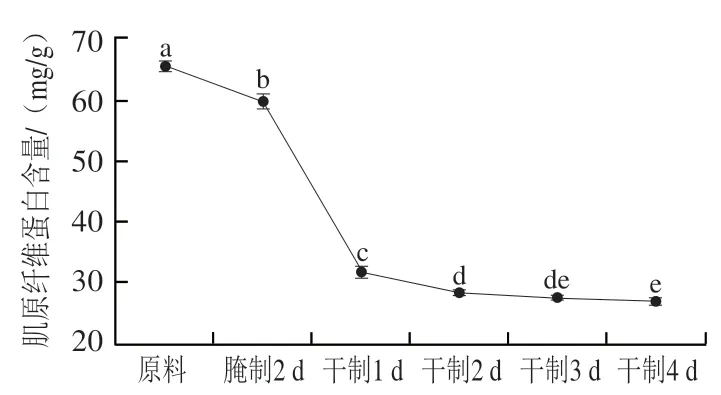
图2 卵形鲳鲹在腌制风干过程中肌原纤维蛋白含量的变化Fig. 2 Change in myofibrillar protein content of T. ovatus during curing and air-drying
肌原纤维蛋白是鱼肉中重要蛋白质,约占总蛋白含量的50%~70%,决定鱼肉质构、热凝胶等加工特性,影响产品品质。由图2可知,卵形鲳鲹在腌制风干过程中肌原纤维蛋白含量呈下降趋势;在腌制2 d后、干制1 d后和2 d后,肌原纤维蛋白含量显著降低(<0.05),可能是加工过程中肌原纤维蛋白的溶出或水解产生多肽和游离氨基酸所致。
2.4 腌制风干对卵形鲳鲹蛋白质羰基含量的影响
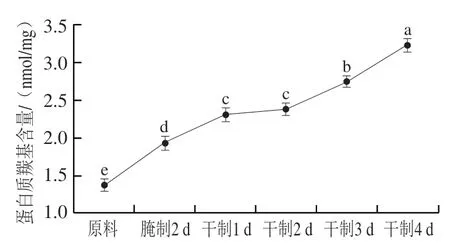
图3 卵形鲳鲹在腌制风干过程中羰基含量的变化Fig. 3 Change in carbonyl content of T. ovatus during curing and air-drying
蛋白质羰基主要由赖氨酸、脯氨酸、精氨酸等直接氧化生成,是评价蛋白氧化的重要指标。由图3可知,随着腌制风干的进行,羰基含量明显增加,蛋白质氧化程度增加;卵形鲳鲹原料的蛋白质羰基含量为1.37 nmol/mg,腌制2 d后蛋白质羰基含量显著增加(<0.05)至1.94 nmol/mg,风干过程中蛋白质羰基含量持续增加,而干制1 d和2 d后蛋白质羰基含量差异不显著(>0.05)。赵亚南等研究发现在一定范围内随着氯化钠添加量的增加,蛋白质羰基含量增加。本实验中,卵形鲳鲹在腌制过程中蛋白质羰基含量的增加,可能是氯化钠渗入鱼肉内部,增强Fe的活性,Fe进一步影响鱼肉蛋白氧化所致。此外,卵形鲳鲹鱼肉中富含脂肪,在腌制风干过程中脂肪发生氧化降解,而脂肪氧化过程中产生自由基,自由基攻击蛋白质氨基酸侧链、肽链骨架,导致氨基酸侧链暴露,发生蛋白质聚合、交联等反应,可能使蛋白质羰基含量增加。
2.5 腌制风干对卵形鲳鲹巯基含量的影响
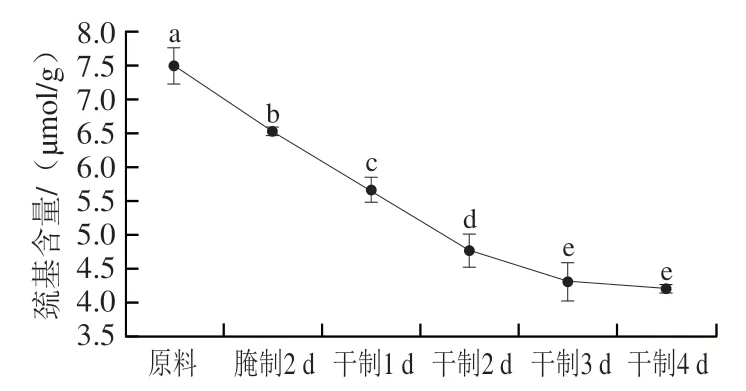
图4 卵形鲳鲹在腌制风干过程中巯基含量的变化Fig. 4 Change in sulfhydryl content of T. ovatus during curing and air-drying
活性巯基主要存在于半胱氨酸,是蛋白质结构和生物体内某些氧化还原反应的重要基团。在同一肽链或不同肽链间的半胱氨酸残基的巯基脱氢氧化形成二硫键,二硫键含量上升,蛋白质变性,所以巯基含量能够表征蛋白质氧化程度。由图4可知,巯基含量的变化趋势与羰基整体相反,随着腌制风干的进行,巯基含量显著下降(<0.05),在干制4 d后巯基含量差异不显著(>0.05)。造成该现象的原因是腌制、风干工艺使鱼肉蛋白质结构被破坏,导致埋藏在蛋白质内部的巯基暴露出来,产生二硫键,使总巯基含量下降。叶路漫研究发现鱼肉总巯基含量随着腌制、风干工艺时间推移而下降,与本研究结果相近。
2.6 腌制风干对卵形鲳鲹肌原纤维蛋白二级结构的影响
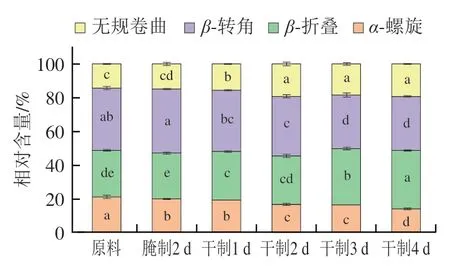
图5 卵形鲳鲹在腌制风干过程中肌原纤维蛋白二级结构组成变化Fig. 5 Change in secondary structure composition of myofibrillar protein from T. ovatus during curing and air-drying
蛋白质二级结构在红外光谱区有若干吸收带,其中酰胺I带(1 600~1 700 cm)是二级结构的关键吸收带,主要由羰基C=O伸缩振动引起。蛋白质的二级结构主要包括-折叠(1 600~1 639 cm)、无规卷曲(1 640~1 650 cm)、-螺旋(1 651~1 660 cm)、-转角(1 661~1 700 cm)。-螺旋具有高度稳定性,主要由多肽链上羰基和氨基之间的氢键维持,是蛋白质中最主要最稳定的结构,而无规卷曲和-结构的紧密程度和构象稳定性相较于-螺旋较差。
由图5可知,随着腌制风干的进行,肌原纤维蛋白二级结构中-螺旋含量减少,而无规卷曲含量增加。有研究表明氯化钠可能对肌原纤维蛋白的二级结构具有一定破坏作用。可能是因为卵形鲳鲹在腌制风干过程中食盐渗透进入鱼肉内部,肌原纤维蛋白质结构被破坏,羰基含量上升,从而使-螺旋含量减少,无规卷曲含量增加。此外,曹锦轩等研究发现腊肉肌原纤维蛋白在加工过程中-螺旋含量减少,与本研究结果类似。通常在加热条件下,-螺旋含量降低代表蛋白质分子展开程度增加,-折叠含量增加代表蛋白质分子间聚集程度增加。由图5可知,卵形鲳鲹在加工过程中-转角含量减少,而-折叠含量增加,蛋白质分子间聚集程度增加。
2.7 卵形鲳鲹肌原纤维蛋白的SDS-PAGE分析
在肉质品加工过程中,肌原纤维蛋白是主要的蛋白质,与肉制品质构特性密切相关。如图6所示,在加工过程中,卵形鲳鲹肌原纤维蛋白的SDS-PAGE条带发生明显变化,其中主要的蛋白质有肌球蛋白重链(myosin heavy chain,MHC)、肌动蛋白、原肌球蛋白等。蛋白分子降解主要表现为分子质量较高的条带出现模糊、弱化、扩展,较低分子质量区域出现新条带或条带颜色加深。由图6可知,MHC条带和肌动蛋白发生明显变化;其中MHC条带在腌制2 d后变化较小,可能是腌制时肌原纤维蛋白发生交联或聚集;而风干后MHC条带消失,表明大分子质量的蛋白质在内源蛋白酶作用下发生降解。肌动蛋白条带在腌制2 d后变细,造成此现象的原因可能是氯化钠浓度升高,导致肌动蛋白溶出;而干制1 d后肌动蛋白条带又变粗,可能是由于在风干过程中大分子质量蛋白降解形成较小分子质量蛋白所致。
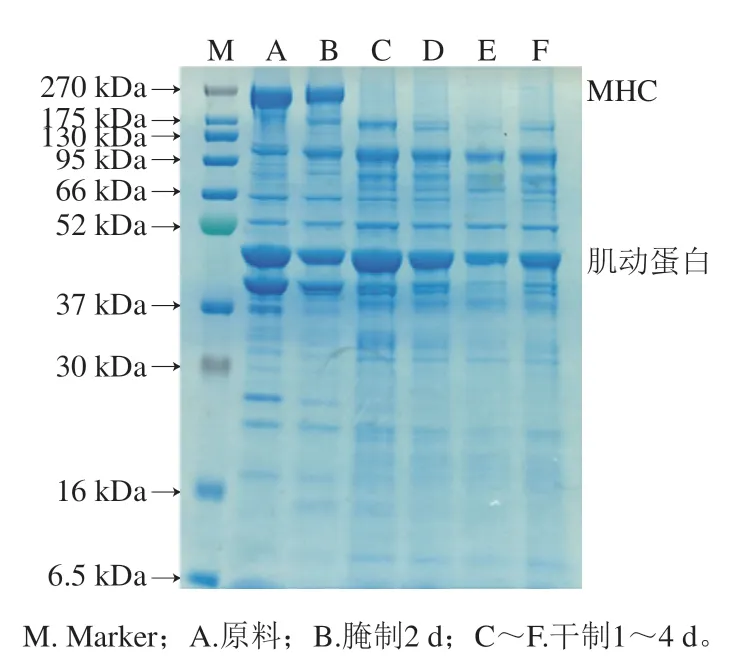
图6 卵形鲳鲹在腌制风干中肌原纤维蛋白的SDS-PAGE图谱Fig. 6 SDS-PAGE pattern of myofibrillar protein from T. ovatus during curing and air-drying
2.8 腌制风干对卵形鲳鲹总氮与非蛋白氮含量的影响
非蛋白氮是蛋白质和多肽在内源蛋白酶和氨肽酶的作用下降解产生。如图7所示,在加工过程中,卵形鲳鲹总氮含量在腌制2 d后显著下降(<0.05),可能是在腌制过程中水溶性蛋白质溶出所致,这与张会丽的研究结果一致;而在风干过程中总氮含量整体呈缓慢下降的趋势。此外,在腌制加工后非蛋白氮含量下降,造成该现象的原因是水溶性含氮物质溶出;而在风干过程中非蛋白氮含量明显增加,主要是在鱼肉内源蛋白酶的作用下,蛋白质水解为多肽、游离氨基酸等造成的。史培磊等研究发现风鹅加工过程中非蛋白氮含量在腌制后下降,而在风干过程中先上升后下降,这与本研究结果有所不同,可能是风干工艺时间不同所致。
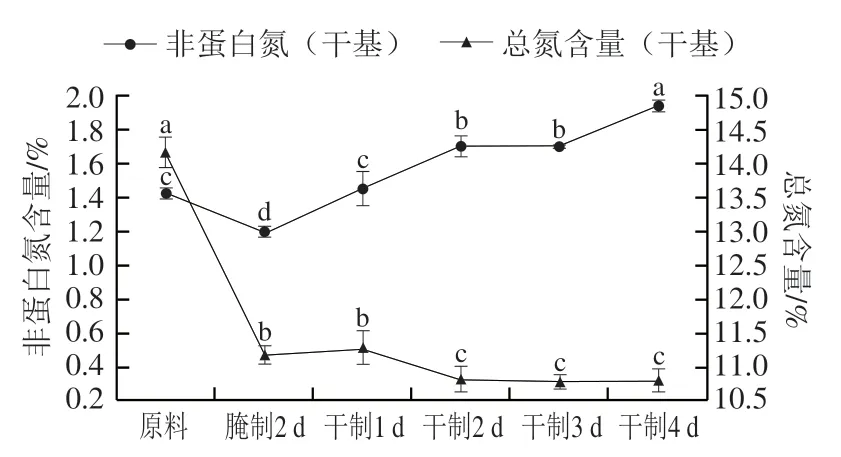
图7 卵形鲳鲹在腌制风干中总氮、非蛋白氮的变化Fig. 7 Change in total nitrogen and non-protein nitrogen contents of T. ovatus during curing and air-drying
蛋白质水解主要通过内源蛋白酶、微生物酶等作用产生降解,蛋白质的水解可以增加肉质的柔软性,主要与肌原纤维的破碎有关,同时产生多肽、游离氨基酸等物质,对风味形成有间接作用。由图8可知,蛋白水解指数在腌制2 d后变化较小,差异不显著(>0.05);而在风干过程中蛋白水解指数明显升高,原因可能是在风干过程中酶活性提高,鱼肉蛋白质降解为多肽、游离氨基酸等,同时含氮物质溶出较少,从而使蛋白水解指数增加。
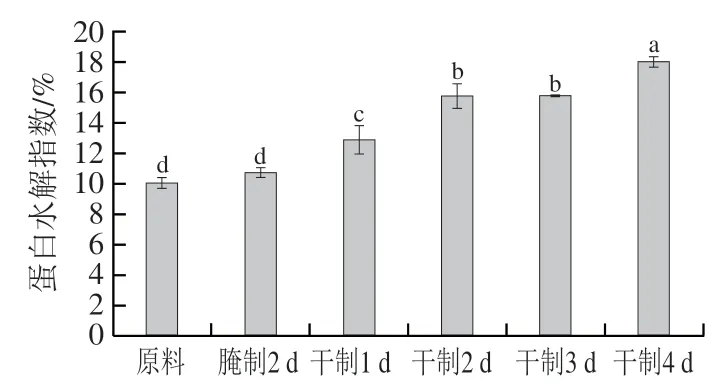
图8 卵形鲳鲹在腌制风干中蛋白水解指数的变化Fig. 8 Change in proteolysis index of T. ovatus during curing and air-drying
2.9 腌制风干对卵形鲳鲹游离氨基酸含量的影响

表2 卵形鲳鲹在腌制风干中游离氨基酸的变化Table 2 Change in free amino acids of T. ovatus during curing and air-drying mg/100 g
游离氨基酸通过蛋白质降解产生,是水产品中重要的风味物质,其呈味特征与含量、阈值有关。其中丙氨酸、甘氨酸、苏氨酸、丝氨酸、脯氨酸呈现甜味;天冬氨酸、谷氨酸呈现鲜味。如表2所示,原料中游离氨基酸总量为380.03 mg/100 g,其中丙氨酸、羟脯氨酸、甘氨酸、谷氨酸含量较高;而在干制4 d后游离氨基酸总量为1 596.07 mg/100 g,其中丙氨酸、甘氨酸、谷氨酸、亮氨酸、赖氨酸含量较高,约占总游离氨基酸含量的50.67%。游离氨基酸总量在腌制2 d后显著增加(<0.05),这与吴燕燕等研究结果相近;在干制前3 d游离氨基酸总量仍显著增加(<0.05),表明在风干过程中蛋白质在内源酶的作用下水解产生游离氨基酸;干制4 d后游离氨基酸总量下降,可能是由于部分氨基酸与还原糖发生美拉德反应以及在微生物作用下生成风味物质所致。鲜味氨基酸和甜味氨基酸在加工过程中含量总体呈上升趋势,有利于鱼肉形成良好风味,而在干制4 d后其含量略有下降。
3 结 论
在腌制风干过程中,卵形鲳鲹鱼肉的盐含量整体呈上升趋势,同时其质构特性发生变化:硬度、胶着性、咀嚼性整体呈上升趋势,而弹性为先上升后下降。在加工过程中,鱼肉蛋白质发生氧化反应,羰基含量增加,而巯基含量下降;肌原纤维蛋白二级结构中-螺旋含量下降,而无规卷曲含量增加;MHC条带和肌动蛋白发生明显变化:MHC条带变浅并在风干后消失,肌原纤维蛋白产生降解;蛋白质水解指数增加,非蛋白氮含量在腌制后下降而风干过程中增加;游离氨基酸总量在干制1 d后达到最高,鲜味氨基酸和甜味氨基酸含量总体呈上升趋势,有利于提高产品风味。
